
Exacerbation of Influenza A Virus Disease Severity by Respiratory Syncytial Virus Co-Infection in a Mouse Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Animals and Virus Inoculation
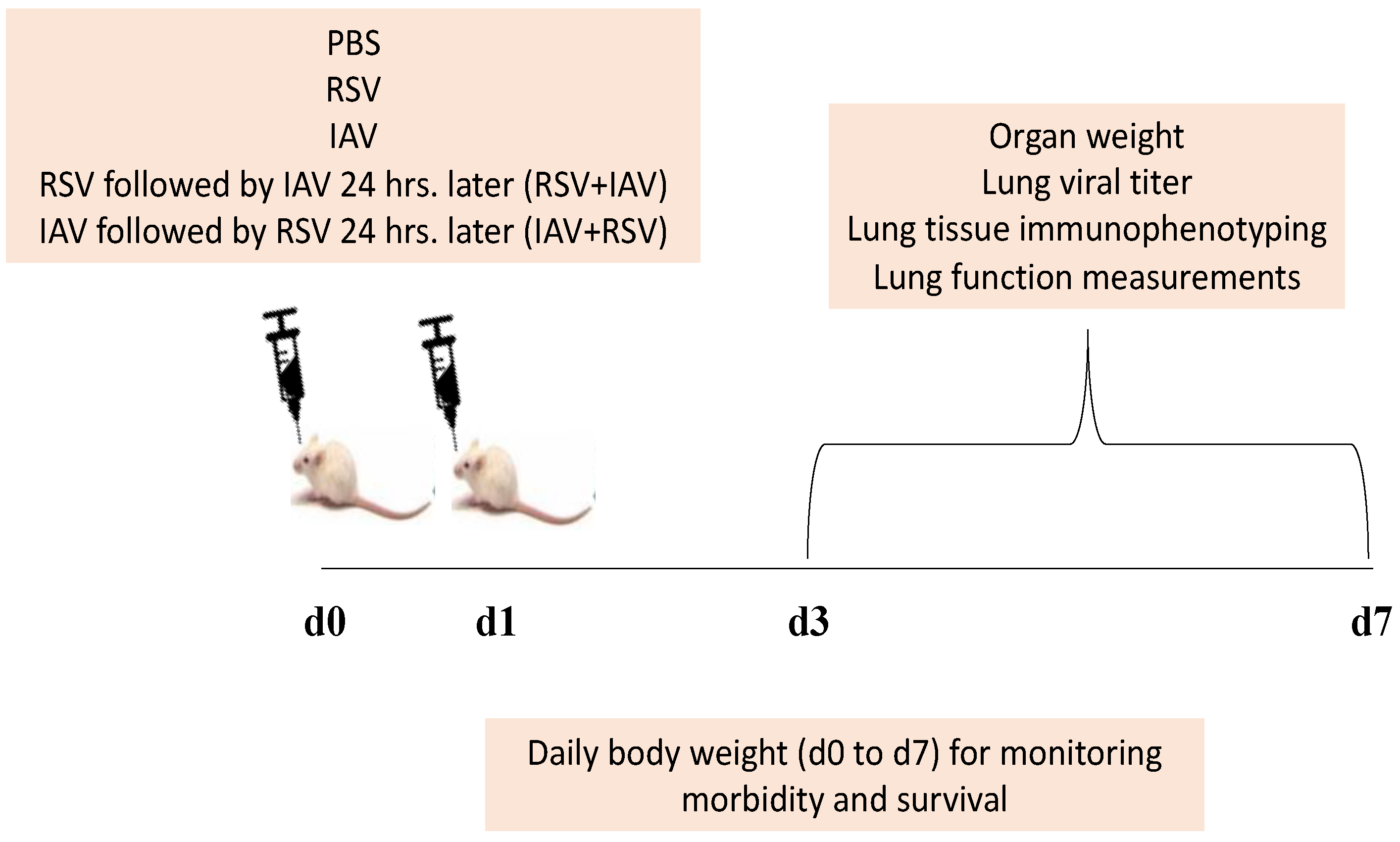
2.3. Experimental Strategy
2.4. Sample Processing
2.5. Quantitative PCR for Determining Viral Loads
2.6. Flow Cytometry by Fluorescence Activated Cell Sorting (FACS)
2.7. Lung Function Assessment
2.8. Statistical Analysis
3. Results
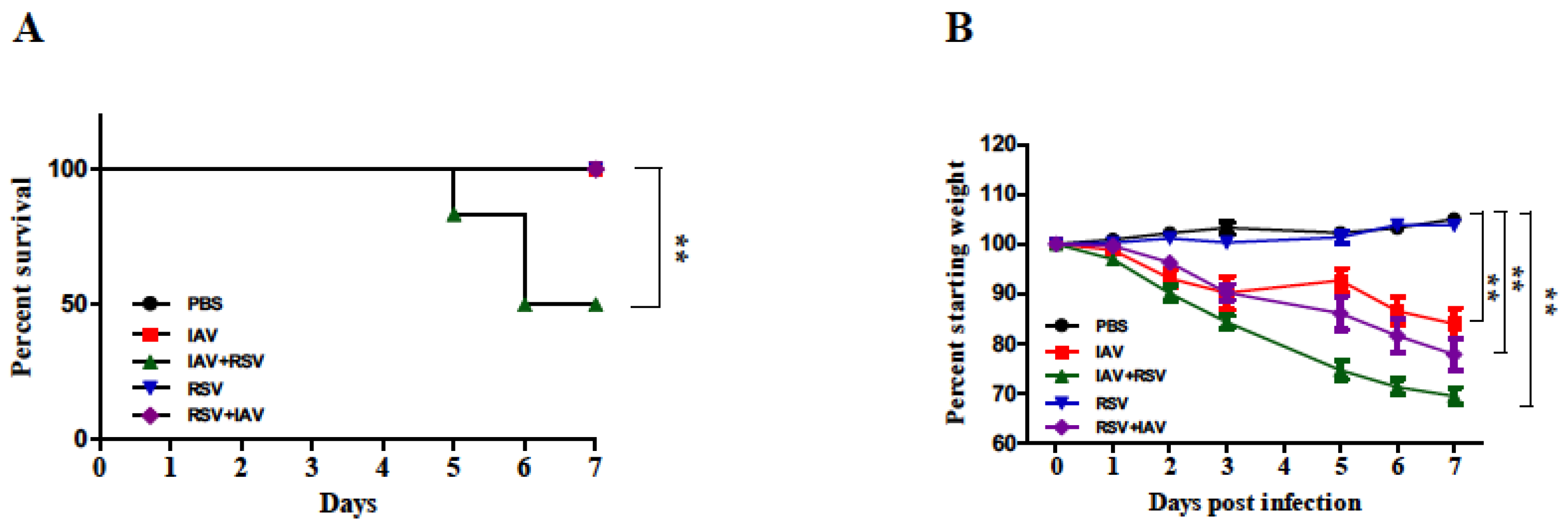
3.1. Body Weight Loss Was More Severe in Mice Infected with IAV 24 h Prior to RSV Inoculation
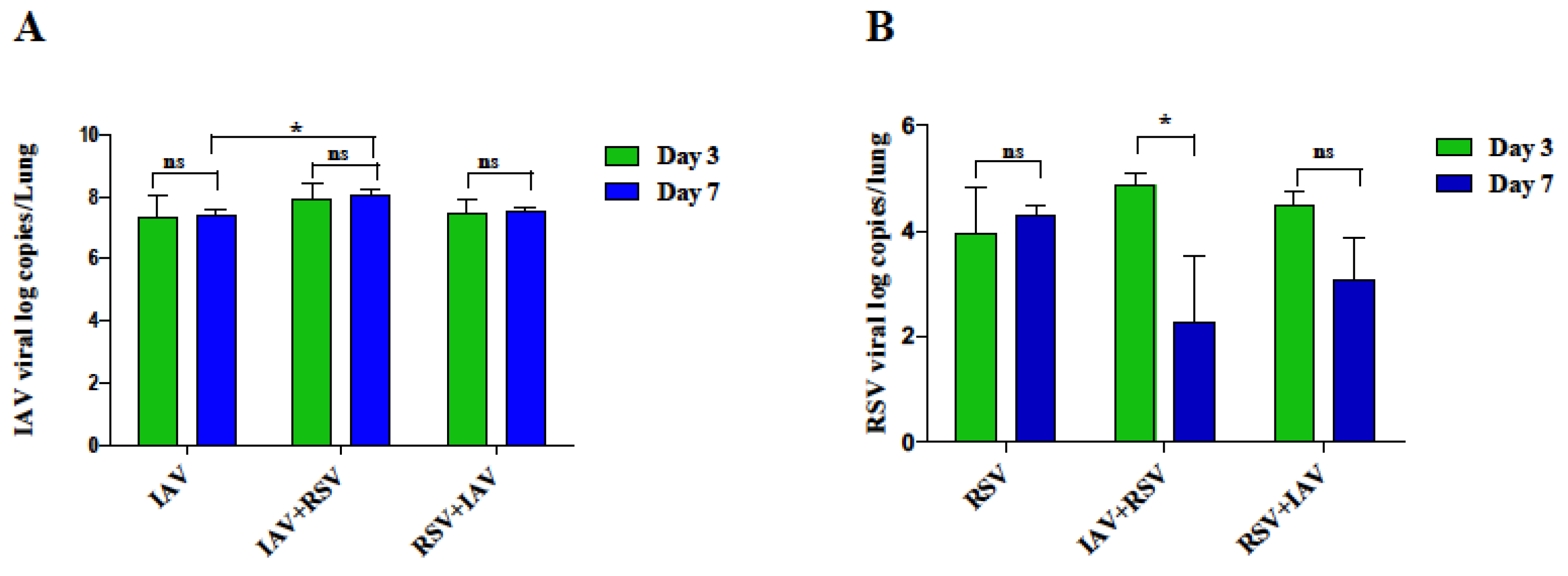
3.2. RSV Infection following IAV Results in Remarkable Increase in IAV Viral Load in the Lungs on Day 7 Post-Infection
3.3. IAV Infected Mice Exhibit Reduced RSV Viral Load following Subsequent RSV Infection on Day 7 Post-Infection
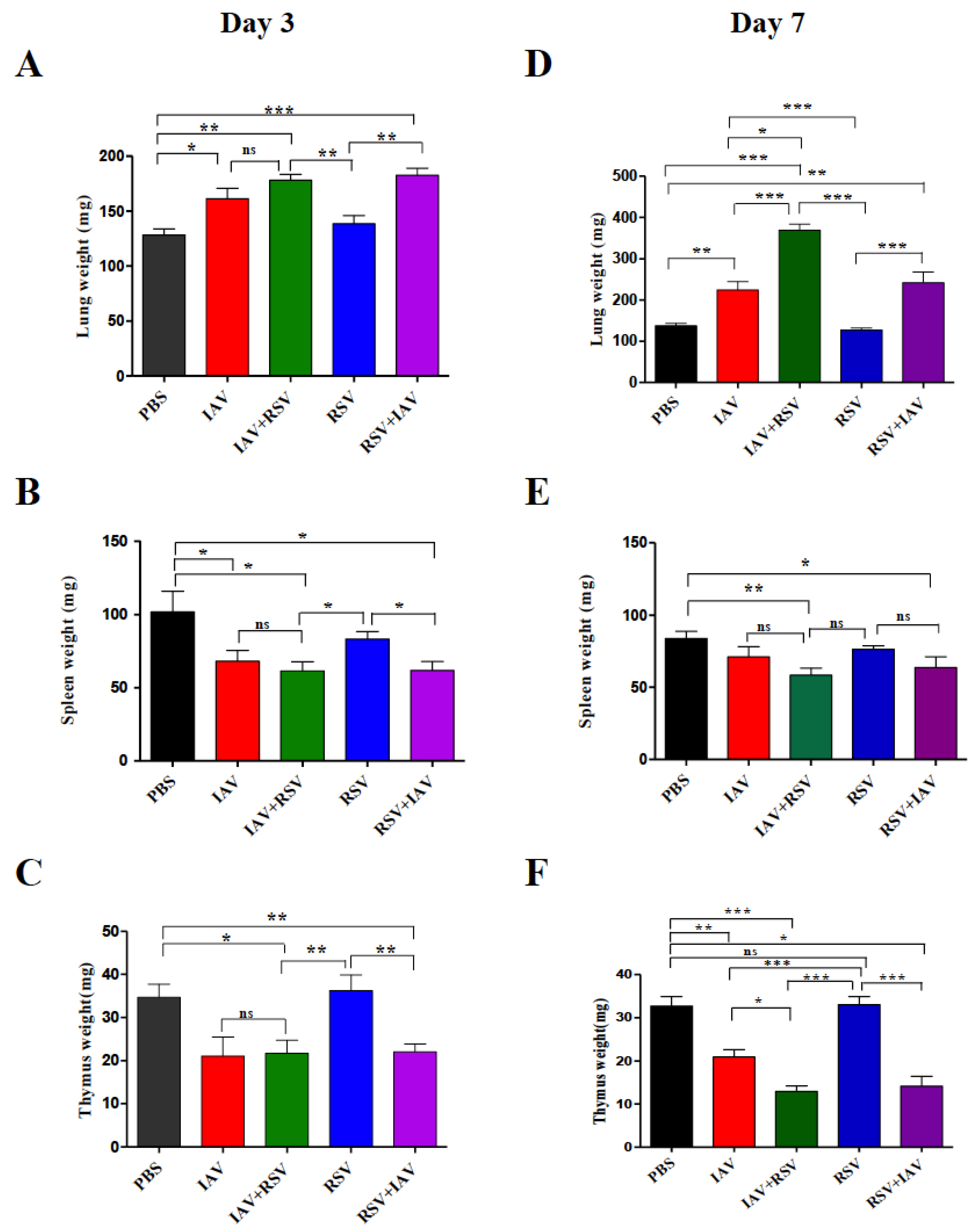
3.4. Co-Infection of IAV with RSV Leads to Significant Weight Loss of Immune System Organs and Significant Enlargement of Lungs
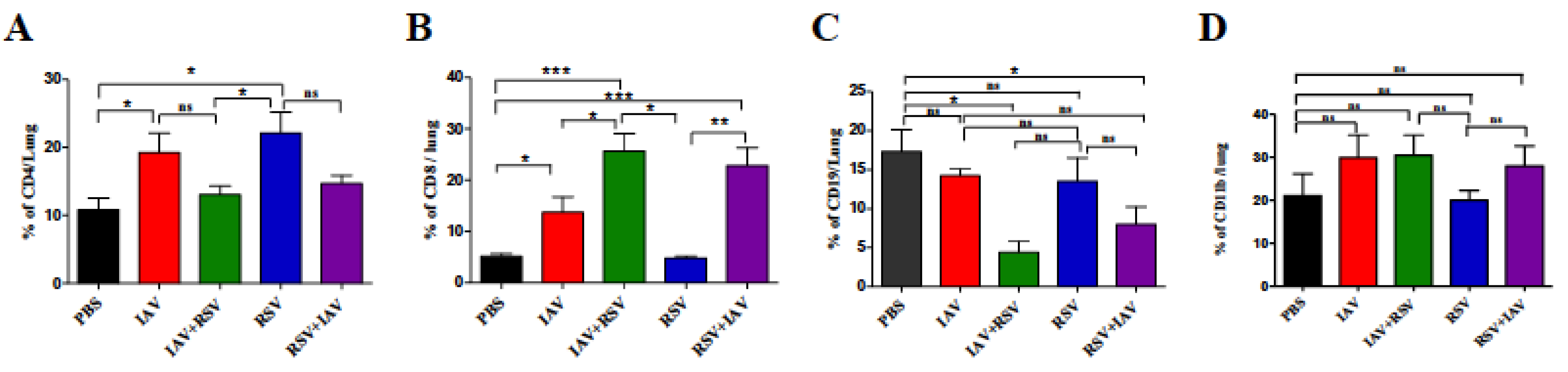
3.5. Co-Infection Resulted in a High Number of CD8 T Cells and Low Numbers of CD4 T Cells and B Cells
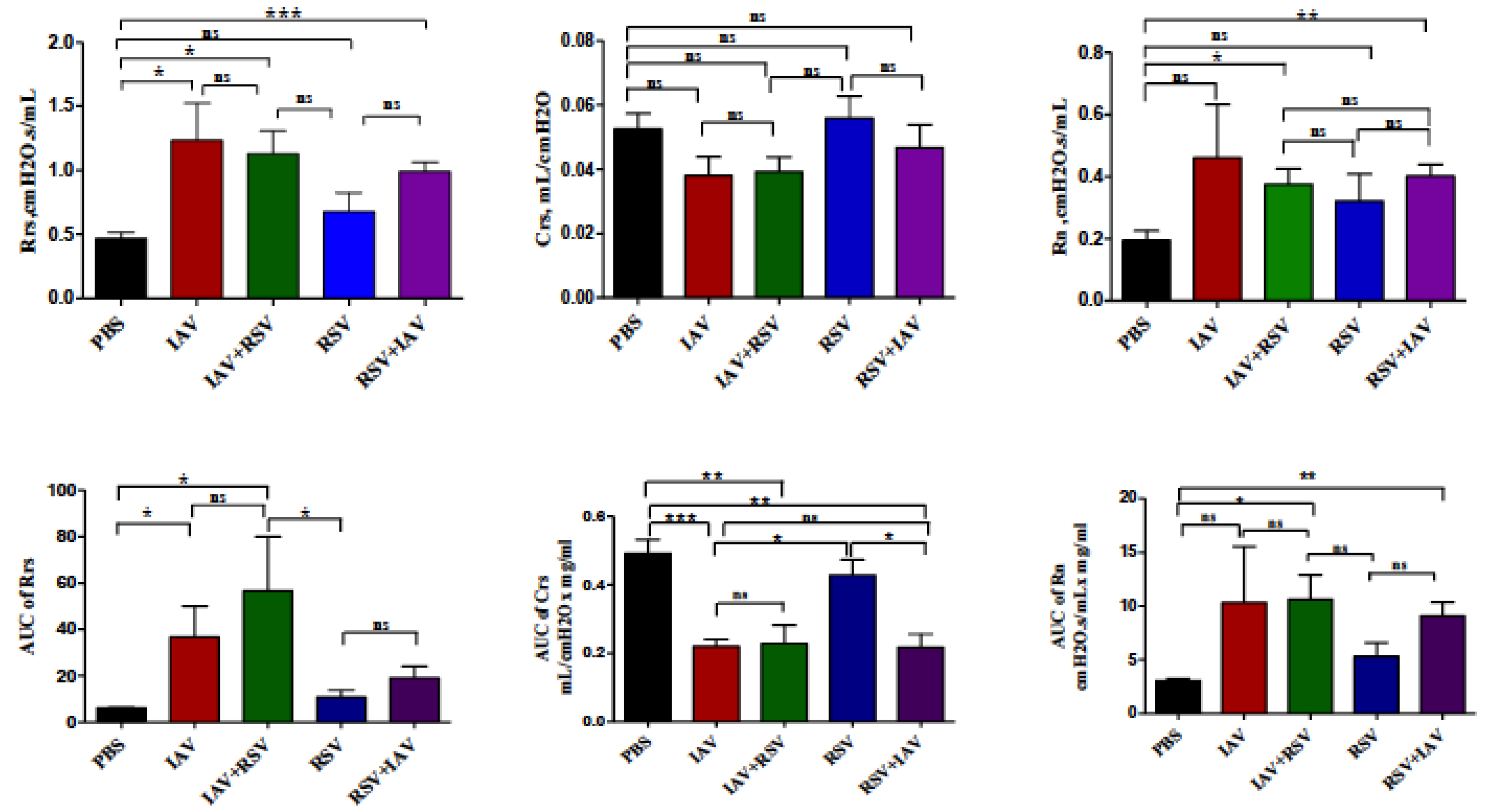
3.6. Co-Infection with IAV+RSV or RSV+IAV Leads to Increased Thoracic Resistance and Large Airway Resistance
