
Effectiveness of Live-Attenuated Genotype III Japanese Encephalitis Viral Vaccine against Circulating Genotype I Viruses in Swine

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
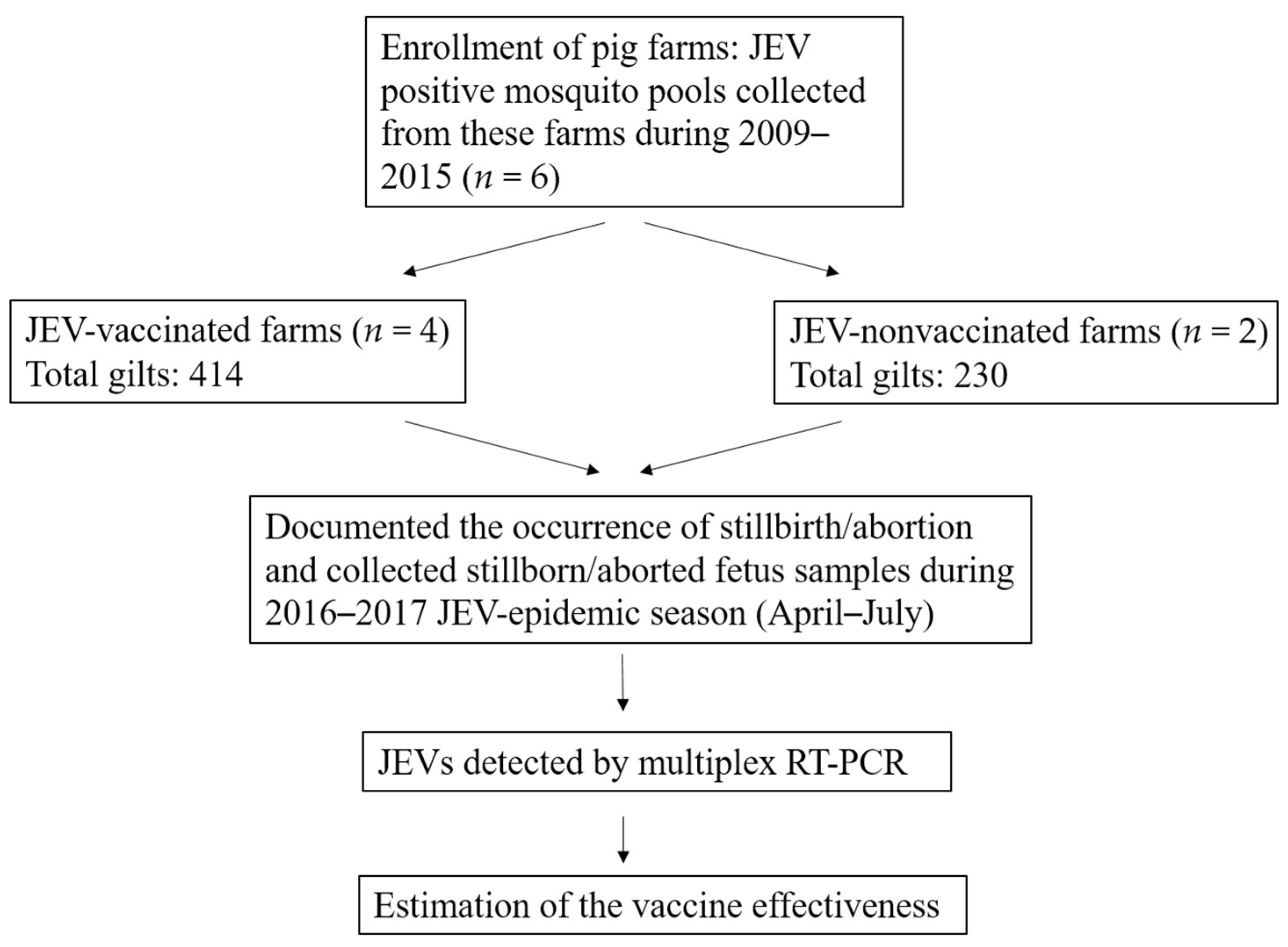
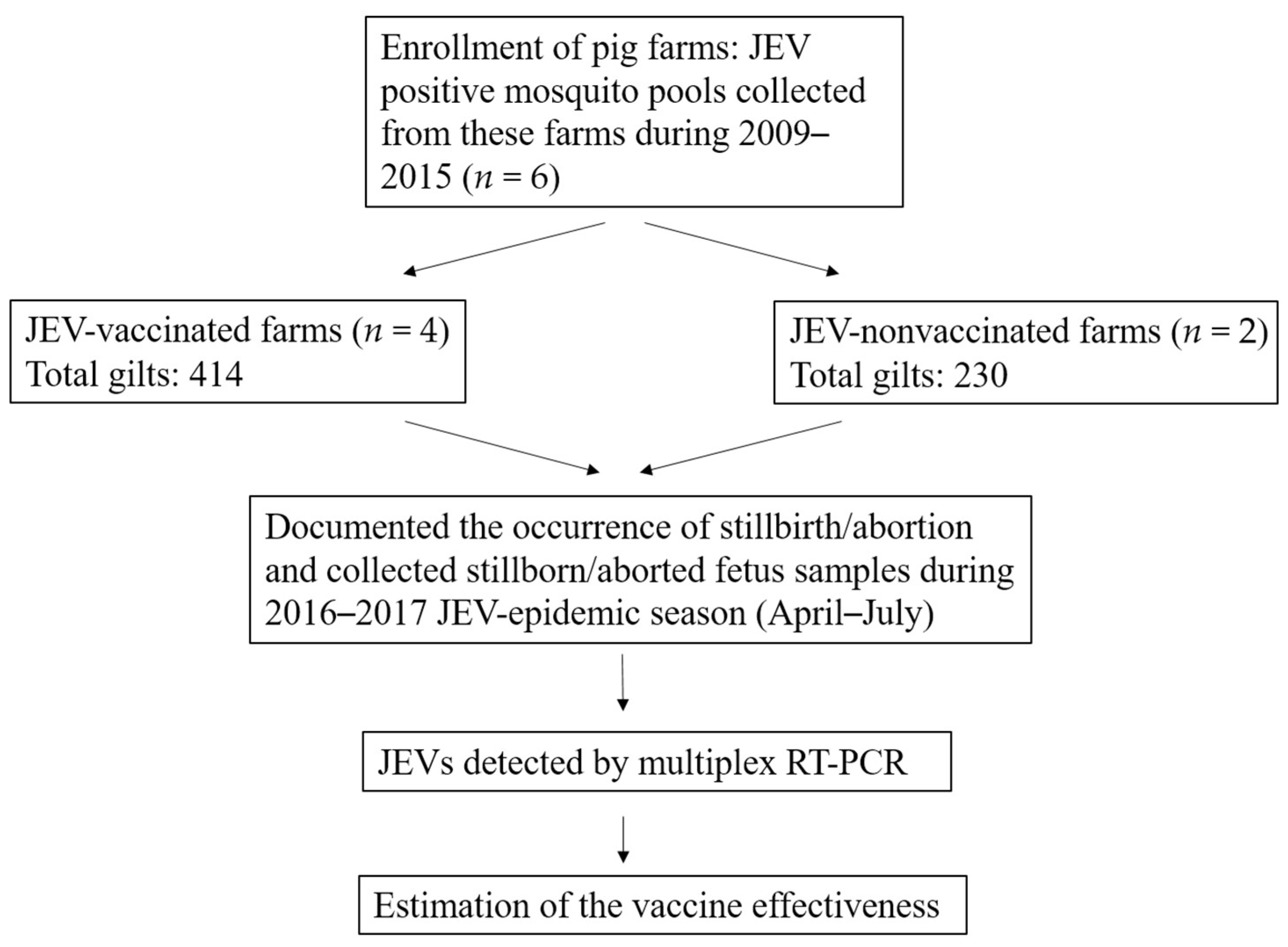
2.1. Study Design
2.2. Sample Collection
2.3. Neutralizing Antibody
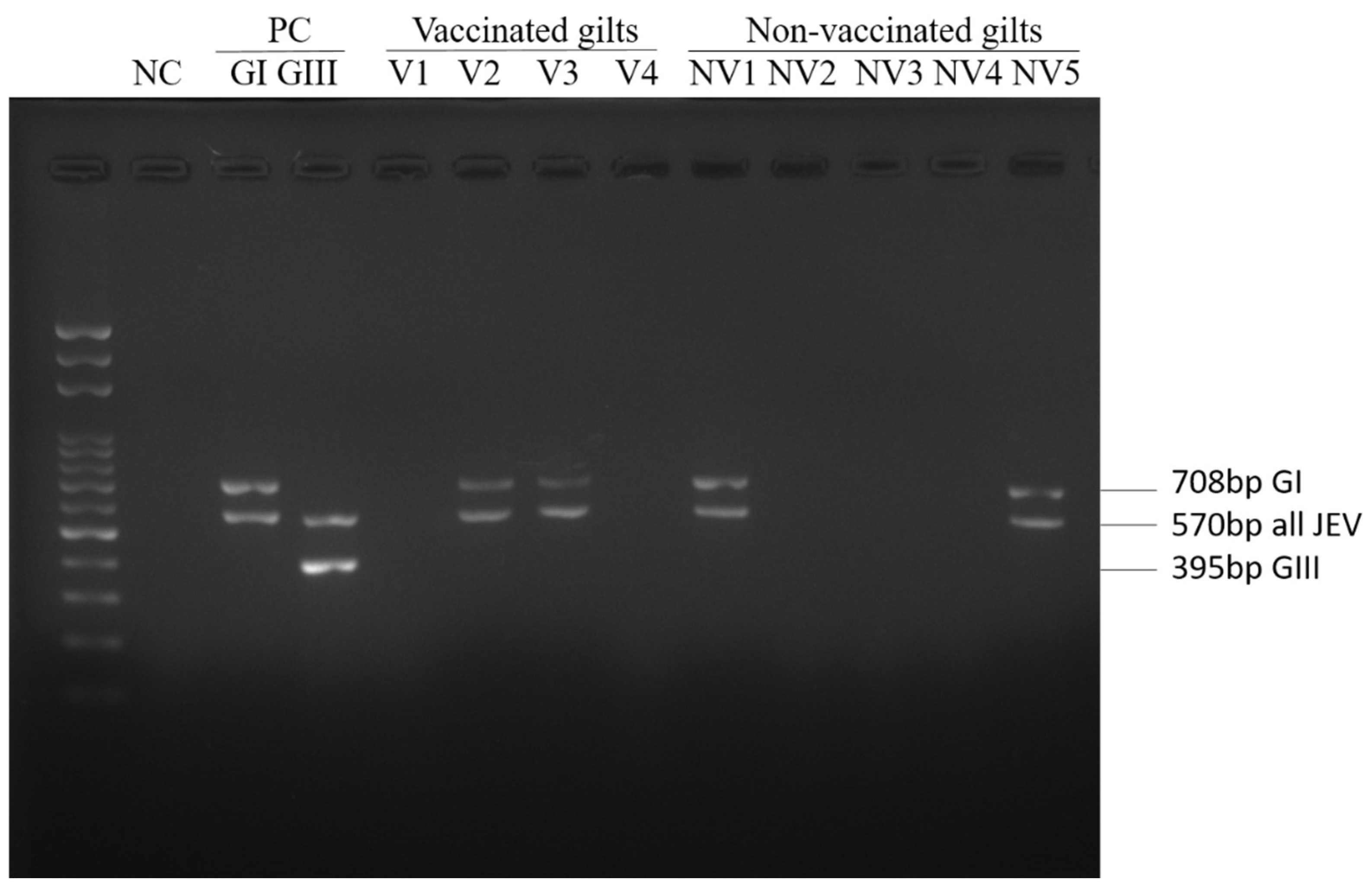
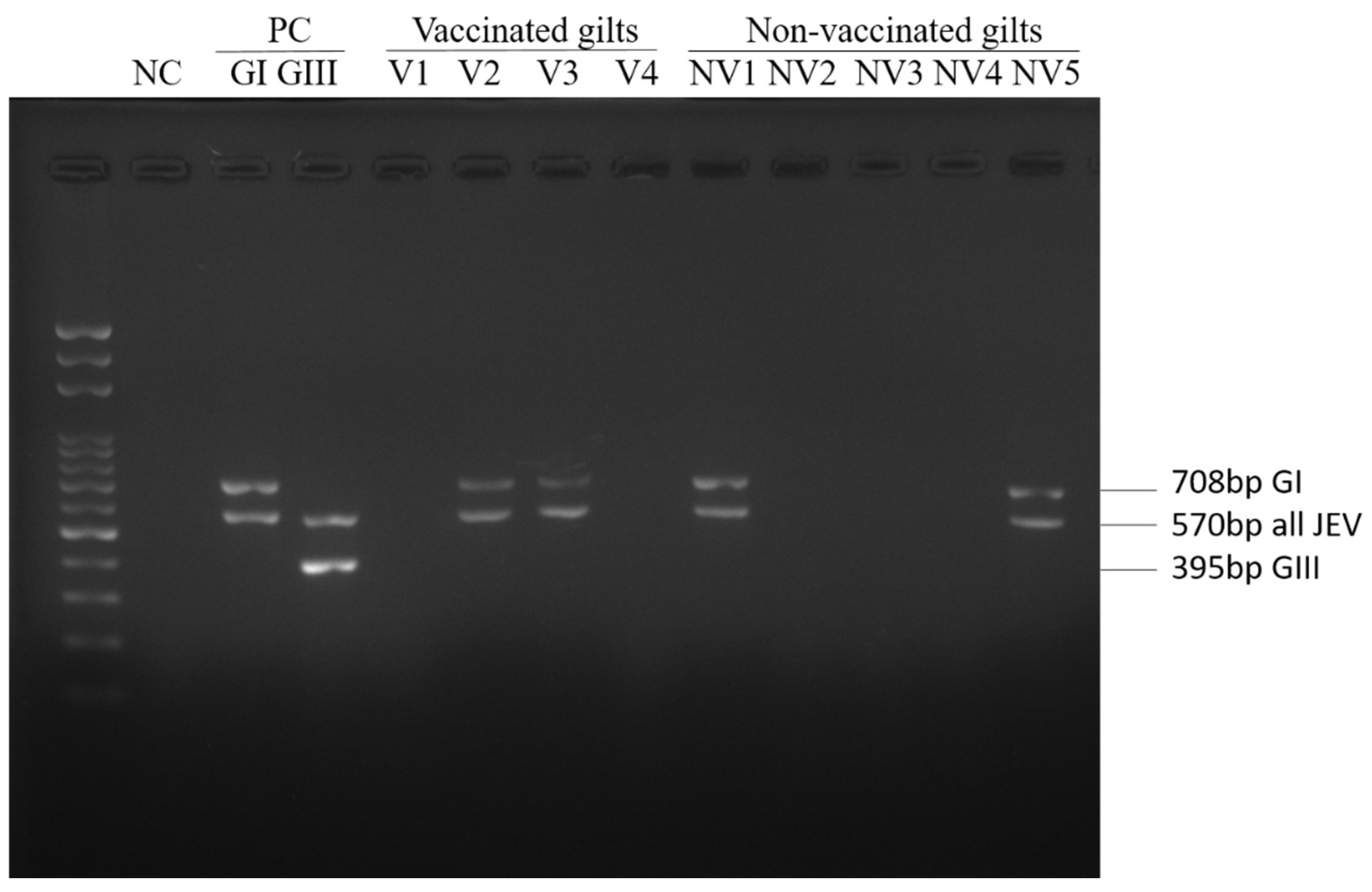
2.4. Multiplex RT-PCR
2.5. Phylogenetic Analysis
2.6. Statistics
3. Results
3.1. Characteristics of Enrolled Pig Farms
3.2. Occurrence of Stillbirth/Abortion
3.3. Estimates of Vaccine Effectiveness
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mansfield, K.L.; Hernandez-Triana, L.M.; Banyard, A.C.; Fooks, A.R.; Johnson, N. Japanese encephalitis virus infection, diagnosis and control in domestic animals. Vet. Microbiol. 2017, 201, 85–92. [Google Scholar] [CrossRef]
- Unni, S.K.; Ruzek, D.; Chhatbar, C.; Mishra, R.; Johri, M.K.; Singh, S.K. Japanese encephalitis virus: From genome to infectome. Microbes Infect. 2011, 13, 312–321. [Google Scholar] [CrossRef]
- Ladreyt, H.; Durand, B.; Dussart, P.; Chevalier, V. How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control. Viruses 2019, 11, 949. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.I.; Lee, Y.M. Japanese encephalitis: The virus and vaccines. Hum. Vaccin Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannice, K.S.; Hills, S.L.; Schwartz, L.M.; Barrett, A.D.; Heffelfinger, J.; Hombach, J.; Letson, G.W.; Solomon, T.; Marfin, A.A.; Anderson, K.; et al. The future of Japanese encephalitis vaccination: Expert recommendations for achieving and maintaining optimal JE control. NPJ Vaccines 2021, 6, 82. [Google Scholar] [CrossRef]
