
Immunoinformatics Identification of the Conserved and Cross-Reactive T-Cell Epitopes of SARS-CoV-2 with Human Common Cold Coronaviruses, SARS-CoV, MERS-CoV and Live Attenuated Vaccines Presented by HLA Alleles of Indonesian Population

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Coronavirus Proteome Sequence Retrieval
2.2. HLA Alleles Predominant in Indonesia Identification
2.3. Cytotoxic T Lymphocyte (CTL) Epitopes Prediction
2.4. Helper T Lymphocyte (HTL) Epitopes Prediction
2.5. IEDB Epitopes Validation
2.6. Conservancy and Cross-Reactivity Analysis of SARS-CoV-2 T-Cell Epitopes with HCCCs
2.7. Cross-Reactivity Analysis of SARS-CoV-2 T-Cell Epitopes with OPV, BCG, and MMR
2.8. Cross-Reactivity of Predicted Epitopes with Human Peptides
2.9. Population Coverage Analysis
2.10. Epitope Selection for Universal Coronavirus Vaccine Construct
3. Results
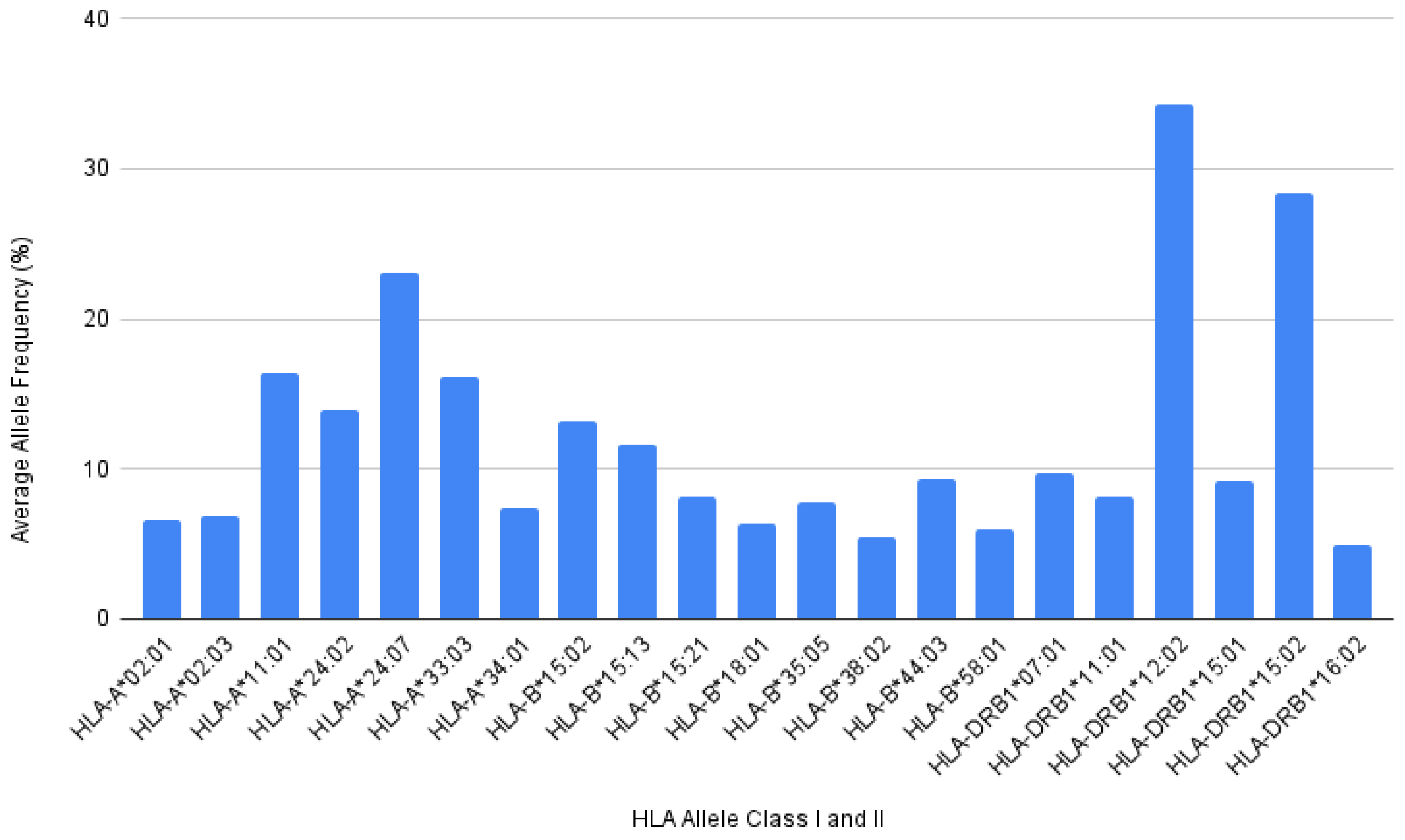
3.1. HLA Allele Frequencies of Indonesian Population
3.2. Prediction of CTL Epitopes from SARS-CoV-2 Proteins
3.3. Prediction of HTL Epitopes from SARS-CoV-2 Proteins
3.4. Identification of Conserved Epitopes in SARS-CoV-2 and Human CCCs
3.5. Epitopes with Identical TCR Contact Residue between SARS-CoV-2 and Human CCC
3.6. Cross-Reactive T-Cell Epitopes between SARS-CoV-2 and BCG
3.7. Cross-Reactive T-Cell Epitopes between SARS-CoV-2 and MMR
3.8. Potential Epitope Set for Coronavirus Universal Vaccine Construct and Its Population Coverage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Naqvi, A.A.T.; Fatima, K.; Mohammad, T.; Fatima, U.; Singh, I.K.; Singh, A.; Atif, S.M.; Hariprasad, G.; Hasan, G.M.; Hassan, I. Insights into SARS-CoV-2 genome, structure, evolution, pathogenesis and therapies: Structural genomics approach. Biochim. Biophys. Acta (BBA)–Mol. Basis Dis. 2020, 1866, 165878. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Coronavirus COVID-19 Dashboard. Available online: https://covid19.who.int/ (accessed on 18 May 2022).
- Zheng, J. SARS-CoV-2: An Emerging Coronavirus that Causes a Global Threat. Int. J. Biol. Sci. 2020, 16, 1678–1685. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Tracking Sars-COV-2 Variants. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants/ (accessed on 18 May 2021).