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2016 Lower Respiratory Infections Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1191–1210. [Google Scholar] [CrossRef] [Green Version]
- Aberle, J.H.; Aberle, S.W.; Pracher, E.; Hutter, H.P.; Kundi, M.; Popow-Kraupp, T. Single versus dual respiratory virus infections in hospitalized infants: Impact on clinical course of disease and interferon-γ response. Pediatric Infect. Dis. J. 2005, 24, 605–610. [Google Scholar] [CrossRef]
- Lee, H.J.; Seo, Y.E.; Han, S.B.; Jeong, D.C.; Kang, J.H. Clinical Impact of Mixed Respiratory Viral Infection in Children with Adenoviral Infection. Infect. Chemother. 2016, 48, 309–316. [Google Scholar] [CrossRef]
- DaPalma, T.; Doonan, B.P.; Trager, N.M.; Kasman, L.M. A systematic approach to virus–virus interactions. Virus Res. 2010, 149, 1–9. [Google Scholar] [CrossRef]
- Opatowski, L.; Baguelin, M.; Eggo, R.M. Influenza interaction with cocirculating pathogens and its impact on surveillance, pathogenesis, and epidemic profile: A key role for mathematical modelling. PLoS Pathog. 2018, 14, e1006770. [Google Scholar] [CrossRef] [Green Version]
- WHO|Influenza. Available online: https://www.who.int/en/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 28 June 2021).
- Shi, T.; McAllister, D.A.; O’Brien, K.L.; Simoes, E.A.F.; Madhi, S.A.; Gessner, B.D.; Polack, F.P.; Balsells, E.; Acacio, S.; Aguayo, C.; et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: A systematic review and modelling study. Lancet 2017, 390, 946–958. [Google Scholar] [CrossRef] [Green Version]