- Fan, Y.C.; Chen, J.M.; Chen, Y.Y.; Lin, J.W.; Chiou, S.S. Reduced neutralizing antibody titer against genotype I virus in swine immunized with a live-attenuated genotype III Japanese encephalitis virus vaccine. Vet. Microbiol. 2013, 163, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.C.; Huang, Y.S.; Chien, L.J.; Lin, T.L.; Yueh, Y.Y.; Tseng, W.L.; Chang, K.J.; Wang, G.R. The epidemiology of Japanese encephalitis on Taiwan during 1966–1997. Am. J. Trop. Med. Hyg. 1999, 61, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus genome organization, expression, and replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.; Ni, H.; Beasley, D.W.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D. Origin and evolution of Japanese encephalitis virus in southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef] [Green Version]
- Schuh, A.J.; Ward, M.J.; Leigh Brown, A.J.; Barrett, A.D. Dynamics of the emergence and establishment of a newly dominant genotype of Japanese encephalitis virus throughout Asia. J. Virol. 2014, 88, 4522–4532. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.C.; Liang, J.J.; Chen, J.M.; Lin, J.W.; Chen, Y.Y.; Su, K.H.; Lin, C.C.; Tu, W.C.; Chiou, M.T.; Ou, S.C.; et al. NS2B/NS3 mutations enhance the infectivity of genotype I Japanese encephalitis virus in amplifying hosts. PLoS Pathog. 2019, 15, e1007992. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Di, D.; Huang, H.; Wang, X.; Xia, Q.; Ma, X.; Liu, K.; Li, B.; Shao, D.; Qiu, Y.; et al. NS5-V372A and NS5-H386Y variations are responsible for differences in interferon alpha/beta induction and co-contribute to the replication advantage of Japanese encephalitis virus genotype I over genotype III in ducklings. PLoS Pathog. 2020, 16, e1008773. [Google Scholar] [CrossRef] [PubMed]