- Lim, Y.X.; Ng, Y.L.; Tam, J.P.; Liu, D.X. Human Coronaviruses: A Review of Virus–Host Interactions. Diseases 2016, 4, 26. [Google Scholar] [CrossRef]
- Poland, G.A.; Ovsyannikova, I.G.; Kennedy, R.B. SARS-CoV-2 immunity: Review and applications to phase 3 vaccine candidates. Lancet 2020, 396, 1595–1606. [Google Scholar] [CrossRef]
- Tomita, Y.; Sato, R.; Ikeda, T.; Sakagami, T. BCG vaccine may generate cross-reactive T cells against SARS-CoV-2: In silico analyses and a hypothesis. Vaccine 2020, 38, 6352–6356. [Google Scholar] [CrossRef] [PubMed]
- Altmann, D.M.; Boyton, R.J. SARS-CoV-2 T cell immunity: Specificity, function, durability, and role in protection. Sci. Immunol. 2020, 5, eabd6160. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Pre-existing immunity to SARS-CoV-2: The knowns and unknowns. Nat. Rev. Immunol. 2020, 20, 457–458. [Google Scholar] [CrossRef]
- Echeverría, G.; Guevara, Á.; Coloma, J.; Ruiz, A.M.; Vasquez, M.M.; Tejera, E.; de Waard, J.H. Pre-existing T-cell immunity to SARS-CoV-2 in unexposed healthy controls in Ecuador, as detected with a COVID-19 Interferon-Gamma Release Assay. Int. J. Infect. Dis. 2021, 105, 21–25. [Google Scholar] [CrossRef]
- Chumakov, K.; Benn, C.S.; Aaby, P.; Kottilil, S.; Gallo, R. Can existing live vaccines prevent COVID-19? Science 2020, 368, 1187–1188. [Google Scholar] [CrossRef]
- Chumakov, K.; Avidan, M.S.; Benn, C.S.; Bertozzi, S.M.; Blatt, L.; Chang, A.Y.; Jamison, D.T.; Khader, S.A.; Kottilil, S.; Netea, M.G.; et al. Old vaccines for new infections: Exploiting innate immunity to control COVID-19 and prevent future pandemics. Proc. Natl. Acad. Sci. USA 2021, 118, e2101718118. [Google Scholar] [CrossRef]
- Reche, P.A. Potential Cross-Reactive Immunity to SARS-CoV-2 From Common Human Pathogens and Vaccines. Front. Immunol. 2020, 11, 586984. [Google Scholar] [CrossRef] [PubMed]
- Haddad-Boubaker, S.; Othman, H.; Touati, R.; Ayouni, K.; Lakhal, M.; Ben Mustapha, I.; Ghedira, K.; Kharrat, M.; Triki, H. In silico comparative study of SARS-CoV-2 proteins and antigenic proteins in BCG, OPV, MMR and other vaccines: Evidence of a possible putative protective effect. BMC Bioinform. 2021, 22, 163. [Google Scholar] [CrossRef]
- Comunale, B.A.; Engineer, L.; Jiang, Y.; Andrews, J.C.; Liu, Q.; Ji, L.; Yurkovich, J.T.; Comunale, R.A.; Xie, Q. Poliovirus Vaccination Induces a Humoral Immune Response That Cross Reacts with SARS-CoV-2. Front. Med. 2021, 8, 1285. [Google Scholar] [CrossRef] [PubMed]
- Tarabini, R.F.; Rigo, M.M.; Fonseca, A.F.; Rubin, F.; Bellé, R.; Kavraki, L.E.; Ferreto, T.C.; Antunes, D.A.; de Souza, A.P.D. Large-Scale Structure-Based Screening of Potential T Cell Cross-Reactivities Involving Peptide-Targets From BCG Vaccine and SARS-CoV-2. Front. Immunol. 2022, 12, 5858. [Google Scholar] [CrossRef] [PubMed]
- Eggenhuizen, P.J.; Ng, B.H.; Chang, J.; Fell, A.L.; Cheong, R.M.Y.; Wong, W.Y.; Gan, P.-Y.; Holdsworth, S.R.; Ooi, J.D. BCG Vaccine Derived Peptides Induce SARS-CoV-2 T Cell Cross-Reactivity. Front. Immunol. 2021, 12, 3034. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Basic Immunology: Functions and Disorders of the Immune System; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Wieczorek, M.; Abualrous, E.T.; Sticht, J.; Álvaro-Benito, M.; Stolzenberg, S.; Noé, F.; Freund, C. Major Histocompatibility Complex (MHC) Class I and MHC Class II Proteins: Conformational Plasticity in Antigen Presentation. Front. Immunol. 2017, 8, 292. [Google Scholar] [CrossRef]
- Langton, D.J.; Bourke, S.C.; Lie, B.A.; Reiff, G.; Natu, S.; Darlay, R.; Burn, J.; Echevarria, C. The influence of HLA genotype on the severity of COVID-19 infection. HLA 2021, 98, 14–22. [Google Scholar] [CrossRef]
- Migliorini, F.; Torsiello, E.; Spiezia, F.; Oliva, F.; Tingart, M.; Maffulli, N. Association between HLA genotypes and COVID-19 susceptibility, severity and progression: A comprehensive review of the literature. Eur. J. Med. Res. 2021, 26, 84. [Google Scholar] [CrossRef]
- Gustiananda, M. What do T cells see in SARS-CoV2? Indones. J. Life Sci. 2020, 2, 29–43. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, J.; Mangalam, A.K.; Channappanavar, R.; Fett, C.; Meyerholz, D.K.; Agnihothram, S.; Baric, R.S.; David, C.S.; Perlman, S. Airway Memory CD4+ T Cells Mediate Protective Immunity against Emerging Respiratory Coronaviruses. Immunity 2016, 44, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Pinho, M.P.; Buckley, P.R.; Woodhouse, I.B.; Ogg, G.; Simmons, A.; Napolitani, G.; Koohy, H. Potential CD8+ T Cell Cross-Reactivity against SARS-CoV-2 Conferred by Other Coronavirus Strains. Front. Immunol. 2020, 11, 579480. [Google Scholar] [CrossRef]
- Gustiananda, M.; Sulistyo, B.P.; Agustriawan, D.; Andarini, S. Immunoinformatics Analysis of SARS-CoV-2 ORF1ab Polyproteins to Identify Promiscuous and Highly Conserved T-Cell Epitopes to Formulate Vaccine for Indonesia and the World Population. Vaccines 2021, 9, 1459. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Galarza, F.F.; McCabe, A.; Dos Santos, E.J.M.; Jones, J.; Takeshita, L.; Ortega-Rivera, N.D.; Del Cid-Pavon, G.M.; Ramsbottom, K.; Ghattaoraya, G.; Alfirevic, A.; et al. Allele frequency net database (AFND) 2020 update: Gold-standard data classification, open access genotype data and new query tools. Nucleic Acids Res. 2019, 48, D783–D788. [Google Scholar] [CrossRef] [PubMed]
