
Cytochrome P450 Monooxygenases CYP6AY3 and CYP6CW1 Regulate Rice Black-Streaked Dwarf Virus Replication in Laodelphax striatellus (Fallén)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Quantitative Real-Time PCR (RT-qPCR)
2.3. RNA Interference (RNAi)
2.4. Insecticide Treatment
2.5. Treatment of P450 Inhibitor PBO
2.6. Identification of P450 Genes in CYP3 and CYP4 Clades
2.7. Preparation of Different Tissues and L. striatellus at Different Developmental Stages
2.8. Analyzing the Effect of RBSDV Infection on P450 Genes Expression
2.9. Immunostaining
2.10. Data Analysis
3. Results
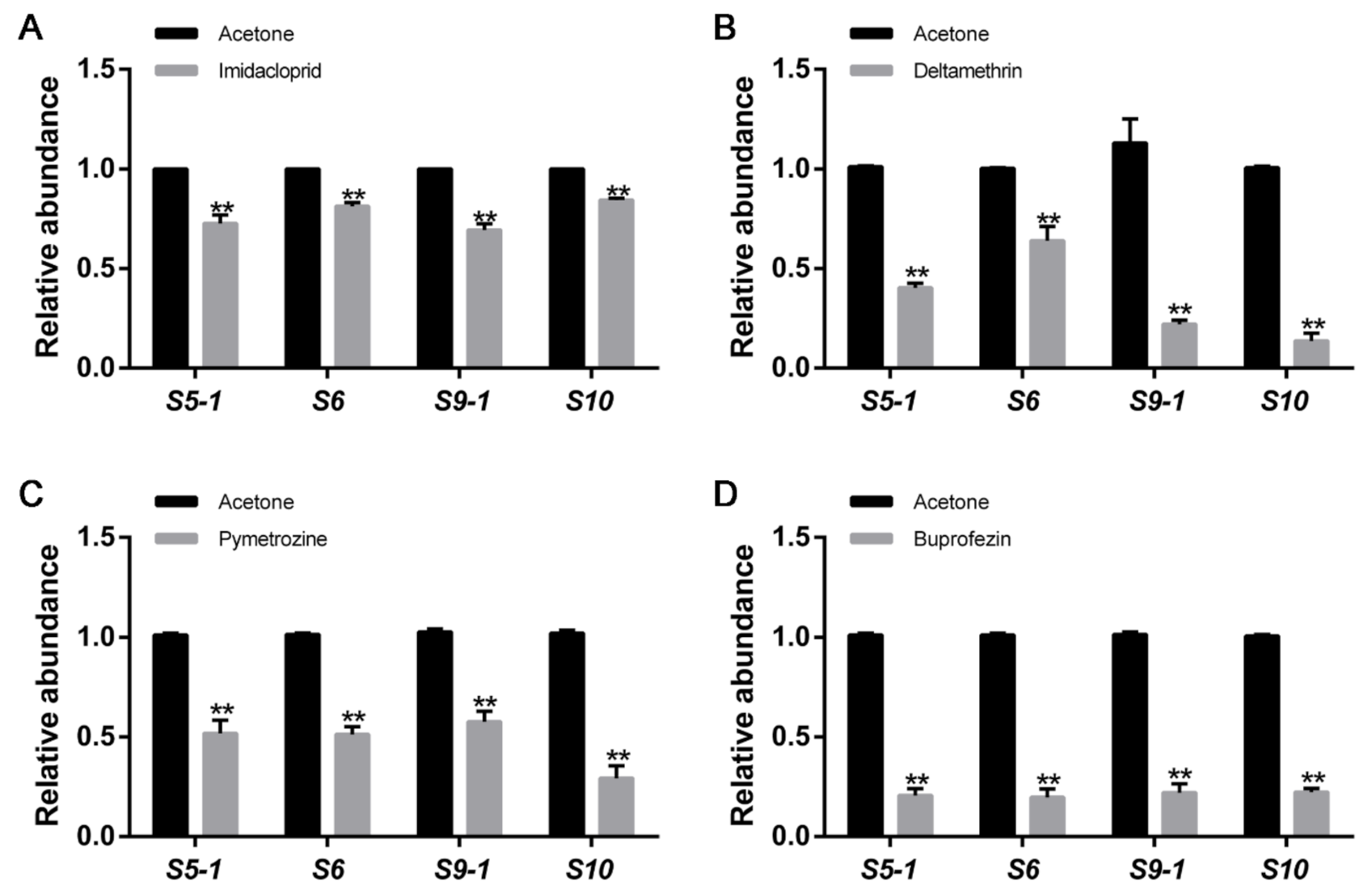
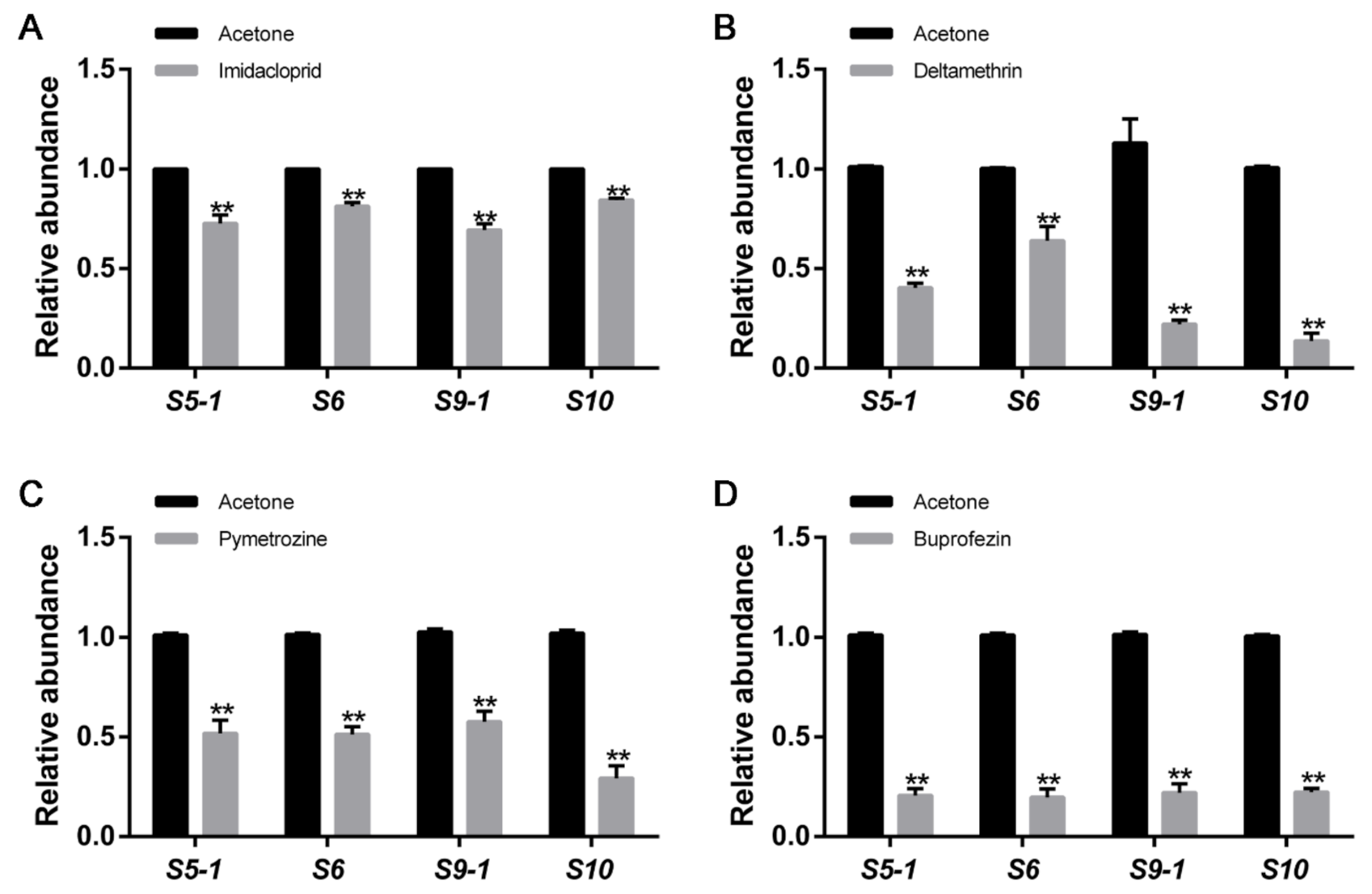
3.1. Application of Four Insecticides Inhibits RBSDV Abundance in L. striatellus