- Erra, E.O.; Askling, H.H.; Yoksan, S.; Rombo, L.; Riutta, J.; Vene, S.; Lindquist, L.; Vapalahti, O.; Kantele, A. Cross-protective capacity of Japanese encephalitis (JE) vaccines against circulating heterologous JE virus genotypes. Clin. Infect. Dis. 2013, 56, 267–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.C.; Chen, J.M.; Chiu, H.C.; Chen, Y.Y.; Lin, J.W.; Shih, C.C.; Chen, C.M.; Chang, C.C.; Chang, G.J.; Chiou, S.S. Partially neutralizing potency against emerging genotype I virus among children received formalin-inactivated Japanese encephalitis virus vaccine. PLoS Negl. Trop. Dis. 2012, 6, e1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, S.T.; Chang, L.C.; Lin, S.Y.; Chuang, T.Y.; Ma, C.H.; Inoue, Y.K.; Okuno, T. The effect of vaccination with a live attenuated strain of Japanese encephalitis virus on stillbirths in swine in Taiwan. Bull. World Health Organ. 1972, 46, 465–471. [Google Scholar]
- Yang, S.E.; Pan, M.J.; Tseng, H.F.; Liau, M.Y. The efficacy of mouse-brain inactivated Nakayama strain Japanese encephalitis vaccine—Results from 30 years experience in Taiwan. Vaccine 2006, 24, 2669–2673. [Google Scholar] [CrossRef]
- Shimizu, T.; Kawakami, Y.; Fukuhara, S.; Matumoto, M. Experimental stillbirth in pregnant swine infected with Japanese encephalitis virus. Jpn J. Exp. Med. 1954, 24, 363–375. [Google Scholar]
- Takashima, I.; Watanabe, T.; Ouchi, N.; Hashimoto, N. Ecological studies of Japanese encephalitis virus in Hokkaido: Interepidemic outbreaks of swine abortion and evidence for the virus to overwinter locally. Am. J. Trop. Med. Hyg. 1988, 38, 420–427. