
African Swine Fever in Wild Boar in Europe—A Review

 , , , , , , ,
, , , , , , ,  ,
, 
Abstract
1. Introduction
2. Clinical Signs and Pathomorphological Lesions
2.1. Clinical Signs
2.2. Gross Pathological Findings
2.3. Histopathological Findings
3. Immunology
4. Epidemiology
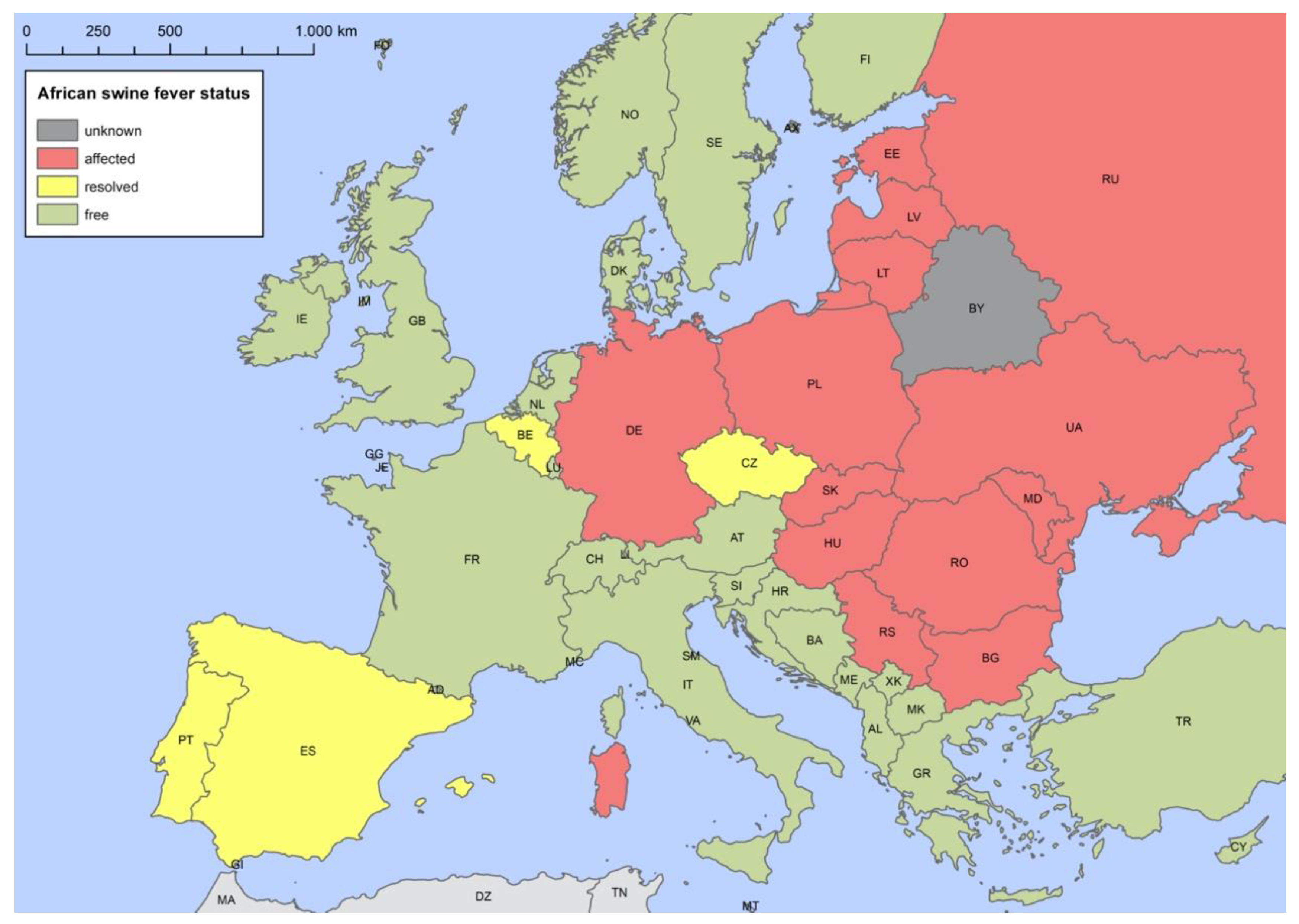
4.1. Occurrence of ASFV in Europe
4.1.1. Iberian Peninsula
4.1.2. Sardinia
4.1.3. Russian Federation (RF)
4.1.4. Baltic States, Poland and Germany
4.1.5. Czech Republic and Belgium
4.1.6. Hungary, Romania, Slovak Republic and Serbia
4.2. Tenacity of ASFV

{kind=link}
{kind=link}
{kind=link}
| Material | ASFV Stability | Method | Reference | |
|---|---|---|---|---|
| Blood, organs and tissues | Defibrinated blood (RT) | 140 days | In vivo assay | Montgomery 1921 [129] |
| Blood (−20 °C) | 6 years | In vivo assay | De Kock et al., 1940 [130] | |
| Preserved blood (4 °C) | 18 months | In vivo assay | Plowright and Parker 1967 [131] | |
| Spleen suspension (−20 °C) | 105 weeks | In vivo assay | Plowright and Parker 1967 [131] | |
| Spleen, kidney, lung (−20 °C) | 112 days | Virus isolation in macrophages | Mazur-Panasiuk and Woźniakowski 2020 [126] | |
| Spleen, lung (4 °C) | 56 days | |||
| Kidney (4 °C) | <28 days | |||
| Spleen, kidney (RT) | 7 days | |||
| Bone marrow (6–8 °C) | >6 months | In vivo assay | Kovalenko et al., 1972 [132] | |
| Bone marrow, skin (−20 °C) | 3 months | Virus isolation in macrophages | Fischer et al., 2020 [122] | |
| Bone marrow (4 °C) | 1 month | |||
| Bone marrow, muscle (RT) | <7 days | |||
| Muscle (−20 °C) | >24 months | |||
| Muscle (4 °C) | 3 months | |||
| Skin (4 °C) | 6 months | |||
| Skin (RT) | 3 months | |||
| Feces and urine | Faeces (4 °C) | 159 days | In vivo assay | Kovalenko et al., 1972 [132] |
| Urine (4 °C) | 60 days | |||
| Faeces, urine (4 °C) | 5 days | Virus isolation in macrophages | Olesen et al., 2020 [133] | |
| Faeces (4 °C and 12 °C) | 5 days | Virus isolation in macrophages | Davies et al., 2017 [124] | |
| Faeces (RT) | 3 days | |||
| Urine (4 °C, 12 °C, RT) | 5 days | |||
| Soil, water, field crops and feed | Beach sand (RT) | 14 days | Virus isolation in macrophages or cell culture | Carlson et al., 2020 [125] |
| Yard soil (RT) | 7 days | |||
| Swamp mud (RT) | 3 days | |||
| Forest soil (RT) | 0 days/none | |||
| Wet soil, leaf litter (4 °C & RT) | <3 days | Virus isolation in macrophages | Mazur-Panasiuk and Woźniakowski 2020 [126] | |
| Water (−20 °C, 4 °C, 23 °C, 37 °C) | >14 days | |||
| Water (−16 to −20 °C, 4–6 °C) | ≥60 days | Virus isolation in macrophages | Sindryakova et al., 2016 [134] | |
| Water (RT) | 50 days | |||
| Field crops (drying at RT) | <2h | Virus isolation in macrophages | Fischer et al., 2020 [127] | |
| Compound feed (−16 to −20 °C) | ≥60 days | Virus isolation in macrophages | Sindryakova et al., 2016 [134] | |
| Compound feed (4-6 °C) | 30 days | |||
| Compound feed (RT) | 1 day | |||
| Compound feed (RT) | ≥30 days | Virus isolation in macrophages, in vivo assay | Dee et al., 2018 [135] | |
| Soy oil cake (RT) | ≥30 days | |||
| Compound feed (RT) | ≥30 days | Virus isolation in macrophages | Stoian et al., 2019 [136] | |
| Soy oil cake (RT) | ≥30 days |
4.3. Transmission of ASFV among Wild Boar
4.4. Modeling
5. Diagnosis of ASF in Wild Boar
5.1. Sample Matrices
5.2. Detection of ASF Virus, ASFV Antigen and Genome
6. ASF Control in Wild Boar
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Penrith, M.L.; Thomson, G.R.; Bastos, A.D.; Phiri, O.C.; Lubisi, B.A.; Du Plessis, E.C.; Macome, F.; Pinto, F.; Botha, B.; Esterhuysen, J. An investigation into natural resistance to African swine fever in domestic pigs from an endemic area in southern Africa. Rev. Sci. Tech. 2004, 23, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M. ICTV Virus Taxonomy Profile: Asfarviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef]
- Frant, M.; Lyjak, M.; Bocian, L.; Barszcz, A.; Niemczuk, K.; Wozniakowski, G. African swine fever virus (ASFV) in Poland: Prevalence in a wild boar population (2017–2018). Vet. Med. 2020, 65, 143–158. [Google Scholar] [CrossRef]
- Nurmoja, I.; Schulz, K.; Staubach, C.; Sauter-Louis, C.; Depner, K.; Conraths, F.J.; Viltrop, A. Development of African swine fever epidemic among wild boar in Estonia—Two different areas in the epidemiological focus. Sci. Rep. 2017, 7, 12562. [Google Scholar] [CrossRef]
- Olsevskis, E.; Schulz, K.; Staubach, C.; Serzants, M.; Lamberga, K.; Pule, D.; Ozolins, J.; Conraths, F.J.; Sauter-Louis, C. African swine fever in Latvian wild boar—A step closer to elimination. Transbound. Emerg. Dis. 2020, 67, 2615–2629. [Google Scholar] [CrossRef]
- Sauter-Louis, C.; Forth, J.H.; Probst, C.; Staubach, C.; Hlinak, A.; Rudovsky, A.; Holland, D.; Schlieben, P.; Goldner, M.; Schatz, J.; et al. Joining the club: First detection of African swine fever in wild boar in Germany. Transbound. Emerg. Dis. 2020, 68, 1744–1752. [Google Scholar] [CrossRef]
- Schulz, K.; Olsevskis, E.; Staubach, C.; Lamberga, K.; Serzants, M.; Cvetkova, S.; Conraths, F.J.; Sauter-Louis, C. Epidemiological evaluation of Latvian control measures for African swine fever in wild boar on the basis of surveillance data. Sci. Rep. 2019, 9, 4189. [Google Scholar] [CrossRef]
- Wozniakowski, G.; Kozak, E.; Kowalczyk, A.; Lyjak, M.; Pomorska-Mol, M.; Niemczuk, K.; Pejsak, Z. Current status of African swine fever virus in a population of wild boar in eastern Poland (2014–2015). Arch. Virol. 2016, 161, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Laddomada, A.; Patta, C.; Oggiano, A.; Caccia, A.; Ruiu, A.; Cossu, P.; Firinu, A. Epidemiology of classical swine fever in Sardinia: A serological survey of wild boar and comparison with African swine fever. Vet. Rec. 1994, 134, 183–187. [Google Scholar] [CrossRef]
- Perez, J.; Fernandez, A.I.; Sierra, M.A.; Herraez, P.; Fernandez, A.; Martin de las Mulas, J. Serological and immunohistochemical study of African swine fever in wild boar in Spain. Vet. Rec. 1998, 143, 136–139. [Google Scholar] [CrossRef]
- Lange, M.; Siemen, H.; Blome, S.; Thulke, H.H. Analysis of spatio-temporal patterns of African swine fever cases in Russian wild boar does not reveal an endemic situation. Prev. Vet. Med. 2014, 117, 317–325. [Google Scholar] [CrossRef]
- EFSA. African Swine Fever—Scientific Opinion. EFSA J. 2015, 2015, 4163. [Google Scholar] [CrossRef]
- Schulz, K.; Staubach, C.; Blome, S.; Nurmoja, I.; Viltrop, A.; Conraths, F.J.; Kristian, M.; Sauter-Louis, C. How to Demonstrate Freedom from African Swine Fever in Wild Boars—Estonia as an Example. Vaccines 2020, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.; Staubach, C.; Blome, S.; Viltrop, A.; Nurmoja, I.; Conraths, F.J.; Sauter-Louis, C. Analysis of Estonian surveillance in wild boar suggests a decline in the incidence of African swine fever. Sci. Rep. 2019, 9, 8490. [Google Scholar] [CrossRef]
- Smietanka, K.; Wozniakowski, G.; Kozak, E.; Niemczuk, K.; Fraczyk, M.; Bocian, L.; Kowalczyk, A.; Pejsak, Z. African Swine Fever Epidemic, Poland, 2014–2015. Emerg. Infect. Dis. 2016, 22, 1201–1207. [Google Scholar] [CrossRef]
- Pautienius, A.; Schulz, K.; Staubach, C.; Grigas, J.; Zagrabskaite, R.; Buitkuviene, J.; Stankevicius, R.; Streimikyte, Z.; Oberauskas, V.; Zienius, D.; et al. African swine fever in the Lithuanian wild boar population in 2018: A snapshot. Virol. J. 2020, 17, 148. [Google Scholar] [CrossRef]
- Pejsak, Z.; Niemczuk, K.; Frant, M.; Mazur, M.; Pomorska-Mol, M.; Zietek-Barszcz, A.; Bocian, L.; Lyjak, M.; Borowska, D.; Wozniakowski, G. Four years of African swine fever in Poland. New insights into epidemiology and prognosis of future disease spread. Pol. J. Vet. Sci. 2018, 21, 835–841. [Google Scholar] [CrossRef]