- Troeger, C.E.; Blacker, B.F.; Khalil, I.A.; Zimsen, S.R.M.; Albertson, S.B.; Abate, D.; Abdela, J.; Adhikari, T.B.; Aghayan, S.A.; Agrawal, S.; et al. Mortality, morbidity, and hospitalisations due to influenza lower respiratory tract infections, 2017: An analysis for the Global Burden of Disease Study 2017. Lancet Respir. Med. 2019, 7, 69–89. [Google Scholar] [CrossRef] [Green Version]
- Sanders, C.J.; Doherty, P.C.; Thomas, P.G. Respiratory epithelial cells in innate immunity to influenza virus infection. Cell Tissue Res. 2011, 343, 13–21. [Google Scholar] [CrossRef]
- Lee, K.H.; Gordon, A.; Foxman, B. The role of respiratory viruses in the etiology of bacterial pneumonia. Evol. Med. Public Health 2016, 2016, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Shinjoh, M.; Omoe, K.; Saito, N.; Matsuo, N.; Nerome, K. In vitro growth profiles of respiratory syncytial virus in the presence of influenza virus. Acta Virol. 2000, 44, 91–97. [Google Scholar] [PubMed]
- Price, O.H.; Sullivan, S.G.; Sutterby, C.; Druce, J.; Carville, K.S. Using routine testing data to understand circulation patterns of influenza A, respiratory syncytial virus and other respiratory viruses in Victoria, Australia. Epidemiol. Infect. 2019, 147, e221. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Sharma, S.; Barua, S.; Tripathi, B.N.; Rouse, B.T. Virological and immunological outcomes of coinfections. Clin. Microbiol. Rev. 2018, 31. [Google Scholar] [CrossRef] [Green Version]