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Fan, Y.C.; Tu, W.C.; Chang, R.Y.; Shih, C.C.; Lu, I.H.; Chien, M.S.; Lee, W.C.; Chen, T.H.; Chang, G.J.; et al. Japanese encephalitis virus genotype replacement, Taiwan, 2009–2010. Emerg. Infect. Dis. 2011, 17, 2354–2356. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Lin, J.W.; Fan, Y.C.; Tu, W.C.; Chang, G.J.; Chiou, S.S. First detection of the Africa/Caribbean/Latin American subtype of Culex flavivirus in Asian country, Taiwan. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 387–396. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Lin, J.W.; Fan, Y.C.; Chiou, S.S. Detection and differentiation of genotype I and III Japanese encephalitis virus in mosquitoes by multiplex reverse transcriptase-polymerase chain reaction. Transbound. Emerg. Dis. 2014, 61, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Su, C.L.; Yang, C.F.; Teng, H.J.; Lu, L.C.; Lin, C.; Tsai, K.H.; Chen, Y.Y.; Chen, L.Y.; Chang, S.F.; Shu, P.Y. Molecular epidemiology of Japanese encephalitis virus in mosquitoes in Taiwan during 2005–2012. PLoS Negl. Trop. Dis. 2014, 8, e3122. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Wu, R.; Feng, Y.; Zhao, Q.; Wen, X.; Huang, X.; Wen, Y.; Yan, Q.; Huang, Y.; Ma, X.; et al. Genomic changes in an attenuated genotype I Japanese encephalitis virus and comparison with virulent parental strain. Virus Genes 2018, 54, 424–431. [Google Scholar] [CrossRef]
- Nidaira, M.; Kyan, H.; Taira, K.; Okano, S.; Oshiro, T.; Kato, T.; Kudo, N.; Azama, Y.; Mahoe, Y.; Kudaka, J.; et al. Survey of Japanese encephalitis virus in pigs and wild boars on Ishigaki and Iriomote Islands in Okinawa, Japan. Epidemiol. Infect. 