

Characterization of Grapevine Fanleaf Virus Isolates in ‘Chardonnay’ Vines Exhibiting Severe and Mild Symptoms in Two Vineyards

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Vineyard Sites
2.2. Disease Symptom Monitoring, Grapevine Leaf Sampling, and GFLV Detection
2.3. Statistical Analyses of Qualitative and Quantitative Phenotypic Traits
2.4. RNA Sequencing and Downstream Bioinformatics Analyses
2.5. GFLV Sequences Analyses, Genetic Diversity and Recombination Detection
3. Results
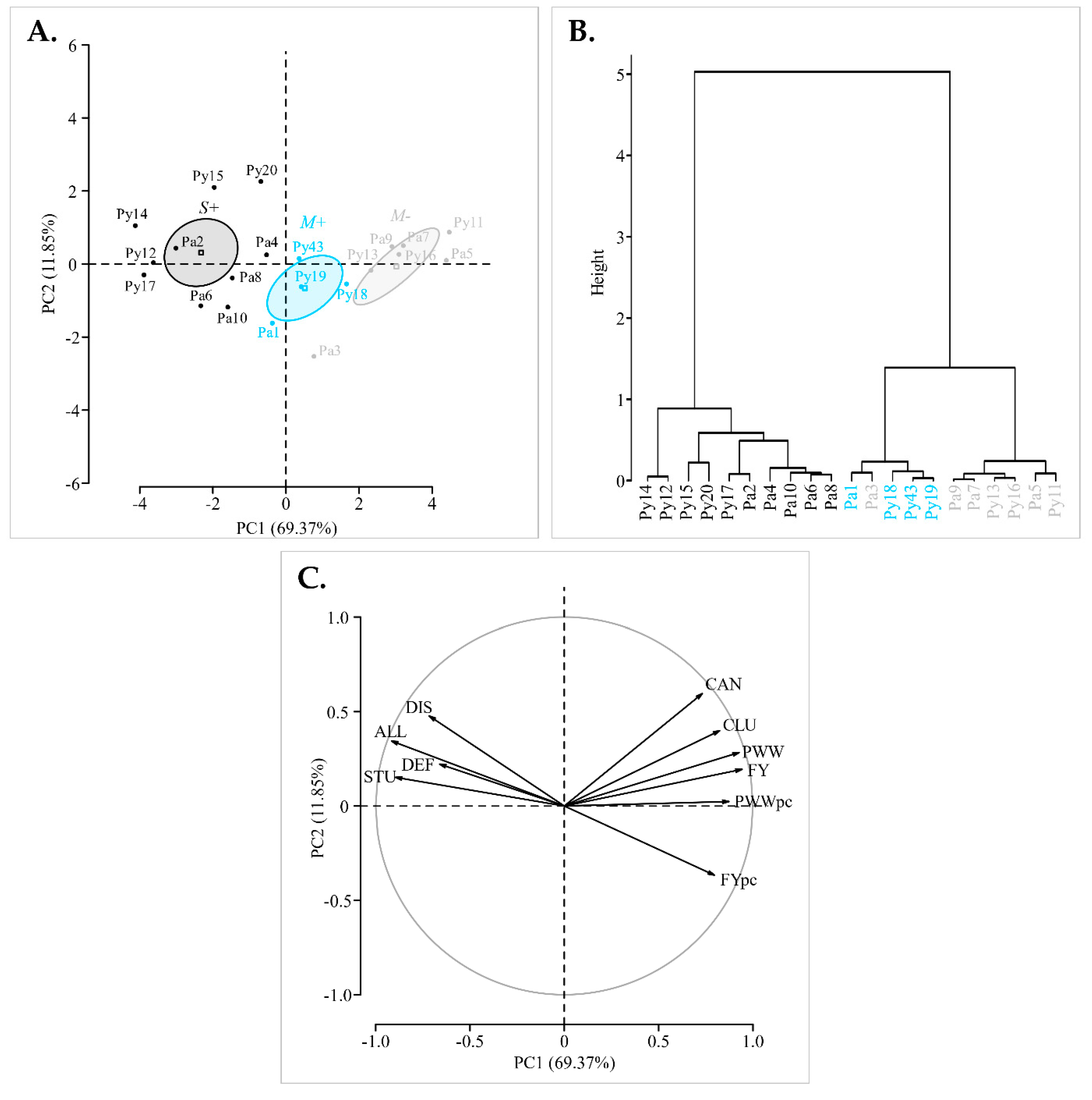
3.1. Identification of ‘Chardonnay’ Vines Showing Distinct Fanleaf Symptoms
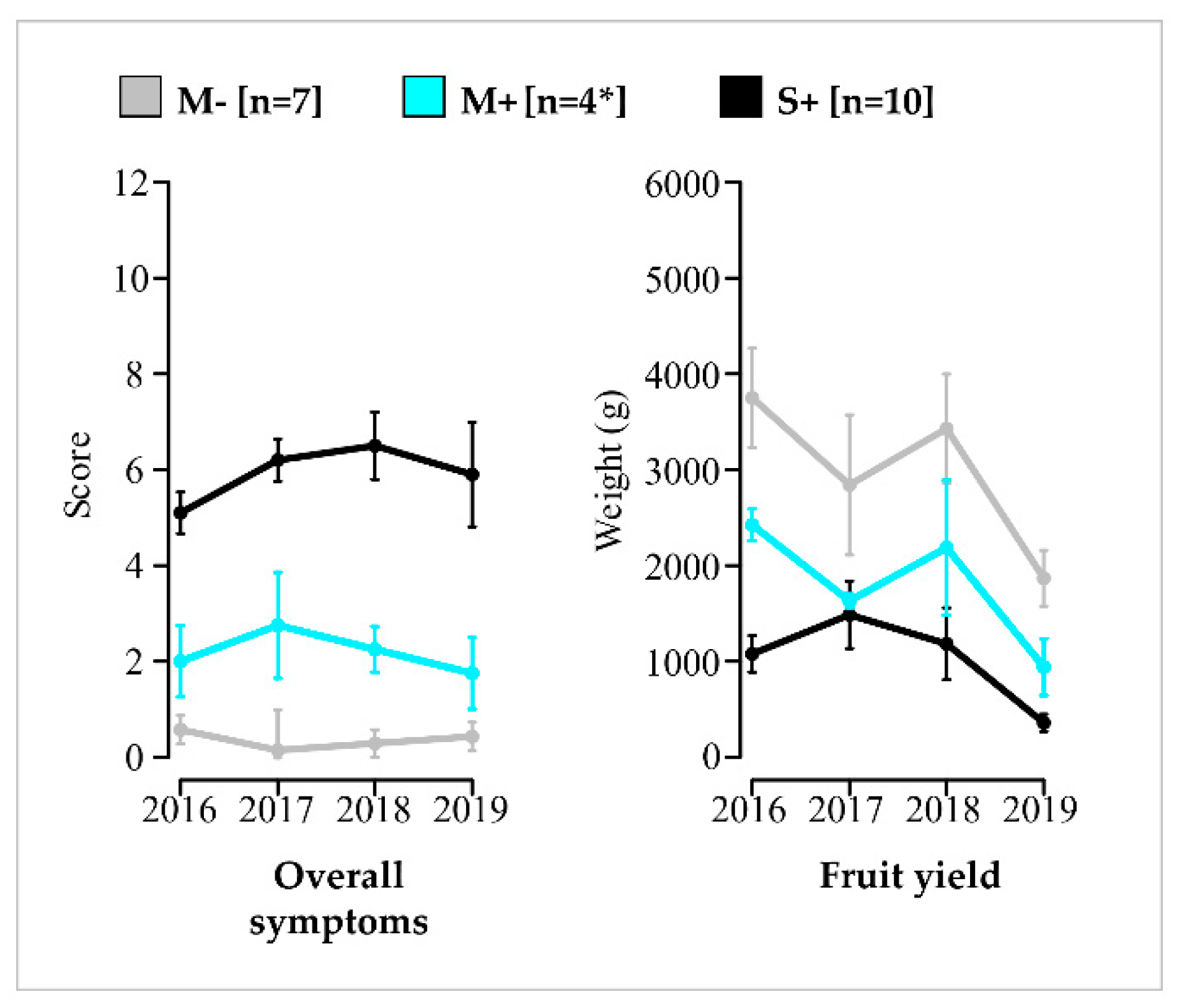
3.2. Phenotypic Differentiation and Overtime Stability of M−, M+ and S+ Categories
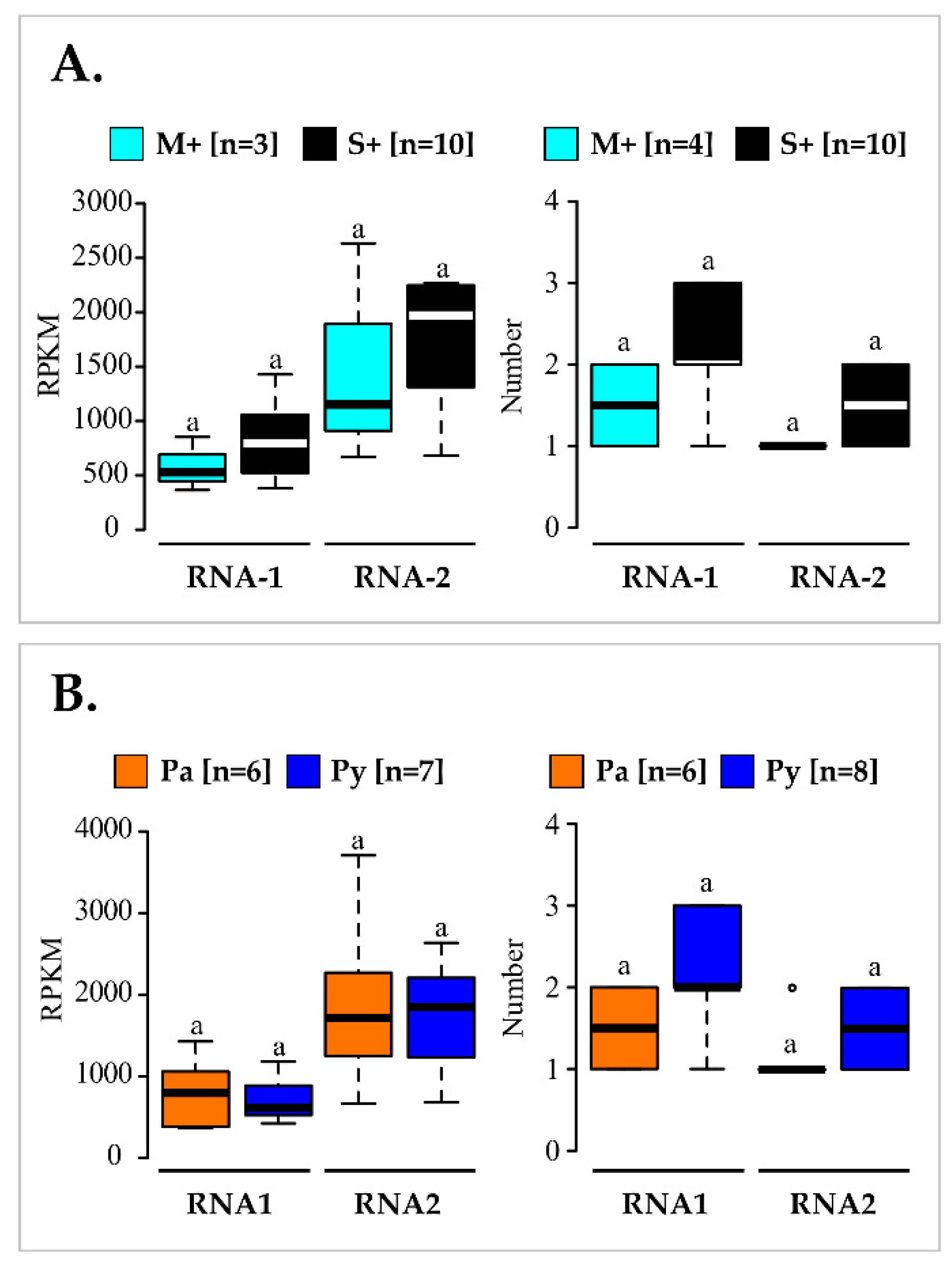
3.3. Virome Description and Genetic Characterization of GFLV Isolates: Mixed Infections of Highly Divergent Molecular Variants
3.4. Comparison of GFLV Molecular Variants in the M+ and S+ Categories and in Pa and Py Vineyard Sites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martelli, G.P. Virus Diseases of Grapevine; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014. [Google Scholar] [CrossRef]
- Martelli, G.P. An overview on grapevine viruses, viroids, and the diseases they cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 31–46. [Google Scholar]
- Fuchs, M. Grapevine viruses: A multitude of diverse species with simple but overall poorly adopted management solutions in the vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Andret-Link, P.; Schmitt-Keichinger, C.; Demangeat, G.; Komar, V.; Fuchs, M. The specific transmission of Grapevine fanleaf virus by its nematode vector Xiphinema index is solely determined by the viral coat protein. Virology 2004, 320, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Demangeat, G.; Esmenjaud, D.; Voisin, R.; Bidault, J.-M.; Grenan, S.; Claverie, M. Le court-noué de la vigne: État des connaissances sur cette maladie. Phytoma 2005, 587, 38–42. [Google Scholar]
- Schmitt-Keichinger, C.; Hemmer, C.; Berthold, F.; Ritzenthaler, C. Molecular, cellular, and structural biology of Grapevine fanleaf virus. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 83–107. [Google Scholar]
- Legin, R. Expérimentation pour étudier l’effet des principales viroses sur la végétation et la production de la vigne. Ann. Phytopathol. 1972, 2, 49–57. [Google Scholar]
- Legin, R.; Bass, P.; Etienne, L.; Fuchs, M. Selection of mild virus strains of fanleaf degeneration by comparative field performance of infected grapevines. Vitis 1993, 32, 103–110. [Google Scholar]
- Mannini, F.; Digiaro, M. The effects of viruses and viral diseases on grapes and wine. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 453–482. [Google Scholar]
- Vuittenez, A. Variation des symptômes de la dégénérescence infectieuse de la vigne. Interprétation d’expériences de transmission de la maladie par greffage. Comptes Rendus Séances L’académie Sci. 1956, 243, 515–517. [Google Scholar]
- Vigne, E.; Komar, V.; Tannière, M.; Demangeat, G.; Duchêne, E.; Steyer, D.; Lemarquis, G.; Ritzenthaler, C.; Lemaire, O.O. Comparative pathogenic effects of distinct Grapevine fanleaf virus strains on Vitis vinifera cvs Gewurztraminer and Chardonnay. In Proceedings of the 18th Congress of the International Council for the Study of Virus and Virus-Like Diseases of the Grapevine (ICVG), Ankara, Turkey, 7–11 September 2015; pp. 236–237. [Google Scholar]