- Alsuwaidi, A.R.; George, J.A.; Almarzooqi, S.; Hartwig, S.M.; Varga, S.M.; Souid, A.-K.K. Sirolimus alters lung pathology and viral load following influenza A virus infection. Respir. Res. 2017, 18, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsuwaidi, A.R.; Almarzooqi, S.; Albawardi, A.; Benedict, S.; Kochiyil, J.; Mustafa, F.; Hartwig, S.M.; Varga, S.M.; Souid, A.-K. Cellular bioenergetics, caspase activity and glutathione in murine lungs infected with influenza A virus. Virology 2013, 446, 180–188. [Google Scholar] [CrossRef]
- Hu, A.; Colella, M.; Tam, J.S.; Rappaport, R.; Cheng, S.M. Simultaneous detection, subgrouping, and quantitation of respiratory syncytial virus A and B by real-time PCR. J. Clin. Microbiol. 2003, 41, 149–154. [Google Scholar] [CrossRef] [Green Version]
- George, J.A.; Bashir, G.; Qureshi, M.M.; Mohamed, Y.A.; Azzi, J.; Al-Ramadi, B.K.; Fernández-Cabezudo, M.J. Cholinergic Stimulation Prevents the Development of Autoimmune Diabetes: Evidence for the Modulation of Th17 Effector Cells via an IFN$γ$-Dependent Mechanism. Front. Immunol. 2016, 7, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.J.; Ijezie, E.C.; Balemba, O.B.; Miura, T.A. Attenuation of Influenza A Virus Disease Severity by Viral Coinfection in a Mouse Model. J. Virol. 2018, 92, e00881-18. [Google Scholar] [CrossRef] [Green Version]