- Stranzl, T.; Larsen, M.V.; Lundegaard, C.; Nielsen, M. NetCTLpan: Pan-specific MHC class I pathway epitope predictions. Immunogenetics 2010, 62, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Calis, J.J.A.; Maybeno, M.; Greenbaum, J.A.; Weiskopf, D.; De Silva, A.D.; Sette, A.; Keşmir, C.; Peters, B. Properties of MHC Class I Presented Peptides That Enhance Immunogenicity. PLoS Comput. Biol. 2013, 9, e1003266. [Google Scholar] [CrossRef] [PubMed]
- Reynisson, B.; Barra, C.; Kaabinejadian, S.; Hildebrand, W.H.; Peters, B.; Nielsen, M. Improved Prediction of MHC II Antigen Presentation through Integration and Motif Deconvolution of Mass Spectrometry MHC Eluted Ligand Data. J. Proteome Res. 2020, 19, 2304–2315. [Google Scholar] [CrossRef]
- Dhanda, S.K.; Vir, P.; Raghava, G.P.S. Designing of interferon-gamma inducing MHC class-II binders. Biol. Direct 2013, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef]
- Bui, H.-H.; Sidney, J.; Li, W.; Fusseder, N.; Sette, A. Development of an epitope conservancy analysis tool to facilitate the design of epitope-based diagnostics and vaccines. BMC Bioinform. 2007, 8, 361. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Bui, H.-H.; Sidney, J.; Dinh, K.; Southwood, S.; Newman, M.J.; Sette, A. Predicting population coverage of T-cell epitope-based diagnostics and vaccines. BMC Bioinform. 2006, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Vardhana, S.; Baldo, L.; Morice, W.G.; Wherry, E.J. Understanding T cell responses to COVID-19 is essential for informing public health strategies. Sci. Immunol. 2022, 7, eabo1303. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef]
- Pacholczyk, M.; Rieske, P. In Silico Studies Suggest T-Cell Cross-Reactivity between SARS-COV-2 and Less Dangerous Coronaviruses. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Kundu, R.; Narean, J.S.; Wang, L.; Fenn, J.; Pillay, T.; Fernandez, N.D.; Conibear, E.; Koycheva, A.; Davies, M.; Tolosa-Wright, M.; et al. Cross-reactive memory T cells associate with protection against SARS-CoV-2 infection in COVID-19 contacts. Nat. Commun. 2022, 13, 80. [Google Scholar] [CrossRef]
- Deng, H.; Yan, X.; Yuan, L. Human genetic basis of coronavirus disease 2019. Signal Transduct. Target. Ther. 2021, 6, 344. [Google Scholar] [CrossRef]
- Ferretti, A.P.; Kula, T.; Wang, Y.; Nguyen, D.M.; Weinheimer, A.; Dunlap, G.S.; Xu, Q.; Nabilsi, N.; Perullo, C.R.; Cristofaro, A.W.; et al. Unbiased Screens Show CD8+ T Cells of COVID-19 Patients Recognize Shared Epitopes in SARS-CoV-2 that Largely Reside outside the Spike Protein. Immunity 2020, 53, 1095–1107.e3. [Google Scholar] [CrossRef]
- Yoshimoto, F.K. The Proteins of Severe Acute Respiratory Syndrome Coronavirus-2 (SARS CoV-2 or n-COV19), the Cause of COVID-19. J. Protein Chem. 2020, 39, 198–216. [Google Scholar] [CrossRef]
- Aftab, S.O.; Ghouri, M.Z.; Masood, M.U.; Haider, Z.; Khan, Z.; Ahmad, A.; Munawar, N. Analysis of SARS-CoV-2 RNA-dependent RNA polymerase as a potential therapeutic drug target using a computational approach. J. Transl. Med. 2020, 18, 275. [Google Scholar] [CrossRef] [PubMed]
- Shu, T.; Huang, M.; Wu, D.; Ren, Y.; Zhang, X.; Han, Y.; Mu, J.; Wang, R.; Qiu, Y.; Zhang, D.-Y.; et al. SARS-Coronavirus-2 Nsp13 Possesses NTPase and RNA Helicase Activities That Can Be Inhibited by Bismuth Salts. Virol. Sin. 2020, 35, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.A.; Douangamath, A.; Yadzani, S.; Yosaatmadja, Y.; Aimon, A.; Brandão-Neto, J.; Dunnett, L.; Gorrie-Stone, T.; Skyner, R.; Fearon, D.; et al. Structure, mechanism and crystallographic fragment screening of the SARS-CoV-2 NSP13 helicase. Nat. Commun. 2021, 12, 4848. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.C.-C.; Laurent-Rolle, M.; Pawlak, J.B.; Wilen, C.B.; Cresswell, P. Translational shutdown and evasion of the innate immune response by SARS-CoV-2 NSP14 protein. Proc. Natl. Acad. Sci. USA 2021, 118, e2101161118. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Mao, Y.; Li, Y.; Qi, Y. BCG Vaccination: A potential tool against COVID-19 and COVID-19-like Black Swan incidents. Int. Immunopharmacol. 2022, 108, 108870. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.-U.; Seong, B.-L. Prospects on Repurposing a Live Attenuated Vaccine for the Control of Unrelated Infections. Front. Immunol. 2022, 13, 877845. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Wang, H.; Gong, W. The Potential Roles of BCG Vaccine in the Prevention or Treatment of COVID-19. Front. Biosci. 2022, 27, 157. [Google Scholar] [CrossRef]
- Lundberg, L.; Bygdell, M.; von Feilitzen, G.S.; Woxenius, S.; Ohlsson, C.; Kindblom, J.M.; Leach, S. Recent MMR vaccination in health care workers and COVID-19: A test negative case-control study. Vaccine 2021, 39, 4414–4418. [Google Scholar] [CrossRef]
- Sanchez, M.M.; Saleeb, P.; Kottilil, S.; Mathur, P. Oral Polio Vaccine to Protect against COVID-19: Out of the Box Strategies? Open Forum Infect. Dis. 2021, 8, ofab367. [Google Scholar] [CrossRef]
- Buckley, P.R.; Lee, C.H.; Pinho, M.P.; Babu, R.O.; Woo, J.; Antanaviciute, A.; Simmons, A.; Ogg, G.; Koohy, H. HLA-dependent variation in SARS-CoV-2 CD8 + T cell cross-reactivity with human coronaviruses. Immunology 2022, 166, 78–103. [Google Scholar] [CrossRef]