- Digiaro, M.; Elbeaino, T.; Martelli, G.P. Grapevine fanleaf virus and other old world nepoviruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 47–82. [Google Scholar]
- Martin, I.R.; Vigne, E.; Velt, A.; Hily, J.-M.; Garcia, S.; Baltenweck, R.; Komar, V.; Rustenholz, C.; Hugueney, P.; Lemaire, O.; et al. Severe stunting symptoms upon Nepovirus infection are reminiscent of a chronic hypersensitive-like response in a perennial woody fruit crop. Viruses 2021, 13, 2138. [Google Scholar] [CrossRef]
- Vigne, E.; Gottula, J.; Schmitt-Keichinger, C.; Komar, V.; Ackerer, L.; Belval, L.; Rakotomalala, L.; Lemaire, O.; Ritzenthaler, C.; Fuchs, M. A strain-specific segment of the RNA-dependent RNA polymerase of Grapevine fanleaf virus determines symptoms in Nicotiana species. J. Gen. Virol. 2013, 94, 2803–2813. [Google Scholar] [CrossRef]
- Martin, I.R.; Vigne, E.; Berthold, F.; Komar, V.; Lemaire, O.; Fuchs, M.; Schmitt-Keichinger, C. The 50 distal amino acids of the 2A(HP) homing protein of Grapevine fanleaf virus elicit a hypersensitive reaction on Nicotiana occidentalis. Mol. Plant Pathol. 2018, 19, 731–743. [Google Scholar] [CrossRef]
- Osterbaan, L.J.; Choi, J.; Kenney, J.; Flasco, M.; Vigne, E.; Schmitt-Keichinger, C.; Rebelo, A.R.; Heck, M.; Fuchs, M. The identity of a single residue of the RNA-dependent RNA polymerase of Grapevine fanleaf virus modulates vein clearing in Nicotiana benthamiana. Mol. Plant Microbe Interact. 2019, 32, 790–801. [Google Scholar] [CrossRef]
- Hily, J.M.; Poulicard, N.; Kubina, J.; Reynard, J.S.; Spilmont, A.S.; Fuchs, M.; Lemaire, O.; Vigne, E. Metagenomic analysis of nepoviruses: Diversity, evolution and identification of a genome region in members of subgroup A that appears to be important for host range. Arch. Virol. 2021, 166, 2789–2801. [Google Scholar] [CrossRef] [PubMed]
- Vigne, E.; Bergdoll, M.; Guyader, S.; Fuchs, M. Population structure and genetic variability within isolates of Grapevine fanleaf virus from a naturally infected vineyard in France: Evidence for mixed infection and recombination. J. Gen. Virol. 2004, 85, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Vigne, E.; Garcia, S.; Komar, V.; Lemaire, O.; Hily, J.-M. Comparison of serological and molecular methods with high-throughput sequencing for the detection and quantification of Grapevine fanleaf virus in vineyard samples. Front. Microbiol. 2018, 9, 2726. [Google Scholar] [CrossRef] [PubMed]
- Garcia, S.; Hily, J.-M.; Komar, V.; Gertz, C.; Demangeat, G.; Lemaire, O.; Vigne, E. Detection of multiple variants of Grapevine fanleaf virus in single Xiphinema index nematodes. Viruses 2019, 11, 1139. [Google Scholar] [CrossRef] [PubMed]
- Cepin, U.; Gutiérrez-Aguirre, I.; Ravnikar, M.; Pompe-Novak, M. Frequency of occurrence and genetic variability of Grapevine fanleaf virus satellite RNA. Plant Pathol. 2016, 65, 510–520. [Google Scholar] [CrossRef]
- Gottula, J.; Lapato, D.; Cantilina, K.; Saito, S.; Bartlett, B.; Fuchs, M. Genetic variability, evolution, and biological effects of Grapevine fanleaf virus satellite RNAs. Phytopathology 2013, 103, 1180–1187. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, M.; Lemaire, O. Novel approaches for viral disease management. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 599–621. [Google Scholar]