- Ayegbusi, O.T.; Ajagbe, O.A.; Afowowe, T.O.; Aransi, A.T.; Olusola, B.A.; Awogbindin, I.O.; Ogunsemowo, O.O.; Faneye, A.O.; Odaibo, G.N.; Olaleye, D.O. Virus genes and host correlates of pathology are markedly reduced during respiratory syncytial and influenza virus co-infection in BALB/c mice. Heliyon 2019, 5, e01094. [Google Scholar] [CrossRef] [Green Version]
- Goto, H.; Ihira, H.; Morishita, K.; Tsuchiya, M.; Ohta, K.; Yumine, N.; Tsurudome, M.; Nishio, M. Enhanced growth of influenza A virus by coinfection with human parainfluenza virus type 2. Med. Microbiol. Immunol. 2016, 205, 209–218. [Google Scholar] [CrossRef]
- Pletneva, L.M.; Haller, O.; Porter, D.D.; Prince, G.A.; Blanco, J.C.G. Induction of type I interferons and interferon-inducible Mx genes during respiratory syncytial virus infection and reinfection in cotton rats. J. Gen. Virol. 2008, 89, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.F.; Carolan, L.A.; Korenkov, D.; Druce, J.; Mccaw, J.; Reading, P.C.; Barr, I.G.; Laurie, K.L. Investigating viral interference between influenza a virus and human respiratory syncytial virus in a ferret model of infection. J. Infect. Dis. 2018, 218, 