- EFSA; Miteva, A.; Papanikolaou, A.; Gogin, A.; Boklund, A.; Bøtner, A.; Linden, A.; Viltrop, A.; Schmidt, C.G.; Ivanciu, C.; et al. Epidemiological analyses of African swine fever in the European Union (November 2018 to October 2019). EFSA J. 2020, 18, e05996. [Google Scholar] [CrossRef]
- OIE. Self-Declaration of Belgium’s African Swine Fever-Free Status in All Swine Species; Self-declaration submitted to the OIE on 27th October 2020 by Dr. Herman Claeyes (Federal Public Service for Public Health, Food Chain Safety and the Environment), Delegate of Belgium to the OIE, and by Dr. Jean-Francois Heymans, Chief Veterinary Officer of the Belgian veterinary services and Deputy Delegate of Belgium to the OIE; OIE: Paris, France, 2020. [Google Scholar]
- OIE. Self-Declaration of the Recovery of Freedom from African Swine Fever in All Suids by the Czech Republic; Self-declaration submitted to the OIE on 1 April 2019 by Dr. Zbynek Semerád, Director General State Veterinary Administration, Ministry of Agriculture, Czech Republic, OIE Delegate for Czech Republic; OIE: Paris, France, 2019. [Google Scholar]
- Morelle, K.; Jezek, M.; Licoppe, A.; Podgorski, T. Deathbed choice by ASF-infected wild boar can help find carcasses. Transbound. Emerg. Dis. 2019, 66, 1821–1826. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, C.; Blome, S.; Malogolovkin, A.; Parilov, S.; Kolbasov, D.; Teifke, J.P.; Beer, M. Characterization of african Swine Fever virus caucasus isolate in European wild boars. Emerg. Infect. Dis. 2011, 17, 2342–2345. [Google Scholar] [CrossRef]
- Pietschmann, J.; Guinat, C.; Beer, M.; Pronin, V.; Tauscher, K.; Petrov, A.; Keil, G.; Blome, S. Course and transmission characteristics of oral low-dose infection of domestic pigs and European wild boar with a Caucasian African swine fever virus isolate. Arch. Virol. 2015, 160, 1657–1667. [Google Scholar] [CrossRef]
- Tauscher, K.; Pietschmann, J.; Wernike, K.; Teifke, J.P.; Beer, M.; Blome, S. On the situation of African swine fever and the biological characterization of recent virus isolates. BMTW 2015, 128, 169–176. [Google Scholar]
- Zani, L.; Forth, J.H.; Forth, L.; Nurmoja, I.; Leidenberger, S.; Henke, J.; Carlson, J.; Breidenstein, C.; Viltrop, A.; Hoper, D.; et al. Deletion at the 5’-end of Estonian ASFV strains associated with an attenuated phenotype. Sci. Rep. 2018, 8, 6510. [Google Scholar] [CrossRef]
- Sehl, J.; Pikalo, J.; Schäfer, A.; Franzke, K.; Pannhorst, K.; Elnagar, A.; Blohm, U.; Blome, S.; Breithaupt, A. Comparative Pathology of Domestic Pigs and Wild Boar Infected with the Moderately Virulent African Swine Fever Virus Strain "Estonia 2014". Pathogens 2020, 9, 662. [Google Scholar] [CrossRef]
- Blome, S.; Gabriel, C.; Dietze, K.; Breithaupt, A.; Beer, M. High virulence of African swine fever virus caucasus isolate in European wild boars of all ages. Emerg. Infect. Dis. 2012, 18, 708. [Google Scholar] [CrossRef]
- Nurmoja, I.; Petrov, A.; Breidenstein, C.; Zani, L.; Forth, J.H.; Beer, M.; Kristian, M.; Viltrop, A.; Blome, S. Biological characterization of African swine fever virus genotype II strains from north-eastern Estonia in European wild boar. Transbound. Emerg. Dis. 2017, 64, 2034–2041. [Google Scholar] [CrossRef]
- Pikalo, J.; Schoder, M.E.; Sehl, J.; Breithaupt, A.; Tignon, M.; Cay, A.B.; Gager, A.M.; Fischer, M.; Beer, M.; Blome, S. The African swine fever virus isolate Belgium 2018/1 shows high virulence in European wild boar. Transbound. Emerg. Dis. 2020, 67, 1654–1659. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Nurmoja, I.; Soler, A.; Delicado, V.; Simón, A.; Martin, E.; Perez, C.; Nieto, R.; Arias, M. Evolution in Europe of African swine fever genotype II viruses from highly to moderately virulent. Vet. Microbiol. 2018, 219, 70–79. [Google Scholar] [CrossRef]
- Palliola, E.; Ioppolo, A.; Pestalozza, S. Note II: Possibility of experimental infection through ingestion and contact. Vet. Ital. 1968, 29, 371–387. [Google Scholar]
- Ravaioli, F.; Palliola, E.; Ioppolo, A. African swine fever in wild boars, Note I: Possibility of experimental infection through inoculation. Vet. Ital. 1967, 18, 508–513. [Google Scholar]
- Cabezon, O.; Munoz-Gonzalez, S.; Colom-Cadena, A.; Perez-Simo, M.; Rosell, R.; Lavin, S.; Marco, I.; Fraile, L.; de la Riva, P.M.; Rodriguez, F.; et al. African swine fever virus infection in Classical swine fever subclinically infected wild boars. BMC Vet. Res. 2017, 13, 227. [Google Scholar] [CrossRef]
- Larson, G.; Dobney, K.; Albarella, U.; Fang, M.; Matisoo-Smith, E.; Robins, J.; Lowden, S.; Finlayson, H.; Brand, T.; Willerslev, E.; et al. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication. Science 2005, 307, 1618–1621. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Fons, F.; Segales, J.; Gortazar, C. A review of viral diseases of the European wild boar: Effects of population dynamics and reservoir role. Vet. J. 2008, 176, 158–169. [Google Scholar] [CrossRef]
- Lochmiller, R.L.; Deerenberg, C. Trade-offs in evolutionary immunology: Just what is the cost of immunity? Oikos 2000, 88, 87–98. [Google Scholar] [CrossRef]
- Dei Giudici, S.; Franzoni, G.; Bonelli, P.; Bacciu, D.; Sanna, G.; Angioi, P.P.; Ledda, M.; Pilo, G.; Nicolussi, P.; Oggiano, A. Interaction of historical and modern Sardinian African swine fever viruses with porcine and wild-boar monocytes and monocyte-derived macrophages. Arch. Virol. 2019, 164, 739–745. [Google Scholar] [CrossRef]
- Oura, C.A.; Denyer, M.S.; Takamatsu, H.; Parkhouse, R.M. In vivo depletion of CD8+ T lymphocytes abrogates protective immunity to African swine fever virus. J. Gen. Virol. 2005, 86, 2445–2450. [Google Scholar] [CrossRef] [PubMed]
- Hühr, J.; Schäfer, A.; Schwaiger, T.; Zani, L.; Sehl, J.; Mettenleiter, T.C.; Blome, S.; Blohm, U. Impaired T cell responses in domestic pigs and wild boar upon infection with a highly virulent African swine fever virus strain. Transbound. Emerg. Dis. 2020, 67, 3016–3032. [Google Scholar] [CrossRef] [PubMed]
- Schafer, A.; Zani, L.; Pikalo, J.; Huhr, J.; Sehl, J.; Mettenleiter, T.C.; Breithaupt, A.; Blome, S.; Blohm, U. T-cell responses in domestic pigs and wild boar upon infection with the moderately virulent African swine fever virus strain “Estonia 2014”. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Cordón, P.J.; Jabbar, T.; Chapman, D.; Dixon, L.K.; Montoya, M. Absence of long-term protection in domestic pigs immunized with attenuated African swine fever virus isolate OURT88/3 or BeninΔMFG correlates with increased levels of regulatory T cells and IL-10. J. Virol. 2020, 14, e00350-20. [Google Scholar] [CrossRef]
- Mur, L.; Igolkin, A.; Varentsova, A.; Pershin, A.; Remyga, S.; Shevchenko, I.; Zhukov, I.; Sánchez-Vizcaíno, J.M. Detection of African Swine Fever Antibodies in Experimental and Field Samples from the Russian Federation: Implications for Control. Transbound. Emerg. Dis. 2016, 63, e436–e440. [Google Scholar] [CrossRef] [PubMed]
- Pikalo, J.; Zani, L.; Huhr, J.; Beer, M.; Blome, S. Pathogenesis of African swine fever in domestic pigs and European wild boar—Lessons learned from recent animal trials. Virus Res. 2019, 271, 197614. [Google Scholar] [CrossRef]
- Martinez-Aviles, M.; Iglesias, I.; De La Torre, A. Evolution of the ASF Infection Stage in Wild Boar within the EU (2014–2018). Front. Vet. Sci. 2020, 7, 155. [Google Scholar] [CrossRef] [PubMed]
- Walczak, M.; Frant, M.; Juszkiewicz, M.; Mazur-Panasiuk, N.; Szymankiewicz, K.; Bruczynska, M.; Wozniakowski, G. Vertical transmission of anti-ASFV antibodies as one of potential causes of seropositive results among young wild boar population in Poland. Pol. J. Vet. Sci. 2020, 23, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.; Forth, J.H.; Zani, L.; Beer, M.; Blome, S. No evidence for long-term carrier status of pigs after African swine fever virus infection. Transbound. Emerg. Dis. 2018, 65, 1318–1328. [Google Scholar] [CrossRef]