2014, 142, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Hameed, M.; Liu, K.; Anwar, M.N.; Wahaab, A.; Safdar, A.; Di, D.; Boruah, P.; Xu, J.; Wang, X.; Li, B.; et al. The emerged genotype I of Japanese encephalitis virus shows an infectivity similar to genotype III in Culex pipiens mosquitoes from China. PLoS Negl. Trop. Dis. 2019, 13, e0007716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.C.; Lin, J.W.; Liao, S.Y.; Chen, J.M.; Chen, Y.Y.; Chiu, H.C.; Shih, C.C.; Chen, C.M.; Chang, R.Y.; King, C.C.; et al. Virulence of Japanese Encephalitis Virus Genotypes I and III, Taiwan. Emerg. Infect. Dis. 2017, 23, 1883–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beasley, D.W.; Li, L.; Suderman, M.T.; Guirakhoo, F.; Trent, D.W.; Monath, T.P.; Shope, R.E.; Barrett, A.D. Protection against Japanese encephalitis virus strains representing four genotypes by passive transfer of sera raised against ChimeriVax-JE experimental vaccine. Vaccine 2004, 22, 3722–3726. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Wang, X.; Zhang, J.; Guo, S.; Pang, L.; Shi, K.; Liu, K.; Shao, D.; Qiu, Y.; Liu, L.; et al. Partial cross-protection between Japanese encephalitis virus genotype I and III in mice. PLoS Negl. Trop. Dis. 2019, 13, e0007601. [Google Scholar] [CrossRef] [Green Version]
- Feroldi, E.; Boaz, M.; Yoksan, S.; Chokephaibulkit, K.; Thisyakorn, U.; Pancharoen, C.; Monfredo, C.; Bouckenooghe, A. Persistence of Wild-Type Japanese Encephalitis Virus Strains Cross-Neutralization 5 Years After JE-CV Immunization. J. Infect. Dis. 2017, 215, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Bonaparte, M.; Dweik, B.; Feroldi, E.; Meric, C.; Bouckenooghe, A.; Hildreth, S.; Hu, B.; Yoksan, S.; Boaz, M. Immune response to live-attenuated Japanese encephalitis vaccine (JE-CV) neutralizes Japanese encephalitis virus isolates from south-east Asia and India. BMC Infect. Dis. 2014, 14, 156. [Google Scholar] [CrossRef] [Green Version]