406–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drori, Y.; Jacob-Hirsch, J.; Pando, R.; Glatman-Freedman, A.; Friedman, N.; Mendelson, E.; Mandelboim, M. Influenza A Virus Inhibits RSV Infection via a Two-Wave Expression of IFIT Proteins. Viruses 2020, 12, 1171. [Google Scholar] [CrossRef] [PubMed]
- Herold, S.; Becker, C.; Ridge, K.M.; Budinger, G.R.S. Influenza virus-induced lung injury: Pathogenesis and implications for treatment. Eur. Respir. J. 2015, 45, 1463–1478. [Google Scholar] [CrossRef] [Green Version]
- Rossey, I.; Sedeyn, K.; De Baets, S.; Schepens, B.; Saelens, X. CD8+ T cell immunity against human respiratory syncytial virus. Vaccine 2014, 32, 6130–6137. [Google Scholar] [CrossRef]
- Thimme, R.; Wieland, S.; Steiger, C.; Ghrayeb, J.; Reimann, K.A.; Purcell, R.H.; Chisari, F.V. CD8+ T Cells Mediate Viral Clearance and Disease Pathogenesis during Acute Hepatitis B Virus Infection. J. Virol. 2003, 77, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.E.; Varga, S.M. The CD8 T cell response to respiratory virus infections. Front. Immunol. 2018, 9, 678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiege, J.K.; Stone, I.A.; Dumm, R.E.; Waring, B.M.; Fife, B.T.; Agudo, J.; Brown, B.D.; Heaton, N.S.; Langlois, R.A. Long-term surviving influenza infected cells evade CD8+ T cell mediated clearance. PLoS Pathog. 2019, 15, e1008077. [Google Scholar] [CrossRef] [Green Version]