- Wang, F.; Huang, S.; Gao, R.; Zhou, Y.; Lai, C.; Li, Z.; Xian, W.; Qian, X.; Li, Z.; Huang, Y.; et al. Initial whole-genome sequencing and analysis of the host genetic contribution to COVID-19 severity and susceptibility. Cell Discov. 2020, 6, 83. [Google Scholar] [CrossRef]
- Tomita, Y.; Ikeda, T.; Sato, R.; Sakagami, T. Association between HLA gene polymorphisms and mortality of COVID-19: An in silico analysis. Immun. Inflamm. Dis. 2020, 8, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Koff, W.C.; Berkley, S.F. A universal coronavirus vaccine. Science 2021, 371, 759. [Google Scholar] [CrossRef] [PubMed]
- Morens, D.M.; Taubenberger, J.K.; Fauci, A.S. Universal Coronavirus Vaccines—An Urgent Need. N. Engl. J. Med. 2022, 386, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Morens, D.M.; Taubenberger, J.K.; Fauci, A.S. A Centenary Tale of Two Pandemics: The 1918 Influenza Pandemic and COVID-19, Part I. Am. J. Public Health 2021, 111, 1086–1094. [Google Scholar] [CrossRef]
- Morens, D.M.; Taubenberger, J.K.; Fauci, A.S. A Centenary Tale of Two Pandemics: The 1918 Influenza Pandemic and COVID-19, Part II. Am. J. Public Health 2021, 111, 1267–1272. [Google Scholar] [CrossRef]
- Almofti, Y.A.; Abd-Elrahman, K.A.; Eltilib, E.E.M. Vaccinomic approach for novel multi epitopes vaccine against severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2). BMC Immunol. 2021, 22, 22. [Google Scholar] [CrossRef]
- Swadling, L.; Diniz, M.O.; Schmidt, N.M.; Amin, O.E.; Chandran, A.; Shaw, E.; Pade, C.; Gibbons, J.M.; Le Bert, N.; Tan, A.T.; et al. Pre-existing polymerase-specific T cells expand in abortive seronegative SARS-CoV-2. Nature 2022, 601, 110–117. [Google Scholar] [CrossRef]
- Tan, C.C.; Owen, C.J.; Tham, C.Y.; Bertoletti, A.; van Dorp, L.; Balloux, F. Pre-existing T cell-mediated cross-reactivity to SARS-CoV-2 cannot solely be explained by prior exposure to endemic human coronaviruses. Infect. Genet. Evol. 2021, 95, 105075. [Google Scholar] [CrossRef]
- Beans, C. Researchers getting closer to a “universal” flu vaccine. Proc. Natl. Acad. Sci. USA 2022, 119, e2123477119. [Google Scholar] [CrossRef] [PubMed]
- Erbelding, E.J.; Post, D.J.; Stemmy, E.J.; Roberts, P.C.; Augustine, A.D.; Ferguson, S.; Paules, C.I.; Graham, B.S.; Fauci, A.S. A Universal Influenza Vaccine: The Strategic Plan for the National Institute of Allergy and Infectious Diseases. J. Infect. Dis. 2018, 218, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Lawton, G. Sights set on universal vaccine. New Sci. 2021, 249, 8–9. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Start Residue | Peptide | HLA Class I Allele | Immunogenicity | Population Coverage (%) |
|---|---|---|---|---|---|
| ORF1ab | 899 | WSMATYYLF | HLA-A*24:02c,HLA-A*24:07, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05, HLA-B*58:01 | 0.00742 | 89.83 |
| ORF1ab | 915 | LASHMYCSF | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | −0.12588 | 68.68 |
| ORF1ab | 1269 | LVSDIDITF | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | 0.2281 | 68.68 |
| ORF1ab | 1581 | QVVDMSMTY | HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | −0.1306 | 67.15 |
| ORF1ab | 1585 | MSMTYGQQF | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | −0.24791 | 68.68 |
| ORF1ab | 1767 | VMYMGTLSY | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05, HLA-B*44:03, HLA-B*58:01 | −0.03831 | 90.22 |
| ORF1ab | 1805 | MMSAPPAQY | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01, | 0.08593 | 68.68 |
| ORF1ab | 1822 | CASEYTGNY | HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | 0.18319 | 67.15 |
| ORF1ab | 2253 | MSNLGMPSY | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | 0.05143 | 81.53 |
| ORF1ab | 2272 | STNVTIATY | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*58:01 | −0.00384 | 75.56 |
| ORF1ab | 2322 | LVAEWFLAY | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05 | −0.27126 | 81.9 |
| ORF1ab | 2381 | RMYIFFASF | HLA-A*24:02, HLA-A*24:07, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*38:02, HLA-B*58:01 | 0.08761 | 89.49 |
| ORF1ab | 2383 | YIFFASFYY | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05 | 0.3343 | 81.9 |
| ORF1ab | 2593 | YVNTFSSTF | HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | 0.22019 | 67.15 |
| ORF1ab | 2936 | NVLEGSVAY | HLA-A*34:01, HLA-B*15:02, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05 | −0.1059 | 61.73 |
| ORF1ab | 3135 | VPFWITIAY | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05 | −0.05933 | 69.31 |
| ORF1ab | 3372 | QTFSVLACY | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*58:01 | −0.06765 | 75.56 |
| ORF1ab | 3645 | TVAYFNMVY | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:21, HLA-B*35:05 | −0.01723 | 68.69 |
| ORF1ab | 3710 | LMNVLTLVY | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | −0.01717 | 68.68 |
| ORF1ab | 3945 | SEFSSLPSY b | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01b, HLA-B*44:03c | 0.12602 | 69.92 |
| ORF1ab | 3948 | SSLPSYAAF | HLA-A*24:02, HLA-A*24:07, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | −0.22896 | 87.09 |
| ORF1ab | 4263 | STVLSFCAF | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | 0.15992 | 68.68 |
| ORF1ab | 4271 | FAVDAAKAY | HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01b, HLA-B*35:05 | 0.03414 | 73.5 |