- Escribano, J.M.; Galindo, I.; Alonso, C. Antibody-mediated neutralization of African swine fever virus: Myths and facts. Virus Res. 2013, 173, 101–109. [Google Scholar] [CrossRef]
- Onisk, D.V.; Borca, M.V.; Kutish, G.; Kramer, E.; Irusta, P.; Rock, D.L. Passively transferred African swine fever virus antibodies protect swine against lethal infection. Virology 1994, 198, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhao, D.; He, X.; Liu, R.; Wang, Z.; Zhang, X.; Li, F.; Shan, D.; Chen, H.; Zhang, J.; et al. A seven-gene-deleted African swine fever virus is safe and effective as a live attenuated vaccine in pigs. Sci. China Life Sci. 2020, 63, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Costard, S.; Wieland, B.; de Glanville, W.; Jori, F.; Rowlands, R.; Vosloo, W.; Roger, F.; Pfeiffer, D.U.; Dixon, L.K. African swine fever: How can global spread be prevented? Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2683–2696. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, A.; Sotgia, S.; Patta, C.; Oggiano, A.; Carboni, A.; Cossu, P.; Laddomada, A. Temporal and spatial patterns of African swine fever in Sardinia. Prev. Vet. Med. 1998, 35, 297–306. [Google Scholar] [CrossRef]
- Vepkhvadze, N.G.; Menteshashvili, I.; Kokhreidze, M.; Goginashvili, K.; Tigilauri, T.; Mamisashvili, E.; Gelashvili, L.; Abramishvili, T.; Donduashvili, M.; Ghvinjilia, G.; et al. Active surveillance of African swine fever in domestic swine herds in Georgia, 2014. Rev. Sci. Tech. 2017, 36, 879–887. [Google Scholar] [CrossRef]
- FAO. African Swine Fever in the Caucasus; FAO Empres Watch: Rome, Italy, 2008. [Google Scholar]
- Markosyan, T.; Sargsyan, K.; Kharatyan, S.; Elbakyan, H.; Hakobyan, V.; Simonyan, L.; Voskanyan, H.; Shirvanyan, A.; Stepanyan, T.; Khachatryan, M.; et al. The epidemiological status of African swine fever in domestic swine herds in the Tavush Province region, Armenia. Rev. Sci. Tech. 2019, 38, 751–760. [Google Scholar] [CrossRef]
- Gogin, A.; Gerasimov, V.; Malogolovkin, A.; Kolbasov, D. African swine fever in the North Caucasus region and the Russian Federation in years 2007–2012. Virus Res. 2013, 173, 198–203. [Google Scholar] [CrossRef]
- GF-TADs. Expert Mission on African Swine Fever in Belarus; FAO, OIE. 2015. Available online: https://rr-europe.oie.int/wp-content/uploads/2020/04/2015-04_sge-asf_belarus_en.pdf (accessed on 25 August 2021).
- DEFRA. African Swine Fever in Wild Boar in Ukraine; London, UK. 2014. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/339703/poa-asf-ukraine-20140108.pdf (accessed on 25 August 2021).
- State Food and Veterinary Service. African Swine Fever in Lithuania—Update; Presentation at the Standing Committee on Plants, Animals; Food and Feed (SCoPAFF): Brussels, Belgium, 2014.
- Poland, G.V.I. African Swine Fever in Poland—Update. Available online: https://ec.europa.eu/food/system/files/2018-01/reg-com_ahw_20180117_asf_pol.pdf (accessed on 15 June 2021).
- Mazur-Panasiuk, N.; Walczak, M.; Juszkiewicz, M.; Wozniakowski, G. The Spillover of African Swine Fever in Western Poland Revealed Its Estimated Origin on the Basis of O174L, K145R, MGF 505-5R and IGR I73R/I329L Genomic Sequences. Viruses 2020, 12, 1094. [Google Scholar] [CrossRef]
- Olsevskis, E.; Guberti, V.; Serzants, M.; Westergaard, J.; Gallardo, C.; Rodze, I.; Depner, K. African swine fever virus introduction into the EU in 2014: Experience of Latvia. Res. Vet. Sci. 2016, 105, 28–30. [Google Scholar] [CrossRef]
- GF-TADs. Expert Mission on African Swine Fever in Moldova. 2016. Available online: https://rr-europe.oie.int/wp-content/uploads/2020/04/2016-10_sge-asf_moldova_en.pdf (accessed on 25 August 2021).
- EFSA (European Food Safety Authority, Panel on Animal Health and Welfare). Epidemiological analyses of African swine fever in the European Union (November 2018 to October 2019). EFSA J. 2020, 18, 5996. [Google Scholar] [CrossRef]
- Zani, L.; Dietze, K.; Dimova, Z.; Forth, J.H.; Denev, D.; Depner, K.; Alexandrov, T. African Swine Fever in a Bulgarian Backyard Farm-A Case Report. Vet. Sci. 2019, 6, 94. [Google Scholar] [CrossRef]
- Linden, A.; Licoppe, A.; Volpe, R.; Paternostre, J.; Lesenfants, C.; Cassart, D.; Garigliany, M.; Tignon, M.; van den Berg, T.; Desmecht, D.; et al. Summer 2018: African swine fever virus hits north-western Europe. Transbound. Emerg. Dis. 2019, 66, 54–55. [Google Scholar] [CrossRef] [PubMed]
- Reuters. Serbia Reports Outbreak of African Swine Fever in Boars. Available online: https://www.reuters.com/article/us-serbia-swineflu/serbia-reports-outbreak-of-african-swine-fever-in-boars-idUSKBN1ZE1VQ (accessed on 15 June 2021).
- Mur, L.; Martinez-Lopez, B.; Martinez-Aviles, M.; Costard, S.; Wieland, B.; Pfeiffer, D.U.; Sanchez-Vizcaino, J.M. Quantitative risk assessment for the introduction of African swine fever virus into the European Union by legal import of live pigs. Transbound. Emerg. Dis. 2012, 59, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.; Boadella, M.; Martinez-Lopez, B.; Gallardo, C.; Gortazar, C.; Sanchez-Vizcaino, J.M. Monitoring of African swine fever in the wild boar population of the most recent endemic area of Spain. Transbound. Emerg. Dis. 2012, 59, 526–531. [Google Scholar] [CrossRef]
- Torresi, C.; Fiori, M.; Bertolotti, L.; Floris, M.; Colitti, B.; Giammarioli, M.; Dei Giudici, S.; Oggiano, A.; Malmberg, M.; De Mia, G.M.; et al. The evolution of African swine fever virus in Sardinia (1978–2014) as revealed by whole-genome sequencing and comparative analysis. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.; Atzeni, M.; Martinez-Lopez, B.; Feliziani, F.; Rolesu, S.; Sanchez-Vizcaino, J.M. Thirty-Five-Year Presence of African Swine Fever in Sardinia: History, Evolution and Risk Factors for Disease Maintenance. Transbound. Emerg. Dis. 2016, 63, e165–e177. [Google Scholar] [CrossRef]
- Cappai, S.; Rolesu, S.; Coccollone, A.; Laddomada, A.; Loi, F. Evaluation of biological and socio-economic factors related to persistence of African swine fever in Sardinia. Prev. Vet. Med. 2018, 152, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Viltrop, A.; Boinas, F.S.; Depner, K.; Jori, F.; Kolbasov, D.; Laddomada, A.; Stahl, K.; Chenais, E. African swine fever epidemiology, surveillance and control. In Understanding and Combatting African SWINE Fever, A European Perspective; Iacolina, L., Penrith, M.L., Bellini, S., Chenais, E., Jori, F., Montoya, M., Stahl, K., Gavier-Widen, D., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2021; pp. 243–249. [Google Scholar]
- Jurado, C.; Fernandez-Carrion, E.; Mur, L.; Rolesu, S.; Laddomada, A.; Sanchez-Vizcaino, J.M. Why is African swine fever still present in Sardinia? Transbound. Emerg. Dis. 2018, 65, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Laddomada, A.; Rolesu, S.; Loi, F.; Cappai, S.; Oggiano, A.; Madrau, M.P.; Sanna, M.L.; Pilo, G.; Bandino, E.; Brundu, D.; et al. Surveillance and control of African Swine Fever in free-ranging pigs in Sardinia. Transbound. Emerg. Dis. 2019, 66, 1114–1119. [Google Scholar] [CrossRef]
- Iglesias, I.; Rodriguez, A.; Feliziani, F.; Rolesu, S.; de la Torre, A. Spatio-temporal Analysis of African Swine Fever in Sardinia (2012–2014): Trends in Domestic Pigs and Wild Boar. Transbound. Emerg. Dis. 2017, 64, 656–662. [Google Scholar] [CrossRef]
- Firinu, A.; Scarano, C. African swine fever and classical swine fever (hog cholera) among wild boar in Sardinia. Rev. Sci. Tech. Off. Int. Epiz. 1988, 7, 909–915. [Google Scholar] [CrossRef]
- Martinez-Lopez, B.; Perez, A.M.; Feliziani, F.; Rolesu, S.; Mur, L.; Sanchez-Vizcaino, J.M. Evaluation of the risk factors contributing to the African swine fever occurrence in Sardinia, Italy. Front. Microbiol. 2015, 6, 314. [Google Scholar] [CrossRef]