- Jafri, H.S.; Chavez-Bueno, S.; Mejias, A.; Gomez, A.M.; Rios, A.M.; Nassi, S.S.; Yusuf, M.; Kapur, P.; Hardy, R.D.; Hatfield, J.; et al. Respiratory syncytial virus induces pneumonia, cytokine response, airway obstruction, and chronic inflammatory infiltrates associated with long-term airway hyperresponsiveness in mice. J. Infect. Dis. 2004, 189, 1856–1865. [Google Scholar] [CrossRef] [Green Version]
- Campbell, M.; Sapra, A. Physiology, Airflow Resistance. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Bem, R.A.; Domachowske, J.B.; Rosenberg, H.F. Animal models of human respiratory syncytial virus disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 301, L148–L156. [Google Scholar] [CrossRef] [Green Version]
- Bouvier, N.M.; Lowen, A.C. Animal Models for Influenza Virus Pathogenesis and Transmission. Viruses 2010, 2, 1530–1563. [Google Scholar] [CrossRef] [Green Version]
- Alsuwaidi, A.R.; Albawardi, A.; Almarzooqi, S.; Benedict, S.; Othman, A.R.; Hartwig, S.M.; Varga, S.M.; Souid, A.-K. Respiratory syncytial virus increases lung cellular bioenergetics in neonatal C57BL/6 mice. Virology 2014, 454, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsuwaidi, A.R.; Benedict, S.; Kochiyil, J.; Mustafa, F.; Hartwig, S.M.; Almarzooqi, S.; Albawardi, A.; Rizvi, T.A.; Varga, S.M.; Souid, A.-K. Bioenergetics of murine lungs infected with respiratory syncytial virus. Virol. J. 2013, 10, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| RSV N Forward | 5′GCTCTTAGCAAAGTCAAGTTGAATGA3′ |