- Van Gessel, Y.; Klade, C.S.; Putnak, R.; Formica, A.; Krasaesub, S.; Spruth, M.; Cena, B.; Tungtaeng, A.; Gettayacamin, M.; Dewasthaly, S. Correlation of protection against Japanese encephalitis virus and JE vaccine (IXIARO((R))) induced neutralizing antibody titers. Vaccine 2011, 29, 5925–5931. [Google Scholar] [CrossRef] [PubMed]
- Erra, E.O.; Askling, H.H.; Yoksan, S.; Rombo, L.; Riutta, J.; Vene, S.; Lindquist, L.; Vapalahti, O.; Kantele, A. Cross-protection elicited by primary and booster vaccinations against Japanese encephalitis: A two-year follow-up study. Vaccine 2013, 32, 119–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Zhang, Y.; Zhou, Z.B.; Xia, S.; Shi, W.Q.; Xue, J.B.; Li, Y.Y.; Wu, J.T. New strains of Japanese encephalitis virus circulating in Shanghai, China after a ten-year hiatus in local mosquito surveillance. Parasit Vectors 2019, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Shan, T.; Deng, Y.; Sun, C.; Yuan, S.; Yin, Y.; Tong, G. Molecular characterization of Japanese encephalitis virus strains prevalent in Chinese swine herds. J. Vet. Sci. 2013, 14, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.A.; Borah, J.; Chowdhury, P.; Dutta, P.; Mahanta, J. Characterization of Japanese encephalitis virus (JEV) genotype III clinical isolates in northeast India. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Cherian, S.S.; Walimbe, A.M. Phylogeographic analysis of Japanese encephalitis virus in India (1956–2012). Arch. Virol. 2015, 160, 3097–3104. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.L.; Liu, H.; Wang, H.Y.; Fu, S.H.; Liu, H.Z.; Zhang, H.L.; Li, M.H.; Gao, X.Y.; Wang, J.L.; Sun, X.H.; et al. Emergence of genotype I of Japanese encephalitis virus as the dominant genotype in Asia. J. Virol. 