- Cadenas-Fernandez, E.; Sanchez-Vizcaino, J.M.; Pintore, A.; Denurra, D.; Cherchi, M.; Jurado, C.; Vicente, J.; Barasona, J.A. Free-Ranging Pig and Wild Boar Interactions in an Endemic Area of African Swine Fever. Front. Vet. Sci. 2019, 6, 376. [Google Scholar] [CrossRef]
- Bosch, J.; Barasona, J.A.; Cadenas-Fernandez, E.; Jurado, C.; Pintore, A.; Denurra, D.; Cherchi, M.; Vicente, J.; Sanchez-Vizcaino, J.M. Retrospective spatial analysis for African swine fever in endemic areas to assess interactions between susceptible host populations. PLoS ONE 2020, 15, e0233473. [Google Scholar] [CrossRef]
- Oganesyan, A.S.; Petrova, O.N.; Korennoy, F.I.; Bardina, N.S.; Gogin, A.E.; Dudnikov, S.A. African swine fever in the Russian Federation: Spatio-temporal analysis and epidemiological overview. Virus Res. 2013, 173, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Sargsyan, M.A.; Voskanyan, H.E.; Karalova, E.M.; Hakobyan, L.H.; Karalyan, Z.A. Third wave of African swine fever infection in Armenia: Virus demonstrates the reduction of pathogenicity. Vet. World 2018, 11, 5–9. [Google Scholar] [CrossRef]
- Kolbasov, D.; Titov, I.; Tsybanov, S.; Gogin, A.; Malogolovkin, A. African Swine Fever Virus, Siberia, Russia, 2017. Emerg. Infect. Dis. 2018, 24, 796–798. [Google Scholar] [CrossRef]
- Iglesias, I.; Munoz, M.J.; Montes, F.; Perez, A.; Gogin, A.; Kolbasov, D.; de la Torre, A. Reproductive Ratio for the Local Spread of African Swine Fever in Wild Boars in the Russian Federation. Transbound. Emerg. Dis. 2016, 63, e237–e245. [Google Scholar] [CrossRef] [PubMed]
- Vergne, T.; Gogin, A.; Pfeiffer, D.U. Statistical Exploration of Local Transmission Routes for African Swine Fever in Pigs in the Russian Federation, 2007–2014. Transbound. Emerg. Dis. 2017, 64, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.; Jurado, C.; Gallardo, C.; Fernandez-Pinero, J.; Sanchez-Vizcaino, J.M. Gaps in African swine fever: Analysis and priorities. Transbound. Emerg. Dis. 2018, 65, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Kramer, M.; Höreth-Böntgen, D.; Conraths, F.J.; Depner, K.; Böhle, W.; Gall, Y.; Selhorst, T.; Micklich, M.; Staubach, C.; et al. Die Afrikanische Schweinepest in Osteuropa—Eine Gefahr auch für deutsche Schweinebestände? Tierärztl. Umsch. 2011, 66, 291–296. [Google Scholar]
- Blokhin, A.; Toropova, N.; Burova, O.; Sevskikh, T.; Gogin, A.; Debeljak, Z.; Zakharova, O. Spatio-Temporal Analysis of the Spread of ASF in the Russian Federation in 2017–2019. Acta Vet.-Beog. 2020, 70, 194–206. [Google Scholar] [CrossRef]
- Maciulskis, P.; Masiulis, M.; Pridotkas, G.; Buitkuviene, J.; Jurgelevicius, V.; Jaceviciene, I.; Zagrabskaite, R.; Zani, L.; Pileviciene, S. The African Swine Fever Epidemic in Wild Boar (Sus scrofa) in Lithuania (2014–2018). Vet. Sci. 2020, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Pautienius, A.; Grigas, J.; Pileviciene, S.; Zagrabskaite, R.; Buitkuviene, J.; Pridotkas, G.; Stankevicius, R.; Streimikyte, Z.; Salomskas, A.; Zienius, D.; et al. Prevalence and spatiotemporal distribution of African swine fever in Lithuania, 2014–2017. Virol. J. 2018, 15, 177. [Google Scholar] [CrossRef]
- Schulz, K.; Masiulis, M.; Staubach, C.; Malakauskas, A.; Pridotkas, G.; Conraths, F.J.; Sauter-Louis, C. African Swine Fever and Its Epidemiological Course in Lithuanian Wild Boar. Viruses 2021, 13, 1276. [Google Scholar] [CrossRef] [PubMed]
- Pejsak, Z.; Truszczynski, M.; Niemczuk, K.; Kozak, E.; Markowska-Daniel, I. Epidemiology of African Swine Fever in Poland since the detection of the first case. Pol. J. Vet. Sci. 2014, 17, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Cwynar, P.; Stojkov, J.; Wlazlak, K. African Swine Fever Status in Europe. Viruses 2019, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Deng, X.; Chen, J.; Wang, J.; Chen, Q.; Niu, B. Risk analysis of African swine fever in Poland based on spatio-temporal pattern and Latin hypercube sampling, 2014–2017. BMC Vet. Res. 2019, 15, 160. [Google Scholar] [CrossRef]
- Sauter-Louis, C.; Schulz, K.; Richter, M.; Staubach, C.; Mettenleiter, T.C.; Conraths, F.J. African swine fever: Why the situation in Germany is not comparable to that in the Czech Republic or Belgium. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Chenais, E.; Stahl, K.; Guberti, V.; Depner, K. Identification of Wild Boar-Habitat Epidemiologic Cycle in African Swine Fever Epizootic. Emerg. Infect. Dis. 2018, 24, 810–812. [Google Scholar] [CrossRef]
- Guinat, C.; Gogin, A.; Blome, S.; Keil, G.; Pollin, R.; Pfeiffer, D.U.; Dixon, L. Transmission routes of African swine fever virus to domestic pigs: Current knowledge and future research directions. Vet. Rec. 2016, 178, 262–267. [Google Scholar] [CrossRef]
- Khomenko, S.; Beltán-Alcrudo, D.; Rozstalnyy, A.; Gogin, A.; Kolbasov, D.; Pinto, J.; Lubroth, J.; Martin, V. African Swine Fever in the Russian Federation: Risk Factors for Europe and Beyond. EMPRES Watch 2013, 28, 1–14. [Google Scholar]
- Schulz, K.; Conraths, F.J.; Blome, S.; Staubach, C.; Sauter-Louis, C. African Swine Fever: Fast and Furious or Slow and Steady? Viruses 2019, 11, 866. [Google Scholar] [CrossRef]
- Podgorski, T.; Smietanka, K. Do wild boar movements drive the spread of African Swine Fever? Transbound. Emerg. Dis. 2018, 65, 1588–1596. [Google Scholar] [CrossRef]
- EFSA. Epidemiological analyses on African swine fever in the Baltic countries and Poland. EFSA J. 2017, 15, e04732. [Google Scholar] [CrossRef]
- Podgórski, T.; Apollonio, M.; Keuling, O. Contact Rates in Wild Boar Populations: Implications for Disease Transmission. J. Wildl. Manag. 2018, 82, 1210–1218. [Google Scholar] [CrossRef]
- Schulz, K.; Conraths, F.J.; Staubach, C.; Viltrop, A.; Olsevskis, E.; Nurmoja, I.; Lamberga, K.; Sauter-Louis, C. To sample or not to sample? Detection of African swine fever in wild boar killed in road traffic accidents. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Chenais, E.; Depner, K.; Guberti, V.; Dietze, K.; Viltrop, A.; Stahl, K. Epidemiological considerations on African swine fever in Europe 2014–2018. Porc. Health Manag. 2019, 5, 6. [Google Scholar] [CrossRef]
- Depner, K.; Gortazar, C.; Guberti, V.; Masiulis, M.; More, S.; Olsevskis, E.; Thulke, H.H.; Viltrop, A.; Wozniakowski, G.; Cortinas, A.; et al. Epidemiological analyses of African swine fever in the Baltic States and Poland (Update September 2016–September 2017). EFSA J. 2017, 15, e05068. [Google Scholar] [CrossRef] [PubMed]
- Podgorski, T.; Borowik, T.; Lyjak, M.; Wozniakowski, G. Spatial epidemiology of African swine fever: Host, landscape and anthropogenic drivers of disease occurrence in wild boar. Prev. Vet. Med. 2020, 177, 104691. [Google Scholar] [CrossRef] [PubMed]
- Satran, P. From ASF Infection in Wild Boar to Eradication and Free Status Recovery in the Czech Republic. Available online: http://web.oie.int/RR-Europe/eng/Regprog/docs/docs/SGE%20ASF12/17_CZ_detailed_situation.pdf (accessed on 21 April 2020).
- Satran, P.; Jarosil, T.; Semerad, Z. African swine fever in wild boar in the Czech Republic. In Proceedings of the 12th International Symposium on wild boar and other suids, Láznĕ Bĕlohrad, Czech Republic, 4–7 September 2018. [Google Scholar]
- Vaclavek, P.; Bartak, P. ASF in the Czech Republic: Experience from the NRL. Available online: http://web.oie.int/RR-Europe/eng/Regprog/docs/docs/SGE%20ASF12/BTSF/02_Sampling_and_laboratory_testing_Pavel_Bartak.pdf (accessed on 21 April 2020).
- Vaclavek, P. ASF in the Czech Republic: Management Experience and Lessons Learnt. Available online: http://www.fao.org/fileadmin/user_upload/reu/europe/documents/events2019/ASF_reg/06.pdf (accessed on 20 July 2020).