| RSV N Reverse | 5′TGCTCCGTTGGATGGTGTATT-3′ |
| RSV N Probe | FAM/ACACTCAACAAAGATCAACTTCTGTCATCCAGC TAMRA |
| IAV Forward | AGA TGA GTC TTC TAA CCG AGG TCG |
| IAV Reverse | TGC AAA AAC ATC TTC AAG TCT CTG |
| IAV Probe | FAM-TCA GGC CCC CTC AAA GCC GA-TAMRA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, J.A.; AlShamsi, S.H.; Alhammadi, M.H.; Alsuwaidi, A.R. Exacerbation of Influenza A Virus Disease Severity by Respiratory Syncytial Virus Co-Infection in a Mouse Model. Viruses 2021, 13, 1630. https://doi.org/10.3390/v13081630
George JA, AlShamsi SH, Alhammadi MH, Alsuwaidi AR. Exacerbation of Influenza A Virus Disease Severity by Respiratory Syncytial Virus Co-Infection in a Mouse Model. Viruses. 2021; 13(8):1630. https://doi.org/10.3390/v13081630
Chicago/Turabian StyleGeorge, Junu A., Shaikha H. AlShamsi, Maryam H. Alhammadi, and Ahmed R. Alsuwaidi. 2021. "Exacerbation of Influenza A Virus Disease Severity by Respiratory Syncytial Virus Co-Infection in a Mouse Model" Viruses 13, no. 8: 1630. https://doi.org/10.3390/v13081630
APA StyleGeorge, J. A., AlShamsi, S. H., Alhammadi, M. H., & Alsuwaidi, A. R. (2021). Exacerbation of Influenza A Virus Disease Severity by Respiratory Syncytial Virus Co-Infection in a Mouse Model. Viruses, 13(8), 1630. https://doi.org/10.3390/v13081630