2011, 85, 9847–9853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monath, T.P. Japanese encephalitis vaccines: Current vaccines and future prospects. Curr. Top. Microbiol. Immunol. 2002, 267, 105–138. [Google Scholar]
- Misra, U.K.; Kalita, J. Overview: Japanese encephalitis. Prog. Neurobiol. 2010, 91, 108–120. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Characteristic | Pig Farms | |||||
|---|---|---|---|---|---|---|
| A | B | C | D | E | F | |
| JEV circulating * | Yes | Yes | Yes | Yes | Yes | Yes |
| Location of Taiwan | Central | Central | Southern | Southern | Central | Southern |
| No. of gilts ** | 36 | 50 | 56 | 272 | 54 | 176 |
| Type of farm | farrow-to-finish | farrow-to-finish | farrow-to-finish | matching breed | farrow-to-finish | matching breed |
| JEV Vaccination | Yes | Yes | Yes | Yes | No | No |
| Type of Vaccine | Attenuated | Attenuated | Attenuated | Attenuated | ||
| Time of vaccination | Feb-Mar | Jan-Feb | Feb-Mar | Feb-Mar | ||
| Route of vaccination | subcutaneous | intramuscular | subcutaneous | intramuscular | ||
| Dosage | 2 | 2 | 2 | 2 | ||
| Positivity of Neutralizing Antibody (%) | |||||
|---|---|---|---|---|---|
| Against GIII JEV | Against GI JEV | ||||
| Sow | Gilt (Before Vaccination) | Gilt (After Vaccination) | Sow | Gilt (Before Vaccination) | Gilt (After Vaccination) |
| 80 | 10 | 100 | 80 | 0 | 40 |
| 100 | 0 | 100 | 90 | 0 | 40 |
| 80 | 0 | 100 | 80 | 0 | 30 |
| 90 | 10 | 100 | 90 | 0 | 40 |
| 90 | 0 | 0 | 90 | 0 | 0 |
| 80 | 10 | 0 | 80 | 0 | 0 |
| JEV Vaccination | Vaccination | Non-Vaccination |
|---|---|---|
| Farms | 4 | 2 |
| Gilts | 414 | 230 |
| Litter | 389 | 213 |
| Stillbirth-abortion | 41 | 65 |