- Cukor, J.; Linda, R.; Vaclavek, P.; Satran, P.; Mahlerova, K.; Vacek, Z.; Kunca, T.; Havranek, F. Wild boar deathbed choice in relation to ASF: Are there any differences between positive and negative carcasses? Prev. Vet. Med. 2020, 177, 104943. [Google Scholar] [CrossRef] [PubMed]
- Marcon, A.; Linden, A.; Satran, P.; Gervasi, V.; Licoppe, A.; Guberti, V. R0 Estimation for the African Swine Fever Epidemics in Wild Boar of Czech Republic and Belgium. Vet. Sci. 2019, 7, 2. [Google Scholar] [CrossRef]
- Garigliany, M.; Desmecht, D.; Tignon, M.; Cassart, D.; Lesenfant, C.; Paternostre, J.; Volpe, R.; Cay, A.B.; van den Berg, T.; Linden, A. Phylogeographic Analysis of African Swine Fever Virus, Western Europe, 2018. Emerg. Infect. Dis. 2019, 25, 184–186. [Google Scholar] [CrossRef]
- Forth, J.H.; Tignon, M.; Cay, A.B.; Forth, L.F.; Hoper, D.; Blome, S.; Beer, M. Comparative Analysis of Whole-Genome Sequence of African Swine Fever Virus Belgium 2018/1. Emerg. Infect. Dis. 2019, 25, 1249–1252. [Google Scholar] [CrossRef]
- Gilliaux, G.; Garigliany, M.; Licoppe, A.; Paternostre, J.; Lesenfants, C.; Linden, A.; Desmecht, D. Newly emerged African swine fever virus strain Belgium/Etalle/wb/2018: Complete genomic sequence and comparative analysis with reference p72 genotype II strains. Transbound. Emerg. Dis. 2019, 66, 2566–2591. [Google Scholar] [CrossRef] [PubMed]
- AFSCA. African Swine Fever. Available online: http://www.afsca.be/asp/aktualitat/belgien/ (accessed on 13 April 2020).
- AFSCA. African Swine Fever in Wild Boar in Belgium. Available online: https://ec.europa.eu/food/system/files/2020-11/reg-com_ahw_20201119_pres_asf_bel.pdf (accessed on 21 June 2021).
- Dellicour, S.; Desmecht, D.; Paternostre, J.; Mealengreaux, C.; Licoppe, A.; Gilbert, A.; Linden, A. Unravelling the dispersal dynamics and ecological drivers of the African swine fever outbreak in Belgium. J. Appl. Ecol. 2020. [Google Scholar] [CrossRef]
- Ministry of Agriculture Hungary. The First Occurrence of African Swine Fever in Wild Boars in Hungary; Meeting of the Standing Committee of Plants, Animals; Food and Feed: Brussels, Belgium, 2018. [Google Scholar]
- Guberti, V.; Khomenko, S.; Masiulis, M.; Kerba, S. African swine fever in wild boar ecology and biosecurity. In FAO Animal Production and Health Manual No. 22; FAO, OIE and EC.: Rome, Italy, 2019. [Google Scholar]
- Pepin, K.M.; Golnar, A.J.; Abdo, Z.; Podgorski, T. Ecological drivers of African swine fever virus persistence in wild boar populations: Insight for control. Ecol. Evol. 2020, 10, 2846–2859. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.; Bicout, D.; Boklund, A.; Bøtner, A.; Depner, K.; More, S.J.; Roberts, H.; Stahl, K.; Thulke, H.; Viltrop, A.; et al. Research Gap Analysis on African Swine Fever. EFSA J. 2019, 17, 5811. [Google Scholar] [CrossRef]
- Fischer, M.; Hühr, J.; Blome, S.; Conraths, F.J.; Probst, C. Stability of African Swine Fever Virus in Carcasses of Domestic Pigs and Wild Boar Experimentally Infected with the ASFV “Estonia 2014” Isolate. Viruses 2020, 12, 1118. [Google Scholar] [CrossRef] [PubMed]
- Kovalenko, J.R.; Sidorov, M.A.; Burba, L.G. Afrikanskaia Chuma Svinei. Afr. Swine Fever 1972, unpublished manuscript translated from Russian to German. [Google Scholar]
- Davies, K.; Goatley, L.C.; Guinat, C.; Netherton, C.L.; Gubbins, S.; Dixon, L.K.; Reis, A.L. Survival of African Swine Fever Virus in Excretions from Pigs Experimentally Infected with the Georgia 2007/1 Isolate. Transbound. Emerg. Dis. 2017, 64, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; Fischer, M.; Zani, L.; Eschbaumer, M.; Fuchs, W.; Mettenleiter, T.; Beer, M.; Blome, S. Stability of African Swine Fever Virus in Soil and Options to Mitigate the Potential Transmission Risk. Pathogens 2020, 9, 977. [Google Scholar] [CrossRef] [PubMed]
- Mazur-Panasiuk, N.; Wozniakowski, G. Natural inactivation of African swine fever virus in tissues: Influence of temperature and environmental conditions on virus survival. Vet. Microbiol. 2020, 242, 108609. [Google Scholar] [CrossRef]
- Fischer, M.; Mohnke, M.; Probst, C.; Pikalo, J.; Conraths, F.J.; Beer, M.; Blome, S. Stability of African swine fever virus on heat-treated field crops. Transbound. Emerg. Dis. 2020, 67, 2318–2323. [Google Scholar] [CrossRef] [PubMed]
- Zani, L.; Masiulis, M.; Busauskas, P.; Dietze, K.; Pridotkas, G.; Globig, A.; Blome, S.; Mettenleiter, T.; Depner, K.; Karveliene, B. African swine fever virus survival in buried wild boar carcasses. Transbound. Emerg. Dis. 2020, 67, 2086–2092. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.E. On A Form of Swine Fever Occurring in British East Africa (Kenya Colony). J. Comp. Pathol. 1921, 34, 159–191. [Google Scholar] [CrossRef]
- De Kock, G.; Robinson, E.M.; Keppel, J.J.G. Swine Fever in South Africa. Onderstepoort J. Vet. Res. 1940, 14, 31–93. [Google Scholar]
- Plowright, W.; Parker, J. The stability of African swine fever virus with particular reference to heat and pH inactivation. Arch. Gesamte Virusforsch. 1967, 21, 383–402. [Google Scholar] [CrossRef]
- Kovalenko, J.R.; Sidorov, M.A.; Burba, L.G. Afrikanskaia Chuma Svinei. Ph.D. Thesis, University of Moskau, Moskau, Russia, 1972. translated dissertation. [Google Scholar]
- Olesen, A.S.; Belsham, G.J.; Bruun Rasmussen, T.; Lohse, L.; Bodker, R.; Halasa, T.; Boklund, A.; Botner, A. Potential routes for indirect transmission of African swine fever virus into domestic pig herds. Transbound. Emerg. Dis. 2020, 67, 1472–1484. [Google Scholar] [CrossRef] [PubMed]
- Sindryakova, L.P.; Morgunov, Y.P.; Chichikin, A.Y.; Gazaev, I.; Kudryashov, D.A.; Tsybanov, S. The influence of temperature on the Russion isolate of African swine fever virus in pork products and feed with extrapolation to natural conditions. Agric. Biol. 2016, 51, 467–474. [Google Scholar]
- Dee, S.A.; Bauermann, F.V.; Niederwerder, M.C.; Singrey, A.; Clement, T.; de Lima, M.; Long, C.; Patterson, G.; Sheahan, M.A.; Stoian, A.M.M.; et al. Survival of viral pathogens in animal feed ingredients under transboundary shipping models. PLoS ONE 2018, 13, e0194509. [Google Scholar] [CrossRef]
- Stoian, A.M.M.; Zimmerman, J.; Ji, J.; Hefley, T.J.; Dee, S.; Diel, D.G.; Rowland, R.R.R.; Niederwerder, M.C. Half-Life of African Swine Fever Virus in Shipped Feed. Emerg. Infect. Dis. 2019, 25, 2261–2263. [Google Scholar] [CrossRef] [PubMed]
- Plowright, W.; Perry, C.T.; Peirce, M.A. Transovarial infection with African swine fever virus in the argasid tick, Ornithodoros moubata porcinus, Walton. Res. Vet. Sci. 1970, 11, 582–584. [Google Scholar] [CrossRef]
- Jori, F.; Bastos, A.D. Role of wild suids in the epidemiology of African swine fever. Ecohealth 2009, 6, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Jori, F.; Vial, L.; Penrith, M.L.; Perez-Sanchez, R.; Etter, E.; Albina, E.; Michaud, V.; Roger, F. Review of the sylvatic cycle of African swine fever in sub-Saharan Africa and the Indian ocean. Virus Res. 2013, 173, 212–227. [Google Scholar] [CrossRef]
- Boinas, F.S.; Wilson, A.J.; Hutchings, G.H.; Martins, C.; Dixon, L.J. The persistence of African swine fever virus in field-infected Ornithodoros erraticus during the ASF endemic period in Portugal. PLoS ONE 2011, 6, e20383. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on African Swine Fever. EFSA J. 2010, 8, 1556. [Google Scholar] [CrossRef]
- de Carvalho Ferreira, H.C.; Weesendorp, E.; Quak, S.; Stegeman, J.A.; Loeffen, W.L. Suitability of faeces and tissue samples as a basis for non-invasive sampling for African swine fever in wild boar. Vet. Microbiol. 2014, 172, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Forth, J.H.; Amendt, J.; Blome, S.; Depner, K.; Kampen, H. Evaluation of blowfly larvae (Diptera: Calliphoridae) as possible reservoirs and mechanical vectors of African swine fever virus. Transbound. Emerg. Dis. 2018, 65, e210–e213. [Google Scholar] [CrossRef]
- Herm, R.; Kirik, H.; Vilem, A.; Zani, L.; Forth, J.H.; Muller, A.; Michelitsch, A.; Wernike, K.; Werner, D.; Tummeleht, L.; et al. No evidence for african swine fever virus DNA in haematophagous arthropods collected at wild boar baiting sites in Estonia. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Mellor, P.S.; Kitching, R.P.; Wilkinson, P.J. Mechanical transmission of capripox virus and African swine fever virus by Stomoxys calcitrans. Res. Vet. Sci. 1987, 43, 109–112. [Google Scholar] [CrossRef]
- Herm, R.; Tummeleht, L.; Jurison, M.; Vilem, A.; Viltrop, A. Trace amounts of African swine fever virus DNA detected in insects collected from an infected pig farm in Estonia. Vet. Med. Sci. 2020, 6, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Turcinaviciene, J.; Petrasiunas, A.; Bernotiene, R.; Masiulis, M.; Jonusaitis, V. The contribution of insects to African swine fever virus dispersal: Data from domestic pig farms in Lithuania. Med. Vet. Entomol. 2020, 35, 484–489. [Google Scholar] [CrossRef] [PubMed]
- FAO. African Swine Fever in Georgia; FAO: Rome, Italy, 2007; Available online: http://www.fao.org/docs/eims/upload/230205/EW_ASF_Georgia_Jun07.pdf (accessed on 25 August 2021).
- CVET. Mission of the Community Veterinary Emergency Team (CVET) to Poland; European Commission, Brussels, 2017. Available online: https://ec.europa.eu/food/sites/food/files/animals/docs/reg-com_ahw_20171130_asf_cvet-mission_pol.pdf (accessed on 25 August 2021).
- Gauntlett, F.; Roberts, H.; Smith, J.; Gale, P. African Swine Fever in Wild Boar in Belgium—Preliminiary Outbreak Assessment; 2018. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/740925/poa-asf-boar-belgium.pdf (accessed on 25 August 2021).