| ORF1ab | 4757 | LSFKELLVY | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | 0.08254 | 68.68 |
| ORF1ab | 4904 | RLYYDSMSY a | HLA-A*11:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | 0.06159 | 73.23 |
| ORF1ab | 5057 | MVMCGGSLY | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | −0.12346 | 77.56 |
| ORF1ab | 5245 | LMIERFVSL | HLA-A*02:01c, HLA-A*02:03, HLA-B*15:02, HLA-B*15:21, HLA-B*38:02 | −0.11314 | 54.17 |
| ORF1ab | 5267 | EYADVFHLY | HLA-A*24:02, HLA-A*24:07, HLA-A*33:03, HLA-A*34:01, HLA-B*18:01 | 0.00446 | 85.01 |
| ORF1ab | 5532 | VVYRGTTTY a | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | −0.21535 | 81.53 |
| ORF1ab | 5678 | YVFCTVNAL a | HLA-A*02:01, HLA-A*02:03, HLA-A*34:01, HLA-B*15:02, HLA-B*15:21, HLA-B*35:05, HLA-B*38:02 | 0.24593 | 69.29 |
| ORF1ab | 5980 | SMMGFKMNY | HLA-A*11:01, HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | −0.11462 | 77.56 |
| ORF1ab | 6153 | HSIGFDYVY | HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05, HLA-B*58:01 | −0.21438 | 78.69 |
| ORF1ab | 6424 | MMISAGFSL a | HLA-A*02:01c, HLA-A*02:03, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*38:02 | 0.38891 | 74.8 |
| ORF1ab | 6433 | WVYKQFDTY | HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | 0.06486 | 67.15 |
| ORF1ab | 7000 | EAFLIGCNY | HLA-A*34:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | 0.07781 | 67.15 |
| ORF1ab | 7019 | YVMHANYIF | HLA-A*24:02, HLA-A*24:07, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | −0.12171 | 89.49 |
| ORF2 (S) | 83 | LPFNDGVYF | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05 | 0.11767 | 69.31 |
| ORF2 (S) | 159 | YSSANNCTF | HLA-A*24:07, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | −0.04954 | 80.83 |
| ORF2 (S) | 161 | SANNCTFEY | HLA-A*11:01, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | 0.13273 | 77.97 |
| ORF2 (S) | 268 | YLQPRTFLL a | HLA-A*02:01c, HLA-A*02:03, HLA-A*24:02a, HLA-A*24:07, HLA-B*38:02 | 0.1305 | 74.4 |
| ORF2 (S) | 368 | YNSASFSTF | HLA-A*24:02, HLA-A*24:07, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05 | −0.18217 | 89.49 |
| ORF2 (S) | 686 | VASQSIIAY | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | −0.0709 | 68.68 |
| ORF2 (S) | 690 | SIIAYTMSL a | HLA-A*02:01c, HLA-A*02:03, HLA-A*34:01, HLA-B*15:21, HLA-B*38:02 | −0.12935 | 48.2 |
| ORF2 (S) | 868 | MIAQYTSAL a | HLA-A*02:03b, HLA-A*34:01, HLA-B*15:02, HLA-B*15:21, HLA-B*35:05 | −0.18768 | 57.95 |
| ORF2 (S) | 895 | IPFAMQMAY | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05 | −0.32801 | 69.31 |
| ORF5 | 169 | VATSRTLSY b | HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | −0.17295 | 68.68 |
| ORF9 | 265 | KAYNVTQAF b | HLA-A*24:02, HLA-A*24:07, HLA-B*15:02, HLA-B*15:13, HLA-B*15:21, HLA-B*35:05, HLA-B*58:01 | −0.00587 | 87.09 |
| Protein | Start Residue | Peptide | HLA Class I Allele | Immunogenicity | Population Coverage (%) |
|---|---|---|---|---|---|
| ORF1ab | 44 | HLKDGTCGL | HLA-A*02:03 | 0.40234 | 7.68 |
| ORF1ab | 1634 | YHTTDPSFL | HLA-B*38:02 | 0.44289 | 10.65 |
| ORF1ab | 2001 | TYKPNTWCI | HLA-A*24:02 b; HLA-A*24:07 | 0.5572 | 58.77 |
| ORF1ab | 2789 | ITPVHVMSK | HLA-A*11:01 b | 0.43221 | 29.64 |
| ORF1ab | 3541 | RTILGSALL | HLA-B*58:01 | 0.45285 | 11.11 |
| ORF1ab | 3640 | LPSLATVAY | HLA-B*15:02, HLA-B*15:21, HLA-B*18:01, HLA-B*35:05 | 0.35609 | 55.67 |
| ORF1ab | 4580 | TVQFCDAMR | HLA-A*33:03 | 0.37218 | 29.13 |
| ORF1ab | 6158 | DYVYNPFMI | HLA-A*24:02 b, HLA-A*24:07 | 0.56221 | 58.77 |
| ORF1ab | 6819 | LEKCDLQNY | HLA-B*18:01, HLA-B*44:03 | 0.37101 | 28.15 |
| ORF2 (S) | 1098 | GTHWFVTQR | HLA-A*11:01 c, HLA-A*33:03 b | 0.35133 | 53.67 |
| ORF2 (S) | 1208 | YIKWPWYIW | HLA-B*58:01 | 0.42524 | 11.11 |
| ORF2 (S) | 1211 | WPWYIWLGF | HLA-B*18:01, HLA-B*35:05 | 0.41673 | 27.22 |
| ORF5 (M) | 25 | FLFLTWICL | HLA-A*02:01 a | 0.35397 | 13.07 |
| ORF9 (N) | 103 | LSPRWYFYY | HLA-A*24:07, HLA-B*58:01 | 0.35734 | 45.59 |
| Protein | Start Residue | Peptide | HLA Class II Allele | IFN-γ Score | Population Coverage (%) |
|---|---|---|---|---|---|
| ORF1ab | 457 | EKVNINIVGDFKLNE | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.073961117 | 87.72 |
| ORF1ab | 458 | KVNINIVGDFKLNEE c | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.10696324 | 87.72 |
| ORF1ab | 734 | KAPKEIIFLEGETLP | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.24929914 | 87.72 |
| ORF1ab | 735 | APKEIIFLEGETLPT | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.082149378 | 87.72 |
| ORF1ab | 736 | PKEIIFLEGETLPTE c | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.077069314 | 87.72 |
| ORF1ab | 1053 | KPTVVVNAANVYLKH | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.45376782 | 87.72 |
| ORF1ab | 1171 | TNVYLAVFDKNLYDK | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.32342333 | 91.86 |
| ORF1ab | 1349 | CKSAFYILPSIISNE | HLA-DRB1*11:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.28976547 | 86.38 |
| ORF1ab | 1350 | KSAFYILPSIISNEK a,c | HLA-DRB1*11:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.33784834 | 86.38 |
| ORF1ab | 1363 | EKQEILGTVSWNLRE | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.0096068911 | 65.21 |
| ORF1ab | 1419 | GARFYFYTSKTTVAS | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.48814565 | 65.21 |
| ORF1ab | 1799 | QQESPFVMMSAPPAQ | HLA-DRB1*11:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.31609633 | 86.38 |