- Demangeat, G.; Voisin, R.; Minot, J.C.; Bosselut, N.; Fuchs, M.; Esmenjaud, D. Survival of Xiphinema index in vineyard soil and retention of Grapevine fanleaf virus over extended time in the absence of host plants. Phytopathology 2005, 95, 1151–1156. [Google Scholar] [CrossRef]
- Walker, M.A.; Wolpert, J.A.; Weber, E. Viticultural characteristics of VR hybrid rootstocks in a vineyard site infected with Grapevine fanleaf virus. Vitis 1994, 33, 19–23. [Google Scholar]
- Oliver, J.E.; Vigne, E.; Fuchs, M. Genetic structure and molecular variability of Grapevine fanleaf virus populations. Virus Res. 2010, 152, 30–40. [Google Scholar] [CrossRef]
- Ollat, N.; Claverie, M.; Esmenjaud, D.; Demangeat, G.; Jacquet, O.; Lemaire, O.; Van Helden, M.; Bloy, P.; Audeguin, L. Un porte-greffe pour lutter contre le court-noué: Le Nemadex Alain Bouquet, du nom de son créateur, retarde la contamination des vignes. Phytoma 2011, 649, 29–33. [Google Scholar]
- European Commission. Council Decision of 18 March 2003 concerning the non-inclusion of aldicarb in Annex I to Council Directive 91/414/EEC and the withdrawal of authorisations for plant protection products containing this active substance (2003/199/EC). Off. J. Eur. Union 2003, 76, 21–23. [Google Scholar]
- European Commission. Council Decision of 20 September 2007 concerning the no inclusion of 1,3-dichloropropene in Annex I to Council Directive 91/414/EEC and the withdrawal of authorisations for plant protection products containing that substance (notified under document number C(2007) 4281). Off. J. Eur. Union 2007, 249, 11–13. [Google Scholar]
- Villate, L.; Morin, E.; Demangeat, G.; Van Helden, M.; Esmenjaud, D. Control of Xiphinema index populations by fallow plants under greenhouse and field conditions. Phytopathology 2012, 102, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Negrel, L.; Baltenweck, R.; Demangeat, G.; Le Bohec-Dorner, F.; Rustenholz, C.; Velt, A.; Gertz, C.; Bieler, E.; Durrenberger, M.; Gombault, P.; et al. Comparative metabolomic analysis of four Fabaceae and relationship to in vitro nematicidal activity against Xiphinema index. Molecules 2022, 27, 3052. [Google Scholar] [CrossRef] [PubMed]
- Hemmer, C.; Djennane, S.; Ackerer, L.; Hleibieh, K.; Marmonier, A.; Gersch, S.; Garcia, S.; Vigne, E.; Komar, V.; Perrin, M.; et al. Nanobody-mediated resistance to Grapevine fanleaf virus in plants. Plant Biotechnol. J. 2018, 16, 660–671. [Google Scholar] [CrossRef]
- Djennane, S.; Prado, E.; Dumas, V.; Demangeat, G.; Gersch, S.; Alais, A.; Gertz, C.; Beuve, M.; Lemaire, O.; Merdinoglu, D. A single resistance factor to solve vineyard degeneration due to Grapevine fanleaf virus. Commun. Biol. 2021, 4, 637. [Google Scholar] [CrossRef]
- Pechinger, K.; Chooi, K.M.; MacDiarmid, R.M.; Harper, S.J.; Ziebell, H. A new era for mild strain cross-protection. Viruses 2019, 11, 670. [Google Scholar] [CrossRef]
- McKinney, H.H. Mosaic diseases in the Canary Islands, West Africa and Gibraltar. J. Agric. Res. 1929, 39, 577–578. [Google Scholar]