- Gulenkin, V.M.; Korennoy, F.I.; Karaulov, A.K.; Dudnikov, S.A. Cartographical analysis of African swine fever outbreaks in the territory of the Russian Federation and computer modeling of the basic reproduction ratio. Prev. Vet. Med. 2011, 102, 167–174. [Google Scholar] [CrossRef]
- Boklund, A.; Cay, B.; Depner, K.; Foldi, Z.; Guberti, V.; Masiulis, M.; Miteva, A.; More, S.; Olsevskis, E.; Satran, P.; et al. Epidemiological analyses of African swine fever in the European Union (November 2017 until November 2018). EFSA J. 2018, 16, e05494. [Google Scholar] [CrossRef]
- Halasa, T.; Boklund, A.; Botner, A.; Mortensen, S.; Kjaer, L.J. Simulation of transmission and persistence of African swine fever in wild boar in Denmark. Prev. Vet. Med. 2019, 167, 68–79. [Google Scholar] [CrossRef]
- O’Neill, X.; White, A.; Ruiz-Fons, F.; Gortazar, C. Modelling the transmission and persistence of African swine fever in wild boar in contrasting European scenarios. Sci. Rep. 2020, 10, 5895. [Google Scholar] [CrossRef] [PubMed]
- Depner, K.; Staubach, C.; Probst, C.; Globig, A.; Blome, S.; Dietze, K.; Sauter-Louis, C.; Conraths, F.J. African Swine fever—Epidemiological considerations and consequences for disease control. TU 2016, 71, 72–78. [Google Scholar]
- Sanchez-Cordon, P.J.; Nunez, A.; Neimanis, A.; Wikstrom-Lassa, E.; Montoya, M.; Crooke, H.; Gavier-Widen, D. African Swine Fever: Disease Dynamics in Wild Boar Experimentally Infected with ASFV Isolates Belonging to Genotype I and II. Viruses 2019, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Probst, C.; Globig, A.; Knoll, B.; Conraths, F.J.; Depner, K. Behaviour of free ranging wild boar towards their dead fellows: Potential implications for the transmission of African swine fever. R. Soc. Open Sci. 2017, 4, 170054. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.R.; Bevins, S.N. A Review of African Swine Fever and the Potential for Introduction into the United States and the Possibility of Subsequent Establishment in Feral Swine and Native Ticks. Front. Vet. Sci. 2018, 5, 11. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority, Panel on Animal Health and Welfare). Research gap analysis on African swine fever. EFSA J. 2019, 17, 5811. [Google Scholar] [CrossRef]
- Probst, C.; Gethmann, J.; Amendt, J.; Lutz, L.; Teifke, J.P.; Conraths, F.J. Estimating the Postmortem Interval of Wild Boar Carcasses. Vet. Sci. 2020, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Probst, C.; Gethmann, J.; Hohmann, U.; Knoll, B.; Amendt, J.; Teifke, J.P.; Conraths, F.J. Zersetzungsstadien bei Wildschweinkadavern—Und wie die Liegezeit geschätzt werden kann. Amtstierärztl. Dienst. Lebensm. 2020, 2, 85–94. [Google Scholar]
- Probst, C.; Gethmann, J.; Amler, S.; Globig, A.; Knoll, B.; Conraths, F.J. The potential role of scavengers in spreading African swine fever among wild boar. Sci. Rep. 2019, 9, 11450. [Google Scholar] [CrossRef]
- Bosch, J.; Rodriguez, A.; Iglesias, I.; Munoz, M.J.; Jurado, C.; Sanchez-Vizcaino, J.M.; de la Torre, A. Update on the Risk of Introduction of African Swine Fever by Wild Boar into Disease-Free European Union Countries. Transbound. Emerg. Dis. 2017, 64, 1424–1432. [Google Scholar] [CrossRef] [PubMed]
- Andraud, M.; Halasa, T.; Boklund, A.; Rose, N. Threat to the French Swine Industry of African Swine Fever: Surveillance, Spread, and Control Perspectives. Front. Vet. Sci. 2019, 6, 248. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.A.; Condoleo, R.; Simons, R.R.L.; Gale, P.; Kelly, L.A.; Snary, E.L. The Risk of Infection by African Swine Fever Virus in European Swine Through Boar Movement and Legal Trade of Pigs and Pig Meat. Front. Vet. Sci. 2019, 6, 486. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.; Martinez-Lopez, B.; Sanchez-Vizcaino, J.M. Risk of African swine fever introduction into the European Union through transport-associated routes: Returning trucks and waste from international ships and planes. BMC Vet. Res. 2012, 8, 149. [Google Scholar] [CrossRef]
- Nigsch, A.; Costard, S.; Jones, B.A.; Pfeiffer, D.U.; Wieland, B. Stochastic spatio-temporal modelling of African swine fever spread in the European Union during the high risk period. Prev. Vet. Med. 2013, 108, 262–275. [Google Scholar] [CrossRef]
- Mur, L.; Martinez-Lopez, B.; Costard, S.; de la Torre, A.; Jones, B.A.; Martinez, M.; Sanchez-Vizcaino, F.; Munoz, M.J.; Pfeiffer, D.U.; Sanchez-Vizcaino, J.M.; et al. Modular framework to assess the risk of African swine fever virus entry into the European Union. BMC Vet. Res. 2014, 10, 145. [Google Scholar] [CrossRef]
- de Vos, C.J.; Taylor, R.A.; Simons, R.R.L.; Roberts, H.; Hulten, C.; de Koeijer, A.A.; Lyytikainen, T.; Napp, S.; Boklund, A.; Petie, R.; et al. Cross-Validation of Generic Risk Assessment Tools for Animal Disease Incursion Based on a Case Study for African Swine Fever. Front. Vet. Sci. 2020, 7, 56. [Google Scholar] [CrossRef]
- Alexander, N.S.; Massei, G.; Wint, W. The European Distribution of Sus Scrofa. Model Outputs from the Project Described within the Poster—Where are All the Boars? An Attempt to Gain a Continental Perspective. Open Health Data 2016, 4, e1. [Google Scholar] [CrossRef]
- Jordt, A.M.; Lange, M.; Kramer-Schadt, S.; Nielsen, L.H.; Nielsen, S.S.; Thulke, H.H.; Vejre, H.; Alban, L. Spatio-temporal modeling of the invasive potential of wild boar—A conflict-prone species-using multi-source citizen science data. Prev. Vet. Med. 2016, 124, 34–44. [Google Scholar] [CrossRef]
- Croft, S.; Massei, G.; Smith, G.C.; Fouracre, D.; Aegerter, J.N. Modelling Spatial and Temporal Patterns of African Swine Fever in an Isolated Wild Boar Population to Support Decision-Making. Front. Vet. Sci. 2020, 7, 154. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.; Iglesias, I.; Munoz, M.J.; de la Torre, A. A Cartographic Tool for Managing African Swine Fever in Eurasia: Mapping Wild Boar Distribution Based on the Quality of Available Habitats. Transbound. Emerg. Dis. 2017, 64, 1720–1733. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, A.; Bosch, J.; Iglesias, I.; Munoz, M.J.; Mur, L.; Martinez-Lopez, B.; Martinez, M.; Sanchez-Vizcaino, J.M. Assessing the Risk of African Swine Fever Introduction into the European Union by Wild Boar. Transbound. Emerg. Dis. 2015, 62, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Thulke, H.H. Elucidating transmission parameters of African swine fever through wild boar carcasses by combining spatio-temporal notification data and agent-based modelling. Stoch. Environ. Res. Risk Assess. 2017, 31, 379–391. [Google Scholar] [CrossRef]
- Lange, M.; Guberti, V.; Thulke, H.H. Understanding ASF spread and emergency control concepts in wild boar populations using individual-based modelling and spatio-temporal surveillance data. External Scientific Report. EFSA Support. Publ. 2018, 15, 1521E. [Google Scholar] [CrossRef]
- Barongo, M.B.; Bishop, R.P.; Fevre, E.M.; Knobel, D.L.; Ssematimba, A. A Mathematical Model that Simulates Control Options for African Swine Fever Virus (ASFV). PLoS ONE 2016, 11, e0158658. [Google Scholar] [CrossRef] [PubMed]
- Guinat, C.; Vergne, T.; Jurado-Diaz, C.; Sanchez-Vizcaino, J.M.; Dixon, L.; Pfeiffer, D.U. Effectiveness and practicality of control strategies for African swine fever: What do we really know? Vet. Rec. 2017, 180, 97. [Google Scholar] [CrossRef]
- Gervasi, V.; Marcon, A.; Bellini, S.; Guberti, V. Evaluation of the Efficiency of Active and Passive Surveillance in the Detection of African Swine Fever in Wild Boar. Vet. Sci. 2019, 7, 5. [Google Scholar] [CrossRef]
- OIE. Chapter 3.8.1 African Swine Fever (Infection with African Swine Fever Virus). Available online: https://www.oie.int/app/uploads/2021/03/african-swine-fever.pdf (accessed on 19 August 2021).