| ORF1ab | 1800 | QESPFVMMSAPPAQY c | HLA-DRB1*11:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.15478331 | 86.38 |
| ORF1ab | 1801 | ESPFVMMSAPPAQYE a | HLA-DRB1*11:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.21040286 | 86.38 |
| ORF1ab | 1802 | SPFVMMSAPPAQYELc | HLA-DRB1*11:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.18691649 | 86.38 |
| ORF1ab | 1908 | TEQPIDLVPNQPYPN c | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.18762311 | 87.72 |
| ORF1ab | 1909 | EQPIDLVPNQPYPNA | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.15613773 | 87.72 |
| ORF1ab | 2211 | LEASFNYLKSPNFSK a | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.12722255 | 91.86 |
| ORF1ab | 2212 | EASFNYLKSPNFSKL c | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.37773341 | 91.86 |
| ORF1ab | 2518 | LSHFVNLDNLRANNT c | HLA-DRB1*11:01, HLA-DRB1*12:02, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.29350215 | 86.38 |
| ORF1ab | 2958 | DGSIIQFPNTYLEGS | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.087720744 | 87.72 |
| ORF1ab | 3150 | TKHFYWFFSNYLKRR c | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.34583424 | 65.21 |
| ORF1ab | 3816 | STQEFRYMNSQGLLP | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.28198599 | 94.14 |
| ORF1ab | 3817 | TQEFRYMNSQGLLPP | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.41760878 | 94.14 |
| ORF1ab | 3818 | QEFRYMNSQGLLPPK | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.25437902 | 94.14 |
| ORF1ab | 4559 | ENPDILRVYANLGER | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.011725622 | 87.72 |
| ORF1ab | 4560 | NPDILRVYANLGERV | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.22993566 | 87.72 |
| ORF1ab | 4561 | PDILRVYANLGERVR a | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.26161469 | 87.72 |
| ORF1ab | 5018 | MPNMLRIMASLVLAR c | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.32014953 | 87.72 |
| ORF1ab | 5019 | PNMLRIMASLVLARK a | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.39136883 | 87.72 |
| ORF1ab | 5166 | GLVASIKNFKSVLYY a | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.43106149 | 87.72 |
| ORF1ab | 5167 | LVASIKNFKSVLYYQ | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.093707023 | 87.72 |
| ORF1ab | 5168 | VASIKNFKSVLYYQN | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.29825337 | 87.72 |
| ORF1ab | 5169 | ASIKNFKSVLYYQNN | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.25775961 | 87.72 |
| ORF1ab | 5450 | CTERLKLFAAETLKA | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.10920914 | 87.72 |
| ORF1ab | 5451 | TERLKLFAAETLKAT | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.013340883 | 94.14 |
| ORF1ab | 5769 | PAEIVDTVSALVYDN c | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.31287444 | 65.21 |
| ORF1ab | 5845 | VASKILGLPTQTVDS | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.015678357 | 87.72 |
| ORF1ab | 5846 | ASKILGLPTQTVDSS | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.12146879 | 87.72 |
| ORF1ab | 6002 | EEAIRHVRAWIGFDV c | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.42312165 | 87.72 |
| ORF1ab | 6039 | GVNLVAVPTGYVDTP | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.46784679 | 94.14 |
| ORF1ab | 6087 | VRIKIVQMLSDTLKN | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.08846654 | 87.72 |
| ORF1ab | 6516 | KPVPEVKILNNLGVD c | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.092818209 | 87.72 |
| ORF1ab | 6517 | PVPEVKILNNLGVDI | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.0068847756 | 87.72 |
| ORF1ab | 6518 | VPEVKILNNLGVDIA | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.25937664 | 87.72 |
| ORF1ab | 6519 | PEVKILNNLGVDIAA | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.41267881 | 87.72 |
| ORF1ab | 6750 | LLDDFVEIIKSQDLS c | HLA-DRB1*07:01, HLA-DRB1*11:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.27733231 | 68.03 |
| ORF1ab | 6751 | LDDFVEIIKSQDLSV a | HLA-DRB1*07:01, HLA-DRB1*11:01, HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.04841137 | 95.26 |
| ORF1ab | 6752 | DDFVEIIKSQDLSVV c | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.28882695 | 94.14 |
| ORF2 (S) | 307 | TVEKGIYQTSNFRVQ c | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.45017873 | 65.21 |
| ORF2 (S) | 308 | VEKGIYQTSNFRVQP b | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.20827026 | 65.21 |
| ORF2 (S) | 309 | EKGIYQTSNFRVQPT b,c | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.46467579 | 65.21 |
| ORF2 (S) | 310 | KGIYQTSNFRVQPTE b | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.36164402 | 65.21 |
| ORF4 (E) | 58 | VYSRVKNLNSSRVPD | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.18705221 | 87.72 |
| ORF4 (E) | 59 | YSRVKNLNSSRVPDL c | HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.45329667 | 87.72 |
| ORF5 (M) | 163 | DLPKEITVATSRTLS | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.36193098 | 65.21 |
| ORF5 (M) | 164 | LPKEITVATSRTLSY | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | −0.17020992 | 94.14 |
| ORF5 (M) | 165 | PKEITVATSRTLSYY a | HLA-DRB1*07:01, HLA-DRB1*12:02, HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.2121047 | 94.14 |
| Protein | Start Residue | Peptide | HLA Class II Allele | IFN-γ Score | Population Coverage (%) |
|---|---|---|---|---|---|