- Ziebell, H.; Carr, J.P. Chapter 6-Cross-protection: A century of mystery. In Advances in Virus Research; Carr, J.P., Loebenstein, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 76, pp. 211–264. [Google Scholar]
- Ziebell, H.; MacDiarmid, R. Prospects for engineering and improvement of cross-protective virus strains. Curr. Opin. Virol. 2017, 26, 8–14. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Zhang, S.; Guo, Q.; Sun, R.; Wei, T.; Qu, F. A new mechanistic model for viral cross protection and superinfection exclusion. Front. Plant Sci. 2018, 9, 40. [Google Scholar] [CrossRef]
- Chewachong, G.M.; Miller, S.A.; Blakeslee, J.J.; Francis, D.M.; Morris, T.J.; Qu, F. Generation of an attenuated, cross-protective Pepino mosaic virus variant through alignment-guided mutagenesis of the viral capsid protein. Phytopathology 2015, 105, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Agüero, J.; Gómez-Aix, C.; Sempere, R.N.; García-Villalba, J.; García-Núñez, J.; Hernando, Y.; Aranda, M.A. Stable and broad spectrum cross-protection against Pepino mosaic virus attained by mixed infection. Front. Plant Sci. 2018, 9, 1810. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.S.; Müller, G.W. Tristeza control by cross protection: A U.S.-Brazil cooperative success. Plant Dis. 1980, 64, 538–541. [Google Scholar] [CrossRef]
- Moreno, P.; Ambros, S.; Albiach-Marti, M.R.; Guerri, J.; Pena, L. Citrus tristeza virus: A pathogen that changed the course of the citrus industry. Mol. Plant Pathol. 2008, 9, 251–268. [Google Scholar] [CrossRef]
- Lecoq, H.; Raccah, B. Cross-protection: Interactions between strains exploited to control plant virus diseases. In Biotic Interactions in Plant–Pathogen Associations; Jeger, M.J., Spence, N.J., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 177–192. [Google Scholar] [CrossRef]
- Muller, G.W.; Rezende, J.A.M. Preimmunization: Applications and perspectives in virus disease control. In Diseases of Fruits and Vegetables Volume I: Diagnosis and Management; Naqvi, S.A.M.H., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 361–395. [Google Scholar]
- Cong, Q.Q.; Wang, Y.; Liu, J.; Lan, Y.F.; Guo, Z.K.; Yang, J.G.; Li, X.D.; Tian, Y.P. Evaluation of Potato virus X mild mutants for cross protection against severe infection in China. Virol. J. 2019, 16, 36. [Google Scholar] [CrossRef]
- Huang, X.-D.; Fang, L.; Gu, Q.-S.; Tian, Y.-P.; Geng, C.; Li, X.-D. Cross protection against the watermelon strain of Papaya ringspot virus through modification of viral RNA silencing suppressor. Virus Res. 2019, 265, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, X.D.; Xu, S. Single amino acid substitutions in the coat protein and RNA-dependent RNA polymerase alleviated the virulence of Cucumber green mottle mosaic virus and conferred cross protection against severe infection. Virus Genes 2020, 56, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.-T.-Y.; Lin, T.-T.; Chang, C.-P.; Chen, C.-H.; Nguyen, V.-H.; Yeh, S.-D. Generation of mild recombinants of Papaya ringspot virus to minimize the problem of strain-specific cross protection. Phytopathology 2022, 112, 708–719. [Google Scholar] [CrossRef]
- Salibe, A.A.; Souza, A.A.; Targon, M.L.P.N.; Müller, G.W.; Filho, H.D.C.; Machado, M.A. Selection of a mild sub-isolate of Citrus tristeza virus for preimmunization of Pera sweet orange. In International Organization of Citrus Virologists Conference Proceedings (1957–2010); International Organization of Citrus Virologists: Riverside, CA, USA, 2002. [Google Scholar]