- Gallardo, C.; Fernandez-Pinero, J.; Arias, M. African swine fever (ASF) diagnosis, an essential tool in the epidemiological investigation. Virus Res. 2019, 271, 197676. [Google Scholar] [CrossRef]
- Pikalo, J.; Deutschmann, P.; Fischer, M.; Roszyk, H.; Beer, M.; Blome, S. African Swine Fever Laboratory Diagnosis-Lessons Learned from Recent Animal Trials. Pathogens 2021, 10, 177. [Google Scholar] [CrossRef] [PubMed]
- Elnagar, A.; Pikalo, J.; Beer, M.; Blome, S.; Hoffmann, B. Swift and Reliable “Easy Lab” Methods for the Sensitive Molecular Detection of African Swine Fever Virus. Int. J. Mol. Sci. 2021, 22, 2307. [Google Scholar] [CrossRef]
- Carlson, J.; Zani, L.; Schwaiger, T.; Nurmoja, I.; Viltrop, A.; Vilem, A.; Beer, M.; Blome, S. Simplifying sampling for African swine fever surveillance: Assessment of antibody and pathogen detection from blood swabs. Transbound. Emerg. Dis. 2018, 65, e165–e172. [Google Scholar] [CrossRef]
- Petrov, A.; Schotte, U.; Pietschmann, J.; Drager, C.; Beer, M.; Anheyer-Behmenburg, H.; Goller, K.V.; Blome, S. Alternative sampling strategies for passive classical and African swine fever surveillance in wild boar. Vet. Microbiol. 2014. [Google Scholar] [CrossRef]
- Blome, S.; Goller, K.V.; Petrov, A.; Dräger, C.; Pietschmann, J.; Beer, M. Alternative sampling strategies for passive classical and African swine fever surveillance in wild boar–Extension towards African swine fever virus antibody detection. Vet. Microbiol. 2014, 174, 607–608. [Google Scholar] [CrossRef] [PubMed]
- Michaud, V.; Gil, P.; Kwiatek, O.; Prome, S.; Dixon, L.; Romero, L.; Le Potier, M.F.; Arias, M.; Couacy-Hymann, E.; Roger, F.; et al. Long-term storage at tropical temperature of dried-blood filter papers for detection and genotyping of RNA and DNA viruses by direct PCR. J. Virol. Methods 2007, 146, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Randriamparany, T.; Kouakou, K.V.; Michaud, V.; Fernandez-Pinero, J.; Gallardo, C.; Le Potier, M.F.; Rabenarivahiny, R.; Couacy-Hymann, E.; Raherimandimby, M.; Albina, E. African Swine Fever Diagnosis Adapted to Tropical Conditions by the Use of Dried-blood Filter Papers. Transbound. Emerg. Dis. 2016, 63, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Braae, U.C.; Johansen, M.V.; Ngowi, H.A.; Rasmussen, T.B.; Nielsen, J.; Uttenthal, A. Detection of African Swine Fever Virus DNA in Blood Samples Stored on FTA Cards from Asymptomatic Pigs in Mbeya Region, Tanzania. Transbound. Emerg. Dis. 2013, 62, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Flannery, J.; Ashby, M.; Moore, R.; Wells, S.; Rajko-Nenow, P.; Netherton, C.L.; Batten, C. Identification of novel testing matrices for African swine fever surveillance. J. Vet. Diagn. Investig. 2020, 32, 961–963. [Google Scholar] [CrossRef]
- McKillen, J.; McMenamy, M.; Hjertner, B.; McNeilly, F.; Uttenthal, A.; Gallardo, C.; Adair, B.; Allan, G. Sensitive detection of African swine fever virus using real-time PCR with a 5’ conjugated minor groove binder probe. J. Virol. Methods 2010, 168, 141–146. [Google Scholar] [CrossRef]
- Grau, F.R.; Schroeder, M.E.; Mulhern, E.L.; McIntosh, M.T.; Bounpheng, M.A. Detection of African swine fever, classical swine fever, and foot-and-mouth disease viruses in swine oral fluids by multiplex reverse transcription real-time polymerase chain reaction. J. Vet. Diagn. Investig. 2015, 27, 140–149. [Google Scholar] [CrossRef]
- Männistö, H.E. Collection of Oral Fluid Samples from Wild Boar in the Field Conditions to Detect African Swine Fever Virus (ASFV). Disseration. Estonian University of Life Sciences, Tartu, Estonia, 2018. Available online: https://dspace.emu.ee/xmlui/bitstream/handle/10492/4270/Hanna_Männistö_2018LA_VM.pdf?sequence=1&isAllowed=y (accessed on 19 August 2021).
- Nieto-Pelegrin, E.; Rivera-Arroyo, B.; Sanchez-Vizcaino, J.M. First Detection of Antibodies Against African Swine Fever Virus in Faeces Samples. Transbound. Emerg. Dis. 2015, 62, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.; Gallardo, C.; Soler, A.; Zimmermman, J.; Pelayo, V.; Nieto, R.; Sanchez-Vizcaino, J.M.; Arias, M. Potential use of oral fluid samples for serological diagnosis of African swine fever. Vet. Microbiol. 2013, 165, 135–139. [Google Scholar] [CrossRef]
- Liu, H.; Shi, K.; Sun, W.; Zhao, J.; Yin, Y.; Si, H.; Qu, S.; Lu, W. Development a multiplex RT-PCR assay for simultaneous detection of African swine fever virus, classical swine fever virus and atypical porcine pestivirus. J. Virol. Methods 2021, 287, 114006. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Noll, L.; Stoy, C.; Porter, E.; Fu, J.; Feng, Y.; Peddireddi, L.; Liu, X.; Dodd, K.A.; et al. Development of a real-time PCR assay for detection of African swine fever virus with an endogenous internal control. Transbound. Emerg. Dis. 2020, 67, 2446–2454. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Jia, R.; Liu, Y.; Zhou, J.; Qi, Y.; Chen, Y.; Liu, D.; Zhao, J.; Shi, H.; Zhang, J.; et al. Development of a novel quantitative real-time PCR assay with lyophilized powder reagent to detect African swine fever virus in blood samples of domestic pigs in China. Transbound. Emerg. Dis. 2020, 67, 284–297. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.T.; Dang, A.K.; Ly, D.V.; Vu, H.T.; Hoang, T.V.; Nguyen, C.T.; Chu, N.T.; Nguyen, V.T.; Nguyen, H.T.; Truong, A.D.; et al. An improvement of real-time polymerase chain reaction system based on probe modification is required for accurate detection of African swine fever virus in clinical samples in Vietnam. Asian-Australas. J. Anim. Sci. 2020, 33, 1683–1690. [Google Scholar] [CrossRef]
- Lin, Y.; Cao, C.; Shi, W.; Huang, C.; Zeng, S.; Sun, J.; Wu, J.; Hua, Q. Development of a triplex real-time PCR assay for detection and differentiation of gene-deleted and wild-type African swine fever virus. J. Virol. Methods 2020, 280, 113875. [Google Scholar] [CrossRef]
- Luo, Y.; Atim, S.A.; Shao, L.; Ayebazibwe, C.; Sun, Y.; Liu, Y.; Ji, S.; Meng, X.Y.; Li, S.; Li, Y.; et al. Development of an updated PCR assay for detection of African swine fever virus. Arch. Virol. 2017, 162, 191–199. [Google Scholar] [CrossRef]
- Liu, L.; Luo, Y.; Accensi, F.; Ganges, L.; Rodríguez, F.; Shan, H.; Ståhl, K.; Qiu, H.J.; Belák, S. Pre-Clinical Evaluation of a Real-Time PCR Assay on a Portable Instrument as a Possible Field Diagnostic Tool: Experiences from the Testing of Clinical Samples for African and Classical Swine Fever Viruses. Transbound. Emerg. Dis. 2017, 64, e31–e35. [Google Scholar] [CrossRef]
- Fernández-Pinero, J.; Gallardo, C.; Elizalde, M.; Robles, A.; Gómez, C.; Bishop, R.; Heath, L.; Couacy-Hymann, E.; Fasina, F.O.; Pelayo, V.; et al. Molecular diagnosis of African Swine Fever by a new real-time PCR using universal probe library. Transbound. Emerg. Dis. 2013, 60, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Tignon, M.; Gallardo, C.; Iscaro, C.; Hutet, E.; Van der Stede, Y.; Kolbasov, D.; De Mia, G.M.; Le Potier, M.F.; Bishop, R.P.; Arias, M.; et al. Development and inter-laboratory validation study of an improved new real-time PCR assay with internal control for detection and laboratory diagnosis of African swine fever virus. J. Virol. Methods 2011, 178, 161–170. [Google Scholar] [CrossRef]
- Ronish, B.; Hakhverdyan, M.; Ståhl, K.; Gallardo, C.; Fernandez-Pinero, J.; Belák, S.; Leblanc, N.; Wangh, L. Design and verification of a highly reliable Linear-After-The-Exponential PCR (LATE-PCR) assay for the detection of African swine fever virus. J. Virol. Methods 2011, 172, 8–15. [Google Scholar] [CrossRef]
- Zsak, L.; Borca, M.V.; Risatti, G.R.; Zsak, A.; French, R.A.; Lu, Z.; Kutish, G.F.; Neilan, J.G.; Callahan, J.D.; Nelson, W.M.; et al. Preclinical diagnosis of African swine fever in contact-exposed swine by a real-time PCR assay. J. Clin. Microbiol. 2005, 43, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Agüero, M.; Fernández, J.; Romero, L.J.; Zamora, M.J.; Sánchez, C.; Belák, S.; Arias, M.; Sánchez-Vizcaíno, J.M. A highly sensitive and specific gel-based multiplex RT-PCR assay for the simultaneous and differential diagnosis of African swine fever and Classical swine fever in clinical samples. Vet. Res. 2004, 35, 551–563. [Google Scholar] [CrossRef]
- King, D.P.; Reid, S.M.; Hutchings, G.H.; Grierson, S.S.; Wilkinson, P.J.; Dixon, L.K.; Bastos, A.D.; Drew, T.W. Development of a TaqMan PCR assay with internal amplification control for the detection of African swine fever virus. J. Virol. Methods 2003, 107, 53–61. [Google Scholar] [CrossRef]
- Blome, S.; Franzke, K.; Beer, M. African swine fever—A review of current knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef] [PubMed]
- Schoder, M.E.; Tignon, M.; Linden, A.; Vervaeke, M.; Cay, A.B. Evaluation of seven commercial African swine fever virus detection kits and three Taq polymerases on 300 well-characterized field samples. J. Virol. Methods 2020, 280, 113874. [Google Scholar] [CrossRef]
- Fraczyk, M.; Wozniakowski, G.; Kowalczyk, A.; Niemczuk, K.; Pejsak, Z. Development of cross-priming amplification for direct detection of the African Swine Fever Virus, in pig and wild boar blood and sera samples. Lett. Appl. Microbiol. 2016, 62, 386–391. [Google Scholar] [CrossRef]