| Samples of Stillbirth/abortion | 82 | 130 |
| Positive of JEV multiplex RT-PCR | ||
| GI | 8(6) * | 19(13) ** |
| GIII | 0 | 0 |
| Study Period | Circulating JEV Genotype | Vaccination Status of Gilt | Incidence (Abortion * or JEV(+) **) | Relative Ratio | Vaccine Effectiveness (%) | Reference |
|---|---|---|---|---|---|---|
| 2016–2017 | GI | Yes | 41/389 * | 0.345 (0.243–0.492) | 65.5 (50.8–75.7) | This study |
| No | 65/213 * | |||||
| Yes | 6/389 ** | 0.253 (0.098–0.655) | 74.7 (34.5–90.2) | This study | ||
| No | 13/213 ** | |||||
| 1969–1970 | GIII | Yes | 1/74 * | 0.044 (0.006–0.316) | 95.6 (68.3–99.4) | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.-C.; Chen, Y.-Y.; Chen, J.-M.; Huang, C.; Huang, M.; Chiou, S.-S. Effectiveness of Live-Attenuated Genotype III Japanese Encephalitis Viral Vaccine against Circulating Genotype I Viruses in Swine. Viruses 2022, 14, 114. https://doi.org/10.3390/v14010114
Fan Y-C, Chen Y-Y, Chen J-M, Huang C, Huang M, Chiou S-S. Effectiveness of Live-Attenuated Genotype III Japanese Encephalitis Viral Vaccine against Circulating Genotype I Viruses in Swine. Viruses. 2022; 14(1):114. https://doi.org/10.3390/v14010114
Chicago/Turabian StyleFan, Yi-Chin, Yi-Ying Chen, Jo-Mei Chen, Chienjin Huang, Mei Huang, and Shyan-Song Chiou. 2022. "Effectiveness of Live-Attenuated Genotype III Japanese Encephalitis Viral Vaccine against Circulating Genotype I Viruses in Swine" Viruses 14, no. 1: 114. https://doi.org/10.3390/v14010114
APA StyleFan, Y.-C., Chen, Y.-Y., Chen, J.-M., Huang, C., Huang, M., & Chiou, S.-S. (2022). Effectiveness of Live-Attenuated Genotype III Japanese Encephalitis Viral Vaccine against Circulating Genotype I Viruses in Swine. Viruses, 14(1), 114. https://doi.org/10.3390/v14010114