- Baba, V.Y.; Giampani, J.S.; Tazima, Z.H.; Yada, I.F.U.; Paccola-Meirelles, L.D.; Leite, R.P. Agronomic performance of Pera and related sweet orange accessions naturally infected with Citrus tristeza virus in northern Paraná state, Brazil. Trop. Plant Pathol. 2014, 39, 442–448. [Google Scholar] [CrossRef]
- Huss, B.; Walter, B.; Fuchs, M. Cross-protection between Arabis mosaic virus and Grapevine fanleaf virus isolates in Chenopodium quinoa. Ann. Appl. Biol. 1989, 114, 45–60. [Google Scholar] [CrossRef]
- Komar, V.; Vigne, E.; Demangeat, G.; Lemaire, O.; Fuchs, M. Cross-protection as control strategy against Grapevine fanleaf virus in naturally infected vineyards. Plant Dis. 2008, 92, 1689–1694. [Google Scholar] [CrossRef] [PubMed]
- PNDV. Résultats de L’enquête Court-Noue Auprès des Viticulteurs Champenois. Available online: https://www.plan-deperissement-vigne.fr/webzine/parole-dexpert/resultats-de-lenquete-court-noue-aupres-des-viticulteurs-champenois (accessed on 2 December 2021).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Hily, J.M.; Demaneche, S.; Poulicard, N.; Tannieres, M.; Djennane, S.; Beuve, M.; Vigne, E.; Demangeat, G.; Komar, V.; Gertz, C.; et al. Metagenomic-based impact study of transgenic grapevine rootstock on its associated virome and soil bacteriome. Plant Biotechnol. J. 2018, 16, 208–220. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2007, 1, 47–50. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Khoosal, A.; Muhire, B. Detecting and analyzing genetic recombination using RDP4. In Bioinformatics: Volume I: Data, Sequence Analysis, and Evolution; Keith, J.M., Ed.; Springer: New York, NY, USA, 2017; pp. 433–460. [Google Scholar]
- Esmenjaud, D.; Walter, B.; Valentin, G.; Guo, Z.T.; Cluzeau, D.J.C. Vertical distribution and infectious potential of Xiphinema index (Thorne et Allen, 1950) (Nematoda: Longidoridae) in fields affected by grapevine fanleaf virus in vineyards in the Champagne region of France. Agronomie 1992, 12, 395–399. [Google Scholar] [CrossRef]
- Villate, L.; Fievet, V.; Hanse, B.; Delemarre, F.; Plantard, O.; Esmenjaud, D.; van Helden, M. Spatial distribution of the dagger nematode Xiphinema index and its associated Grapevine fanleaf virus in French vineyard. Phytopathology 2008, 98, 942–948. [Google Scholar] [CrossRef]
- Perring, T.M.; Farrar, C.A.; Blua, M.J.; Wang, H.L.; Gonsalves, D. Cross protection of cantaloupe with a mild strain of Zucchini yellow mosaic virus: Effectiveness and application. Crop Prot. 1995, 14, 601–606. [Google Scholar] [CrossRef]
- CIVC. La Plantation. Available online: https://www.champagne.fr/fr/vigne-vin/cepages-vignes/plantation (accessed on 29 September 2021).
- CIVC. Chaque Vendange est Unique. Available online: https://www.champagne.fr/fr/vigne-vin/travail-vigne/les-vendanges-en-champagne (accessed on 29 September 2021).