- James, H.E.; Ebert, K.; McGonigle, R.; Reid, S.M.; Boonham, N.; Tomlinson, J.A.; Hutchings, G.H.; Denyer, M.; Oura, C.A.; Dukes, J.P.; et al. Detection of African swine fever virus by loop-mediated isothermal amplification. J. Virol. Methods 2010, 164, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, A.L.; Bustos, M.J.; de Leon, P. Methods for growing and titrating African swine fever virus: Field and laboratory samples. Curr. Protoc. Cell Biol. 2011, 53, 14–26. [Google Scholar] [CrossRef]
- Malmquist, W.A.; Hay, D. Hemadsorption and cytopathic effect produced by African Swine Fever virus in swine bone marrow and buffy coat cultures. Am. J. Vet. Res. 1960, 21, 104–108. [Google Scholar] [PubMed]
- Gallardo, C.; Nieto, R.; Soler, A.; Pelayo, V.; Fernandez-Pinero, J.; Markowska-Daniel, I.; Pridotkas, G.; Nurmoja, I.; Granta, R.; Simon, A.; et al. Assessment of African Swine Fever Diagnostic Techniques as a Response to the Epidemic Outbreaks in Eastern European Union Countries: How To Improve Surveillance and Control Programs. J. Clin. Microbiol. 2015, 53, 2555–2565. [Google Scholar] [CrossRef] [PubMed]
- Sastre, P.; Gallardo, C.; Monedero, A.; Ruiz, T.; Arias, M.; Sanz, A.; Rueda, P. Development of a novel lateral flow assay for detection of African swine fever in blood. BMC Vet. Res. 2016, 12, 206. [Google Scholar] [CrossRef]
- Boklund, A.; Dhollander, S.; Chesnoiu Vasile, T.; Abrahantes, J.C.; Botner, A.; Gogin, A.; Gonzalez Villeta, L.C.; Gortazar, C.; More, S.J.; Papanikolaou, A.; et al. Risk factors for African swine fever incursion in Romanian domestic farms during 2019. Sci. Rep. 2020, 10, 10215. [Google Scholar] [CrossRef] [PubMed]
- Nurmoja, I.; Mõtus, K.; Kristian, M.; Niine, T.; Schulz, K.; Depner, K.; Viltrop, A. Epidemiological analysis of the 2015–2017 African swine fever outbreaks in Estonia. Prev. Vet. Med. 2018. [Google Scholar] [CrossRef]
- Jori, F.; Chenais, E.; Boinas, F.; Busauskas, P.; Dholllander, S.; Fleischmann, L.; Olsevskis, E.; Rijks, J.M.; Schulz, K.; Thulke, H.H.; et al. Application of the World Cafe method to discuss the efficiency of African swine fever control strategies in European wild boar (Sus scrofa) populations. Prev. Vet. Med. 2020, 185, 105178. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.K.; Stahl, K.; Jori, F.; Vial, L.; Pfeiffer, D.U. African Swine Fever Epidemiology and Control. Annu. Rev. Anim. Biosci. 2020, 8, 221–246. [Google Scholar] [CrossRef] [PubMed]
- Mebus, C.A.; House, C.; Ruiz Gonzalvo, F.; Pineda, J.M.; Tapiador, J.; Pire, J.J.; Bergada, J.; Yedloutschnig, R.J.; Sahu, S.; Becerra, V.; et al. Survival of foot-and-mouth disesase, African swine fever, and hog cholera viruses in Spanish serrano cured hams and Iberian cured hams, shoulders and loins. Food Microbiol. 1993, 10, 133–143. [Google Scholar] [CrossRef]
- Saegerman, C. Découverte inattendue de la peste porcine africaine en Belgique. Épidémiol. St. Anim. 2018, 73, 147–164. [Google Scholar]
- Danzetta, M.L.; Marenzoni, M.L.; Iannetti, S.; Tizzani, P.; Calistri, P.; Feliziani, F. African Swine Fever: Lessons to Learn From Past Eradication Experiences. A Systematic Review. Front. Vet. Sci. 2020, 7, 296. [Google Scholar] [CrossRef]
- EFSA. African swine fever in wild boar. EFSA J. 2018, 16, 5344. [Google Scholar] [CrossRef]
- Jori, F.; Relun, A.; Trabucco, B.; Charrier, F.; Maestrini, O.; Chavernac, D.; Cornelis, D.; Casabianca, F.; Etter, E.M.C. Questionnaire-Based Assessment of Wild Boar/Domestic Pig Interactions and Implications for Disease Risk Management in Corsica. Front. Vet. Sci. 2017, 4, 198. [Google Scholar] [CrossRef] [PubMed]
- Bellini, S.; Rutili, D.; Guberti, V. Preventive measures aimed at minimizing the risk of African swine fever virus spread in pig farming systems. Acta Vet. Scand. 2016, 58, 82. [Google Scholar] [CrossRef] [PubMed]
- OIE. African Swine Fever. Available online: https://www.oie.int/en/disease/african-swine-fever/ (accessed on 4 June 2021).
- Urner, N.; Seržants, M.; Užule, M.; Sauter-Louis, C.; Staubach, C.; Lamberga, K.; Oļševskis, E.; Conraths, F.J.; Schulz, K. Hunters’ view on the control of African swine fever in wild boar. A participatory study in Latvia. Prev. Vet. Med. 2021, 186, 105229. [Google Scholar] [CrossRef]
- Stonciute, E.; Schulz, K.; Malakauskas, A.; Conraths, F.J.; Masiulis, M.; Sauter-Louis, C. What Do Lithuanian Hunters Think of African Swine Fever and Its Control-Perceptions. Animals 2021, 11, 525. [Google Scholar] [CrossRef]
- Urner, N.; Mõtus, K.; Nurmoja, I.; Schulz, J.; Sauter-Louis, C.; Staubach, C.; Conraths, F.J.; Schulz, K. Hunters’ Acceptance of Measures against African Swine Fever in Wild Boar in Estonia. Prev. Vet. Med. 2020, 182, 105121. [Google Scholar] [CrossRef] [PubMed]
- Urner, N.; Sauter-Louis, C.; Staubach, C.; Conraths, F.J.; Schulz, K. A Comparison of Perceptions of Estonian and Latvian Hunters With Regard to the Control of African Swine Fever. Front. Vet. Sci. 2021, 8, 642126. [Google Scholar] [CrossRef] [PubMed]
- Malakauskas, A.; Schulz, K.; Kukanauskaité, I.; Masiulis, M.; Conraths, F.J.; Sauter-Louis, C. Why me? African swine fever outbreaks in Lithuanian domestic pigs in 2019. Animals 2021, submitted. [Google Scholar]



| Country | Region | First Occurrence of ASF in Wild Boar | Presumed or Proven Route of Introduction | Reference | Presumed Main Driver for ASF Epidemic in Wild Boar |
|---|---|---|---|---|---|
| Portugal | n.d. | Costard et al., 2009 [50] | Interaction of domestic and free-ranging pigs/Ornithodoros ticks | ||
| Spain | n.d. | Spread from Portugal | |||
| Italy | Sardinia | n.d. | Pork products from Portugal | Mannelli et al., 1998 [51] | Interaction of domestic and free-ranging pigs |
| Georgia | n.d. | Waste from ships | Vepkhvadze et al., 2017 [52] | n.d. | |
| Armenia | North | Oct 2010 | Movement of infected pigs and wild boar from Georgia | FAO 2008 [53]; Markosyan et al., 2019 [54] | Interaction of free-ranging domestic pigs and wild pigs |
| Azerbaijan | n.d. | Pork products from Georgia | FAO, 2008 [53] | n.d. | |
| Russian Federation | Northern Caucasus | Nov 2007 | Wild boar movements from Georgia | Gogin et al., 2013 [55]; FAO, 2008 [53] | Wild boar/free-ranging pigs |
| Belarus | n.d. | n.d. | GF-TADs, 2015 [56] | n.d. | |
| Ukraine | Lugansk (2014) | Jan 2014 | Wild boar movements from Russian Federation, 2014 | DEFRA, 2014 [57] | Interaction of domestic and free-ranging pigs or backyard holdings |
| Lithuania | Jan 2014 | Wild boar movements from Belarus | State Food and Veterinary Service Lithuania, SCoPAFF, Feb. 2014 [58] | Wild boar | |
| Poland | East | Feb 2014 | Wild boar movements from Belarus | Wozniakowski et al., 2016 [8] | Wild boar |
| Warsaw | Nov 2017 | Human activity | General Vet. Inspectorate, Poand, SCoPAFF, Jan 2018 [59] | Wild boar | |
| North | Dec 2017 | Wild boar movements from Kaliningrad, RF | n.d. | Wild boar | |
| West | Nov 2019 | Human activity (?) | Mazur-Panasiuk et al., 2020 [60] | Wild boar | |
| Latvia | East | June 2014 | Wild boar movements from Belarus | Olsevskis et al., 2016 [61] | Wild boar |
| North | July 2014 | Illegal disposal of contaminated material | |||
| Madona | Aug 2014 | Human activity (?) | |||
| Estonia | South | Sep 2014 | Wild boar movements from Latvia | Nurmoja et al., 2017 [4] | Wild boar |
| North | Sep 2014 | Wild boar movements from RF | |||
| Moldova | May 2019 | GF TADs, 2016 [62] | Outbreak in domestic pigs or wild boar movements | ||
| Czech Republic | Zlín | June 2017 | Illegal disposal of food waste | OIE, 2019 [20] | Wild boar |
| Hungary | April 2018 | Illegal disposal of food waste | EFSA, 2020 [63] | n.d. | |
| Romania | Satu Mare | May 2018 | Human activity | EFSA, 2020 [63] | Outbreaks in domestic pigs and pig holding structure |
| Danube Delta | June 2018 | ||||
| Bulgaria | August 2018 | Human activity, wild boar movement (from Romania?) | Zani et al., 2019 [64] | Outbreak in domestic pigs or wild boar movements | |
| Belgium | Wallonia | Sep 2018 | Illegal disposal of food waste | Linden, 2019 [65] | Wild boar |
| Slovak Republic | Aug 2019 | Wild boar movements from Hungary | EFSA, 2020 [63] | n.d. | |
| Serbia | July 2019 | Wild boar movements from Romania / Bulgaria | Reuters, 2020 [66] | n.d. | |
| Germany | Brandenburg and Saxony (Polish Border) | Sept 2020 | Wild boar movements from Poland | Sauter-Louis, et al., 2020 [6] | Wild boar |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sauter-Louis, C.; Conraths, F.J.; Probst, C.; Blohm, U.; Schulz, K.; Sehl, J.; Fischer, M.; Forth, J.H.; Zani, L.; Depner, K.; et al. African Swine Fever in Wild Boar in Europe—A Review. Viruses 2021, 13, 1717. https://doi.org/10.3390/v13091717
Sauter-Louis C, Conraths FJ, Probst C, Blohm U, Schulz K, Sehl J, Fischer M, Forth JH, Zani L, Depner K, et al. African Swine Fever in Wild Boar in Europe—A Review. Viruses. 2021; 13(9):1717. https://doi.org/10.3390/v13091717
Chicago/Turabian StyleSauter-Louis, Carola, Franz J. Conraths, Carolina Probst, Ulrike Blohm, Katja Schulz, Julia Sehl, Melina Fischer, Jan Hendrik Forth, Laura Zani, Klaus Depner, and et al. 2021. "African Swine Fever in Wild Boar in Europe—A Review" Viruses 13, no. 9: 1717. https://doi.org/10.3390/v13091717
APA StyleSauter-Louis, C., Conraths, F. J., Probst, C., Blohm, U., Schulz, K., Sehl, J., Fischer, M., Forth, J. H., Zani, L., Depner, K., Mettenleiter, T. C., Beer, M., & Blome, S. (2021). African Swine Fever in Wild Boar in Europe—A Review. Viruses, 13(9), 1717. https://doi.org/10.3390/v13091717