| ORF9 (N) | 304 | IAQFAPSASAFFGMS b | HLA-DRB1*07:01 | 1.0616777 | 20.49 |
| ORF9 (N) | 309 | PSASAFFGMSRIGME | HLA-DRB1*11:01 | 0.94891009 | 4.83 |
| ORF9 (N) | 303 | QIAQFAPSASAFFGM | HLA-DRB1*07:01 | 0.83655244 | 20.49 |
| ORF1ab | 7076 | GRLIIRENNRVVISS | HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.79853065 | 51.25 |
| ORF9 (N) | 310 | SASAFFGMSRIGMEV | HLA-DRB1*11:01 | 0.79707565 | 4.83 |
| ORF1ab | 7082 | ENNRVVISSDVLVNN b | HLA-DRB1*07:01, HLA-DRB1*15:01, HLA-DRB1*15:02 | 0.79231845 | 61.9 |
| ORF1ab | 7075 | KGRLIIRENNRVVIS | HLA-DRB1*15:01, HLA-DRB1*15:02, HLA-DRB1*16:02 | 0.78951146 | 51.25 |
| ORF1ab | 4124 | WPLIVTALRANSAVK | HLA-DRB1*11:01, HLA-DRB1*1202 | 0.78569358 | 60.59 |
| ORF1ab | 1631 | AFEYYHTTDPSFLGR | HLA-DRB1*07:01 | 0.77956564 | 20.49 |
| ORF1ab | 1630 | EAFEYYHTTDPSFLG b | HLA-DRB1*07:01 | 0.7473454 | 20.49 |
| ORF9 (N) | 302 | PQIAQFAPSASAFFG | HLA-DRB1*07:01 | 0.72358869 | 20.49 |
| ORF2 (S) | 24 | LPPAYTNSFTRGVYY | HLA-DRB1*07:01 | 0.71760044 | 20.49 |
| ORF2 (S) | 306 | FTVEKGIYQTSNFRV a,b | HLA-DRB1*07:01 a | 0.70919003 | 20.49 |
| ORF2 (S) | 681 | PRRARSVASQSIIAY b | HLA-DRB1*07:01 | 0.67693256 | 20.49 |
| ORF1ab | 391 | SGLKTILRKGGRTIA | HLA-DRB1*11:01 | 0.67477291 | 4.83 |
| Epitope Sequence | Immunogenicity | Identity (%) | HLA that Binds | Population Coverage (%) | Position | Location |
|---|---|---|---|---|---|---|
| HEFCSQHTM | 0.03852 | 100 | HLA-B*18:01, HLA-B*38:02, HLA-B*44:03 | 37.13 | 5202—5210 |
|
| SLAIDAYPL | 0.0619 | 100 | HLA-A*02:01 a | 13.07 | 5254—5262 |
|
| Start | Epitope Sequence | Alpha B.1.1.7 (158) | Beta B.1.351 (374) | Delta B.1.617.2 (1157) | Gamma P.1 (9) | Omicron BA.1.1 (884) |
|---|---|---|---|---|---|---|
| 5202 | HEFCSQHTM | 100.00% (111/111) | 100.00% (146/146) | 100.00% (458/458) | 100.00% (8/8) | 100.00% (119/119) |
| 5254 | SLAIDAYPL | 100.00% (111/111) | 100.00% (146/146) | 100.00% (458/458) | 100.00% (8/8) | 100.00% (119/119) |
| SARS-CoV-2 | SARS-CoV | MERS-CoV |
|---|---|---|
| 5202HEFCSQHTM5210 | 5179HEFCSQHTM5187 | 5188HEFCSQHTL5196 |
| 5254SLAIDAYPL5262 | 5230SLAIDAYPL5238 | 5239SLAIDAYPL5247 |
| Epitope Sequence [Domain Function] | HLA that Binds [Population Coverage] | 229E [Position] | NL63 [Position] | HKU1 [Position] | OC43 [Position] |
|---|---|---|---|---|---|
| 5005HLMGWDYPK5013 [RdRp (nsp12), Catalytic residue, Putative inhibitor binding site, Conserved polymerase binding A] | HLA-A*11:01 a HLA-A*33:03 HLA-A*34:01 (62.81) | KLMGWDYPK c (4676–4684) | MLMGWDYPK b (4651–4659) | VLMGWDYPK c (5066–5074) | VLMGWDYPK c (4978–4986) |
| 5251FVSLAIDAY5259 [RdRp (nsp12), Putative RNA binding site] | HLA-A*34:01 HLA-B*15:02 HLA-B*15:21 HLA-B*35:05 (54.16) | YVSLAIDAY b (4922–4930) | YVSLAIDAY b (4897–4905) | FVSLAIDAY b (5312–5320) | FVSLAIDAY b (5224–5232) |
| 6321SIVCRFDTR6329 (Nsp14, N7-Mtase Active site) | HLA-A*33:03 (29.13) | SIVCRFDTR b (5981–5989) | SIVCRFDTR b (5956–5964) | SIVCRFDTR b (6378–6386) | AVVCRFDTR (6290–6298) |
| 6844KYTQLCQYL6852 (Coronavirus nsp13 super family) | HLA-A*24:02 a HLA-A*24:07 (58.77) | KYTQLCQYF b (6503–6511) | KYTQLCQYL b (6474–6482) | KYTQLCQYL b (6929–6937) | KYTQLCQYL b (6842–6850) |
| Epitope Sequence [Position] [Domain Function] | HLA [Population Coverage] | 229E [Position] | NL63 [Position] | HKU1 [Position] | OC43 [Position] |
|---|---|---|---|---|---|
|
MNLKYAISAKNRART a [4934–4948] [RdRp (nsp12), Putative RNA binding site, Putative inhibitor binding site, Putative nsp7 interaction site, Conserved polymerase binding F] | DRB1*11:01 [4.83] | LNLKYAISGKERART b [4605–4619] | LNLKYAISGKERART b [4580–4594] | MNLKYAISAKNRART b [4995–5009] | MNLKYAISAKNRART b [4907–4921] |
|
KAVFISPYNSQNAVA [5832–5846] [Nsp13, Helicase domain] | DRB1*15:01 DRB1*15:02 DRB1*16:02 [51.25] | KAVFISPYNSQNYVA c [5504–5518] | KAVFISPYNSQNYVA c [5479–5493] | SAVFISPYNSQNYVA c [5892–5906] | KAVFISPYNSQNFAA c [5804–5818] |
|
AVFISPYNSQNAVAS
[5833–5847] [Nsp13, Helicase domain] | DRB1*15:01 DRB1*15:02 DRB1*16:02 [51.25] | AVFISPYNSQNYVAAc [5505–5519] | AVFISPYNSQNYVAS c [5480–5494] | AVFISPYNSQNYVAK c [5893–5907] | AVFISPYNSQNFAAK c [5805–5819] |
|
VFISPYNSQNAVASK a [5834–5848] [Nsp13, Helicase domain] | DRB1*15:01 DRB1*15:02 DRB1*16:02 [51.25] | VFISPYNSQNYVAAR [5506–5520] | VFISPYNSQNYVASR [5481–5495] | VFISPYNSQNYVAKRc [5894–5908] | VFISPYNSQNFAAKRc [5806–5820] |
| NCNVDRYPANSIVCR [6311–6325] [Nsp14, N7-Mtase active site] | DRB1*15:01 DRB1*15:02 [47.35] | NCNVDMYPEFSIVCR [5971–5985] | NCNVDMYPEFSIVCR [5946–5960] | NCNVDKYPSNSIVCR b [6368–6382] | NCNVDKYPPNAVVCR [6280–6294] |
| SARS-CoV-2 Peptide | BCG Peptide | Protein Name in SARS-CoV-2 | Protein Name in BCG | HLA Allele | Predicted HLA Binding Affinity a | |
|---|---|---|---|---|---|---|
| SARS-CoV-2 | BCG | |||||
| FVFLVLLPL | FVFLALLPF | ORF2 | transmembrane ABC transporter ATP-binding protein | HLA-A*02:01 | 4.15 | n.b |
| HLA-A*34:01 | 10.54 | 11.56 | ||||
| HLA-B*15:02 | n.b | 3.92 | ||||
| HLA-B*15:13 | n.b | 4.93 | ||||
| HLA-B*15:21 | n.b | 4.98 | ||||
| HLA-B*18:01 | n.b | 9.84 | ||||
| HLA-B*35:05 | n.b | 4.91 | ||||
| WLIVGVALL | WLMVGIAIL | ORF3a | NUDIX domain-containing protein | HLA-A*02:01 | 3.97 | 3.53 |
| HLA-A*02:03 | 3.55 | 3.63 | ||||
| HLA-B*38:02 | n.b | 13.42 | ||||
| FLIVGIELL | ORF3a | hypothetical protein N054_02474 | HLA-A*02:01 | 3.97 | 2.96 | |