- Syller, J. Facilitative and antagonistic interactions between plant viruses in mixed infections. Mol. Plant Pathol. 2012, 13, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.B.; López-Moya, J.J. When viruses play team sports: Mixed infections in plants. Phytopathology 2020, 110, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, B.; Freeborough, M.J.; Maree, H.J.; Celton, J.M.; Rees, D.J.G.; Burger, J.T. Deep sequencing analysis of viruses infecting grapevines: Virome of a vineyard. Virology 2010, 400, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Eichmeier, A.; Komínková, M.; Komínek, P.; Baránek, M. Comprehensive virus detection using next generation sequencing in grapevine vascular tissues of plants obtained from the wine regions of Bohemia and Moravia (Czech Republic). PLoS ONE 2016, 11, e0167966. [Google Scholar] [CrossRef]
- Fajardo, T.V.M.; Silva, F.N.; Eiras, M.; Nickel, O. High-throughput sequencing applied for the identification of viruses infecting grapevines in Brazil and genetic variability analysis. Trop. Plant Pathol. 2017, 42, 250–260. [Google Scholar] [CrossRef]
- Beuve, M.; Hily, J.M.; Alliaume, A.; Reinbold, C.; Le Maguet, J.; Candresse, T.; Herrbach, E.; Lemaire, O. A complex virome unveiled by deep sequencing analysis of RNAs from a French Pinot Noir grapevine exhibiting strong leafroll symptoms. Arch. Virol. 2018, 163, 2937–2946. [Google Scholar] [CrossRef]
- Czotter, N.; Molnar, J.; Szabó, E.; Demian, E.; Kontra, L.; Baksa, I.; Szittya, G.; Kocsis, L.; Deak, T.; Bisztray, G.; et al. NGS of virus-derived small RNAs as a diagnostic method used to determine viromes of Hungarian vineyards. Front. Microbiol. 2018, 9, 122. [Google Scholar] [CrossRef]
- Xiao, H.; Li, C.; Rwahnih, M.A.; Dolja, V.; Meng, B. Metagenomic analysis of Riesling grapevine reveals a complex virome including two new and divergent variants of Grapevine leafroll-associated virus 3. Plant Dis. 2019, 103, 1275–1285. [Google Scholar] [CrossRef]
- Sidharthan, V.K.; Sevanthi, A.M.; Jaiswal, S.; Baranwal, V.K. Robust virome profiling and whole genome reconstruction of viruses and viroids enabled by use of available mRNA and sRNA-Seq datasets in grapevine (Vitis vinifera L.). Front. Microbiol. 2020, 11, 1232. [Google Scholar] [CrossRef]
- Velasco, L.; Padilla, C.V. High-throughput sequencing of small RNAs for the sanitary certification of viruses in grapevine. Front. Plant Sci. 2021, 12, 682879. [Google Scholar] [CrossRef]
- Saldarelli, P.; Giampetruzzi, A.; Maree, H.J.; Rwahnih, M.A. High-Throughput Sequencing: Advantages beyond virus identification. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 625–642. [Google Scholar]
- Angelini, E.; Aboughanem-Sabanadzovic, N.; Dolja, V.V.; Meng, B. Grapevine leafroll-associated virus 2. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 141–165. [Google Scholar]
- Tarquini, G.; Zaina, G.; Ermacora, P.; De Amicis, F.; Franco-Orozco, B.; Loi, N.; Martini, M.; Bianchi, G.L.; Pagliari, L.; Firrao, G.; et al. Agroinoculation of grapevine Pinot gris virus in tobacco and grapevine provides insights on viral pathogenesis. PLoS ONE 2019, 14, e0214010. [Google Scholar] [CrossRef] [PubMed]
- Maclot, F.; Candresse, T.; Filloux, D.; Malmstrom, C.M.; Roumagnac, P.; van der Vlugt, R.; Massart, S. Illuminating an ecological blackbox: Using high throughput sequencing to characterize the plant virome across scales. Front. Microbiol. 2020, 11, 578064. [Google Scholar] [CrossRef] [PubMed]
- Gaspero, G.D.; Radovic, S.; De Luca, E.; Spadotto, A.; Magris, G.; Falginella, L.; Cattonaro, F.; Marroni, F. Evaluation of sensitivity and specificity in RNA-Seq-based detection of grapevine viral pathogens. J. Virol. Methods 2021, 300, 114383. [Google Scholar] [CrossRef] [PubMed]
- Osterbaan, L.J.; Fuchs, M. Dynamic interactions between plant viruses and their hosts for symptom development. J. Plant Pathol. 2019, 101, 885–895. [Google Scholar] [CrossRef]
- Gu, H.; Zhang, C.; Ghabrial, S.A. Novel naturally occurring Bean pod mottle virus reassortants with mixed heterologous RNA1 genomes. Phytopathology 2007, 97, 79–86. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Cooper, J.I.; Edwards, M.L.; Hellen, C.U.T. A satellite RNA of arabis mosaic nepovirus and its pathological impact. Ann. Appl. Biol. 1991, 118, 577–587. [Google Scholar] [CrossRef]
- Shimura, H.; Pantaleo, V.; Ishihara, T.; Myojo, N.; Inaba, J.I.; Sueda, K.; Burgyan, J.; Masuta, C. A viral satellite RNA induces yellow symptoms on tobacco by targeting a gene involved in chlorophyll biosynthesis using the RNA silencing machinery. PLoS Pathog. 2011, 7, e1002021. [Google Scholar] [CrossRef]
- Lamprecht, R.L.; Spaltman, M.; Stephan, D.; Wetzel, T.; Burger, J.T. Complete nucleotide sequence of a South African isolate of Grapevine fanleaf virus and its associated satellite RNA. Viruses 2013, 5, 1815–1823. [Google Scholar] [CrossRef]
- Bashir, N.S.; Zarghani, S.N.; Hejazi, M.S. Diversity of Grapevine fanleaf virus isolates from Iran. Virus Res. 2007, 128, 144–148. [Google Scholar] [CrossRef]
- Pompe-Novak, M.; Gutiérrez-Aguirre, I.; Vojvoda, J.; Blas, M.; Tomažič, I.; Vigne, E.; Fuchs, M.; Ravnikar, M.; Petrovič, N. Genetic variability within RNA2 of Grapevine fanleaf virus. Eur. J. Plant Pathol. 2007, 117, 307–312. [Google Scholar] [CrossRef]
- Vigne, E.; Marmonier, A.; Komar, V.; Lemaire, O.; Fuchs, M. Genetic structure and variability of virus populations in cross-protected grapevines superinfected by Grapevine fanleaf virus. Virus Res. 2009, 144, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Vigne, E.; Demangeat, G.; Komar, V.; Fuchs, M. Characterization of a naturally occuring recombinant isolate of Grapevine fanleaf virus. Arch. Virol. 2005, 150, 2241–2255. [Google Scholar] [CrossRef] [PubMed]
- Mekuria, T.A.; Gutha, L.R.; Martin, R.R.; Naidu, R.A. Genome diversity and intra- and interspecies recombination events in grapevine fanleaf virus. Phytopathology 2009, 99, 1394–1402. [Google Scholar] [CrossRef]
- Folimonova, S.Y.; Robertson, C.J.; Shilts, T.; Folimonov, A.S.; Hilf, M.E.; Garnsey, S.M.; Dawson, W.O. Infection with strains of Citrus tristeza virus does not exclude superinfection by other strains of the virus. J. Virol. 2010, 84, 1314–1325. [Google Scholar] [CrossRef] [PubMed]
- Folimonova, S.Y. Developing an understanding of cross-protection by Citrus tristeza virus. Front. Microbiol. 2013, 4, 76. [Google Scholar] [CrossRef]
- Desbiez, C.; Gal-On, A.; Raccah, B.; Lecoq, H. Characterization of epitopes on Zucchini yellow mosaic Potyvirus coat protein permits studies on the interactions between strains. J. Gen. Virol. 1997, 78, 2073–2076. [Google Scholar] [CrossRef][Green Version]
- Medina-Salguero, A.X.; Cornejo-Franco, J.F.; Grinstead, S.; Mowery, J.; Mollov, D.; Quito-Avila, D.F. Genetic characterization of a mild isolate of Papaya ringspot virus type-P (PRSV-P) and assessment of its cross-protection potential under greenhouse and field conditions. PLoS ONE 2021, 16, e0241652. [Google Scholar] [CrossRef]