| HLA-A*02:03 | 3.55 | 2.97 | ||||
| WLIAGVVFL | ORF3a | membrane protein | HLA-A*02:01 | 3.97 | 2.78 | |
| HLA-A*02:03 | 3.55 | 3.06 | ||||
| WLIVGYFAV | ORF3a | MFS transporter | HLA-A*02:01 | 3.97 | 2.74 | |
| HLA-A*02:03 | 3.55 | 2.99 | ||||
| SVLLFLAFV | YVLLFVSLV | ORF4 | cytochrome c biogenesis protein | HLA-A*02:01 | 3.34 | 3.72 |
| KLLEQWNLV | KLLEEWLAM | ORF5 | Isocitrate lyase | HLA-A*02:01 | 2.87 | 3.74 |
| HLVDFQVTI | HLVDFPVTL | ORF6 | RD1 region associated protein | HLA-A*02:01 | 4.40 | 4.41 |
| SLVDSQVTV | ORF6 | transmembrane transport protein MmpL7 | HLA-A*02:01 | 4.40 | 3.99 | |
| HLA-A*02:03 | n.b | 3.52 | ||||
| FLAFLLFLV | FLAFLDSTI | ORF7b | MFS transporter | HLA-A*02:01 | 2.34 | 3.68 |
| HLA-A*02:03 | 2.48 | 2.97 | ||||
| FYSKWYIRV | GYEKWRIRF | ORF8 | SAM-dependent methyltransferase | HLA-A*24:02 | 6.18 | 7.60 |
| HLA-A*24:07 | 7.83 | 10.09 | ||||
| LLLDRLNQL | YLLDRLAEL | ORF9 | alpha-keto acid decarboxylase family protein | HLA-A*02:01 | 3.06 | 2.29 |
| HLA-A*02:03 | 3.50 | 2.45 | ||||
| LLLDRLPNL | ORF9 | cytochrome P450 | HLA-A*02:01 | 3.06 | 2.81 | |
| HLA-A*02:03 | 3.50 | 3.62 | ||||
| LLRDRLAQL | ORF9 | inositol monophosphatase | HLA-A*02:01 | 3.06 | n.b | |
| HLA-A*02:03 | 3.50 | 3.26 | ||||
| SARS-CoV-2 | BCG | Gene Name in SARS-CoV-2 | Protein Name in BCG | HLA Allele | Predicted HLA Binding Affinity * | |
|---|---|---|---|---|---|---|
| SARS-CoV-2 | BCG | |||||
| VKHVYQLRARSVSPK | TDLLYQLASRSVSST | ORF7a | amidase | HLA-DRB1*11:01 | 3.64 | 2.89 |
| HLA-DRB1*12:02 | 4.38 | - | ||||
| MMR | SARS-CoV-2 Peptides | MMR Peptides | Protein Name in SARS-CoV-2 | Protein Name in MMR | HLA Allele | Predicted HLA Binding Affinity a | |
|---|---|---|---|---|---|---|---|
| SARS-CoV-2 | MMR | ||||||
| Measles | SVLLFLAFV | YVLLAVLFV | ORF4 | hemagglutinin | HLA-A*02:01 | 3.34 | 3.77 |
| ILFLALITL | VMFLSLIGL | ORF7a | hemagglutinin | HLA-A*02:01 | 4.75 | 3.97 | |
| HLA-A*02:03 | - | 4.31 | |||||
| LLLDRLNQL | RLLDRLVRL | ORF9 | nucleocapsid protein | HLA-A*02:01 | 3.96 | 3.66 | |
| HLA-A*02:03 | 3.50 | - | |||||
| Mumps | FVFLVLLPL | FLLLILLPL | ORF2 | small hydrophobic protein | HLA-A*02:01 | 4.15 | 3.58 |
| HLA-A*34:01 | 10.54 | - | |||||
| SVLLFLAFV | SLLLFLLYL | ORF4 | small hydrophobic protein | HLA-A*02:01 | 3.34 | 4.67 | |
| FLAFLLFLV | FLTFLLLIV | ORF7b | small hydrophobic protein | HLA-A*02:01 | 2.34 | 3.82 | |
| HLA-A*02:03 | 2.48 | 3.73 | |||||
| Rubella | LLLDRLNQL | ALLDRLRGV | ORF9 | nonstructural capsid protein, partial | HLA-A*02:01 | 3.06 | 4.84 |
| HLA-A*02:03 | 3.50 | 4.84 | |||||
| Epitope Sequence | HLA that Binds | Immunogenicity | Population Coverage of Individual Epitope (%) | Similarity to Human Peptides |
|---|---|---|---|---|
| HEFCSQHTM | HLA-B*18:01, HLA-B*38:02, HLA-B*44:03 | 0.03852 | 37.13 | None |
| SLAIDAYPL | HLA-A*02:01 | 0.0619 | 13.07 | None |
| HLMGWDYPK | HLA-A*11:01, HLA-A*33:03, HLA-A*34:01 | 0.1053 | 62.81 | None |
| FVSLAIDAY | HLA-A*34:01, HLA-B*15:02, HLA-B*15:21, HLA-B*35:05 | 0.00813 | 54.16 | None |
| KYTQLCQYL | HLA-A*24:02, HLA-A*24:07 | −0.06866 | 58.77 | None |
| MNLKYAISAKNRART | DRB1*11:01 | 0.40189009 | 4.83 | None |
| KAVFISPYNSQNAVA | DRB1*15:01, DRB1*15:02, DRB1*16:02 | 0.4720961 | 51.25 | None |
| AVFISPYNSQNAVAS | DRB1*15:01, DRB1*15:02, DRB1*16:02 | 0.53704543 | 51.25 | None |
| VFISPYNSQNAVASK | DRB1*15:01, DRB1*15:02, DRB1*16:02 | 0.22363359 | 51.25 | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gustiananda, M.; Julietta, V.; Hermawan, A.; Febriana, G.G.; Hermantara, R.; Kristiani, L.; Sidhartha, E.; Sutejo, R.; Agustriawan, D.; Andarini, S.; et al. Immunoinformatics Identification of the Conserved and Cross-Reactive T-Cell Epitopes of SARS-CoV-2 with Human Common Cold Coronaviruses, SARS-CoV, MERS-CoV and Live Attenuated Vaccines Presented by HLA Alleles of Indonesian Population. Viruses 2022, 14, 2328. https://doi.org/10.3390/v14112328
Gustiananda M, Julietta V, Hermawan A, Febriana GG, Hermantara R, Kristiani L, Sidhartha E, Sutejo R, Agustriawan D, Andarini S, et al. Immunoinformatics Identification of the Conserved and Cross-Reactive T-Cell Epitopes of SARS-CoV-2 with Human Common Cold Coronaviruses, SARS-CoV, MERS-CoV and Live Attenuated Vaccines Presented by HLA Alleles of Indonesian Population. Viruses. 2022; 14(11):2328. https://doi.org/10.3390/v14112328
Chicago/Turabian StyleGustiananda, Marsia, Vivi Julietta, Angelika Hermawan, Gabriella Gita Febriana, Rio Hermantara, Lidya Kristiani, Elizabeth Sidhartha, Richard Sutejo, David Agustriawan, Sita Andarini, and et al. 2022. "Immunoinformatics Identification of the Conserved and Cross-Reactive T-Cell Epitopes of SARS-CoV-2 with Human Common Cold Coronaviruses, SARS-CoV, MERS-CoV and Live Attenuated Vaccines Presented by HLA Alleles of Indonesian Population" Viruses 14, no. 11: 2328. https://doi.org/10.3390/v14112328
APA StyleGustiananda, M., Julietta, V., Hermawan, A., Febriana, G. G., Hermantara, R., Kristiani, L., Sidhartha, E., Sutejo, R., Agustriawan, D., Andarini, S., & Parikesit, A. A. (2022). Immunoinformatics Identification of the Conserved and Cross-Reactive T-Cell Epitopes of SARS-CoV-2 with Human Common Cold Coronaviruses, SARS-CoV, MERS-CoV and Live Attenuated Vaccines Presented by HLA Alleles of Indonesian Population. Viruses, 14(11), 2328. https://doi.org/10.3390/v14112328