- Scott, K.A.; Hlela, Q.; Zablocki, O.; Read, D.; van Vuuren, S.; Pietersen, G. Genotype composition of populations of grapefruit-cross-protecting citrus tristeza virus strain GFMS12 in different host plants and aphid-transmitted sub-isolates. Arch. Virol. 2013, 158, 27–37. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M− | M+ | S+ | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pa3 | Pa5 | Pa7 | Pa9 | Py11 | Py13 | Py16 | Pa1 | Py18 | Py19 | Py43 | Pa2 | Pa4 | Pa6 | Pa8 | Pa10 | Py12 | Py14 | Py15 | Py17 | Py20 | |
| 2016 | − | − | − | − | − | − | − | + | + | + | nd | + | + | + | + | + | + | + | + | + | + |
| 2017 | − | − | − | − | − | − | − | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 2018 | − | − | − | − | − | − | − | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 2019 | − | − | − | − | − | − | − | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| M+ | S+ | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pa1 | Py18 | Py19 | Py43 | Pa2 | Pa4 | Pa6 | Pa8 | Pa10 | Py12 | Py14 | Py15 | Py17 | Py20 | |
| I.A | 1-1 | 1-1/1-2 | 1-1/1-2 | 1-1 | 1-1 | |||||||||
| I.B | 1-1 | 1-1 | 1-2 | 1-3 | 1-1 | |||||||||
| I.C | 1-1 | 1-1 | 1-1 | |||||||||||
| I.D | 1-3 | 1-2 | ||||||||||||
| I.E | 1-1 | 1-1 | ||||||||||||
| I.F | 1-2/1-3 | |||||||||||||
| I.G | 1-2 | |||||||||||||
| I.H | 1-2 | |||||||||||||
| I.I | 1-2 | |||||||||||||
| I.J | 1-2 | |||||||||||||
| I.K | 1-2 | |||||||||||||
| I.L | 1-1 | |||||||||||||
| II.A | 2-1 | 2-1 | 2-2 | 2-2 | 2-1 | |||||||||
| II.B | 2-1 | 2-1 | 2-2 | 2-2 | 2-2 | |||||||||
| II.C | 2-1 | 2-1 | 2-1 | 2-1 | 2-1 | |||||||||
| II.D | 2-1 | 2-1 | ||||||||||||
| II.E | 2-1 | |||||||||||||
| II.F | 2-1 | |||||||||||||
| III.A | 3-1/3-2 | 3-1 | 3-1 | |||||||||||
| III.B | 3-1 | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubina, J.; Hily, J.-M.; Mustin, P.; Komar, V.; Garcia, S.; Martin, I.R.; Poulicard, N.; Velt, A.; Bonnet, V.; Mercier, L.; et al. Characterization of Grapevine Fanleaf Virus Isolates in ‘Chardonnay’ Vines Exhibiting Severe and Mild Symptoms in Two Vineyards. Viruses 2022, 14, 2303. https://doi.org/10.3390/v14102303
Kubina J, Hily J-M, Mustin P, Komar V, Garcia S, Martin IR, Poulicard N, Velt A, Bonnet V, Mercier L, et al. Characterization of Grapevine Fanleaf Virus Isolates in ‘Chardonnay’ Vines Exhibiting Severe and Mild Symptoms in Two Vineyards. Viruses. 2022; 14(10):2303. https://doi.org/10.3390/v14102303
Chicago/Turabian StyleKubina, Julie, Jean-Michel Hily, Pierre Mustin, Véronique Komar, Shahinez Garcia, Isabelle Rachel Martin, Nils Poulicard, Amandine Velt, Véronique Bonnet, Laurence Mercier, and et al. 2022. "Characterization of Grapevine Fanleaf Virus Isolates in ‘Chardonnay’ Vines Exhibiting Severe and Mild Symptoms in Two Vineyards" Viruses 14, no. 10: 2303. https://doi.org/10.3390/v14102303
APA StyleKubina, J., Hily, J.-M., Mustin, P., Komar, V., Garcia, S., Martin, I. R., Poulicard, N., Velt, A., Bonnet, V., Mercier, L., Lemaire, O., & Vigne, E. (2022). Characterization of Grapevine Fanleaf Virus Isolates in ‘Chardonnay’ Vines Exhibiting Severe and Mild Symptoms in Two Vineyards. Viruses, 14(10), 2303. https://doi.org/10.3390/v14102303