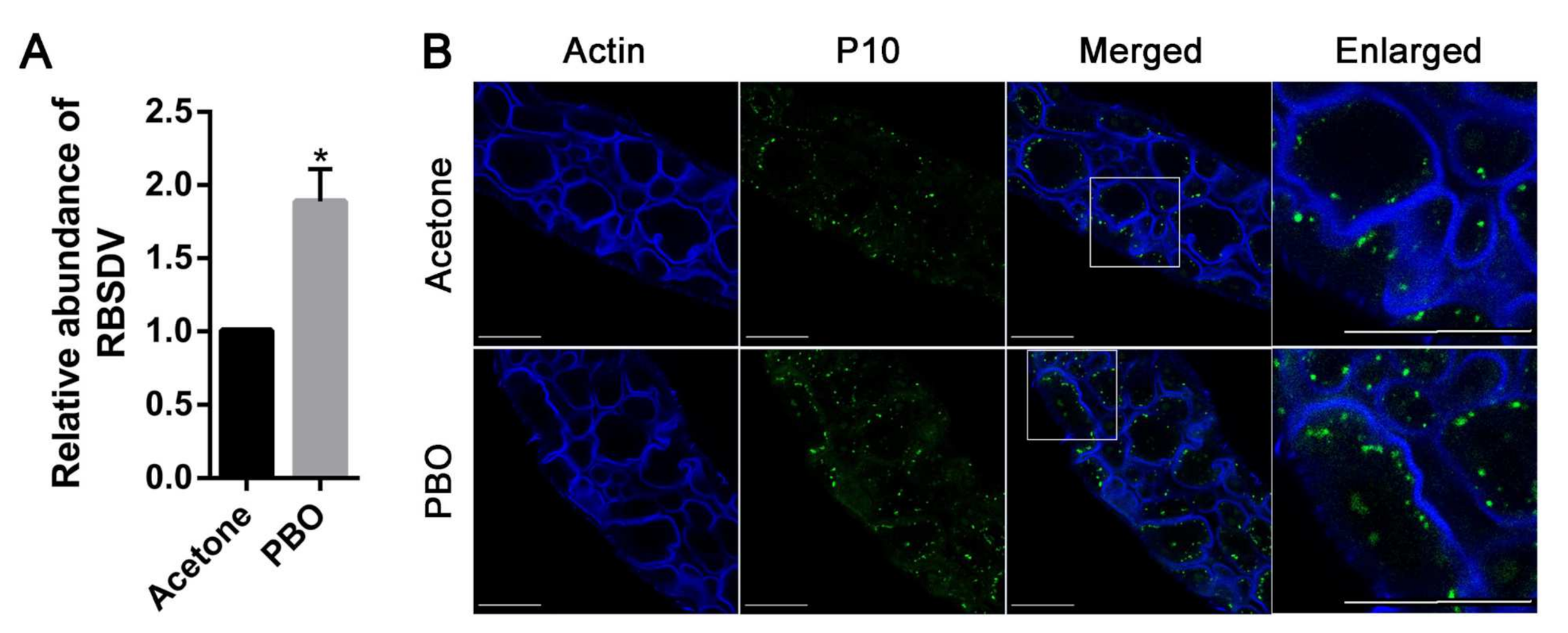
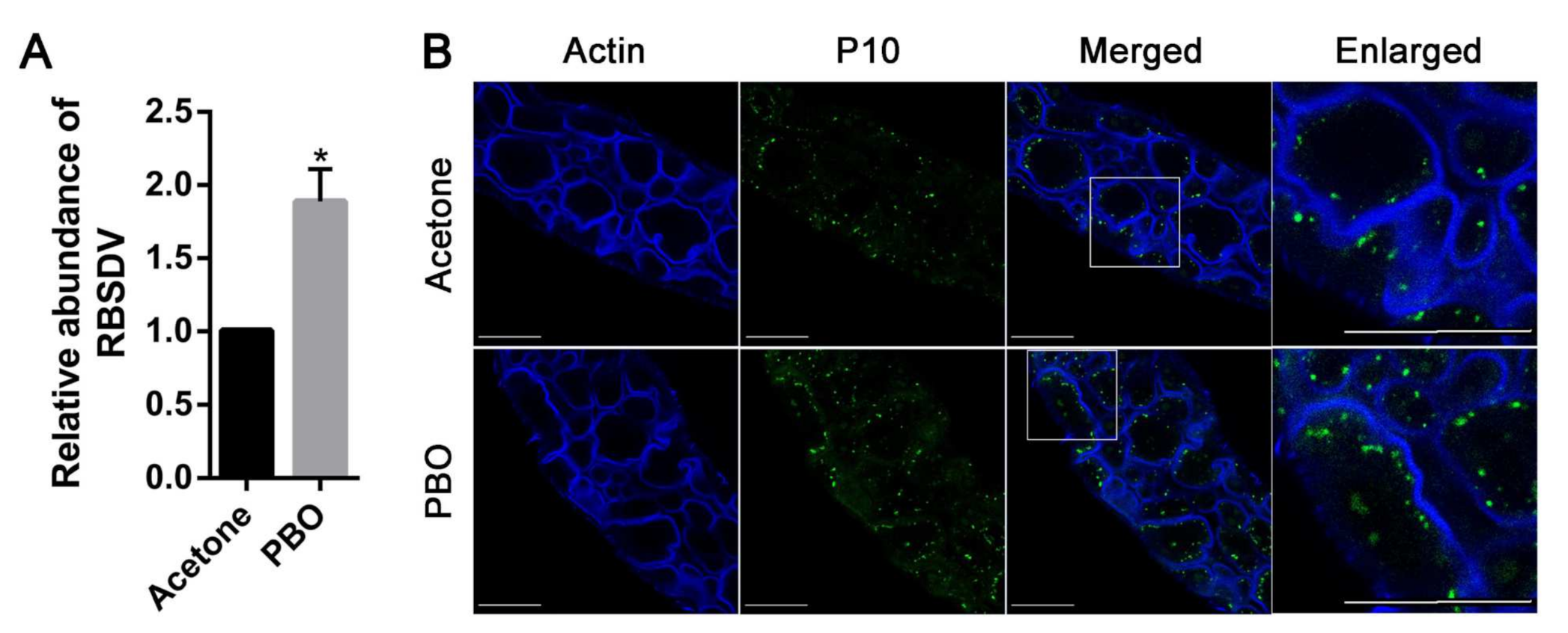
3.2. Inhibition of P450s Promotes RBSDV Infection in L. striatellus
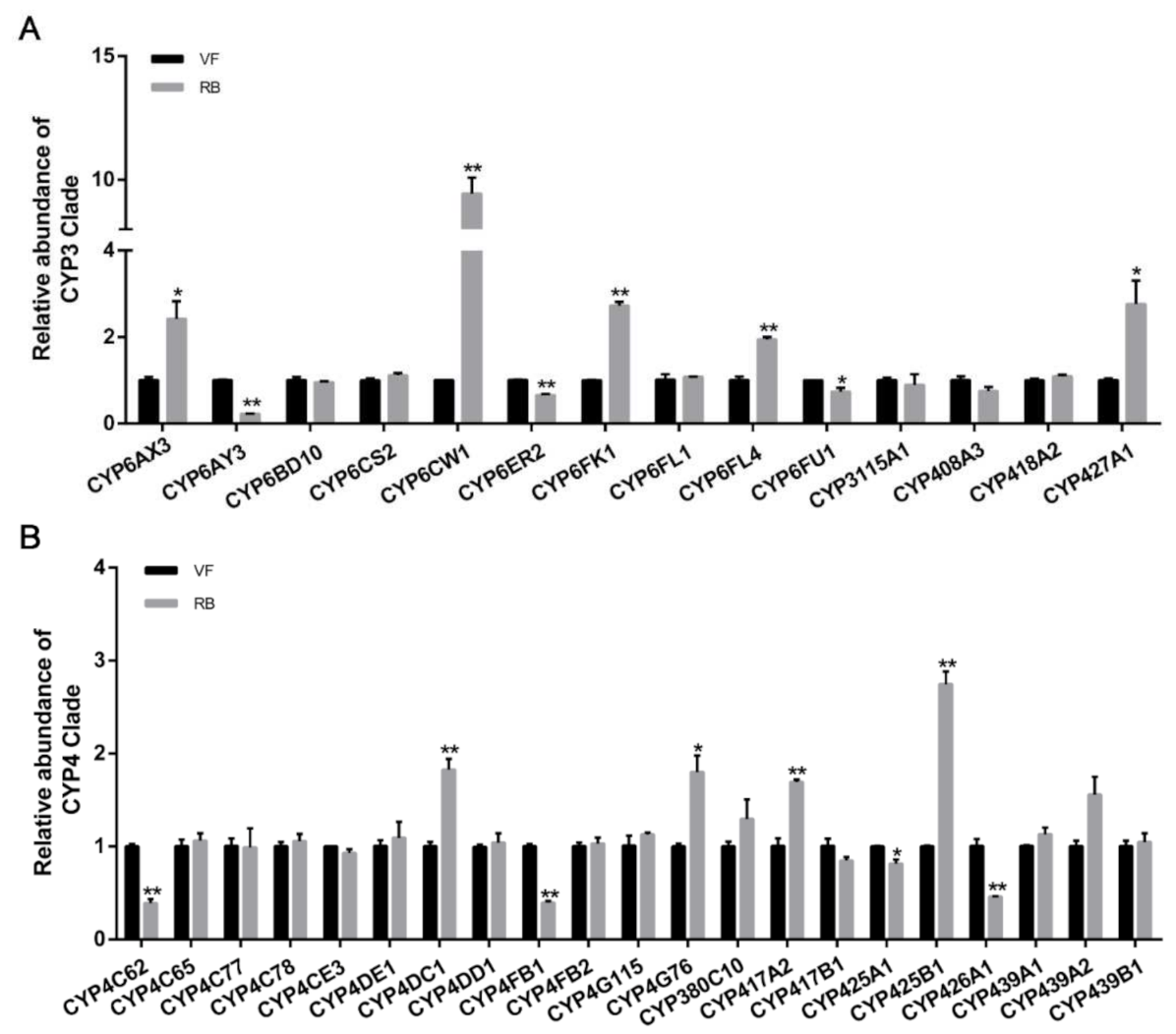
3.3. Transcriptional Response of Thirty-Five P450s under RBSDV Infection
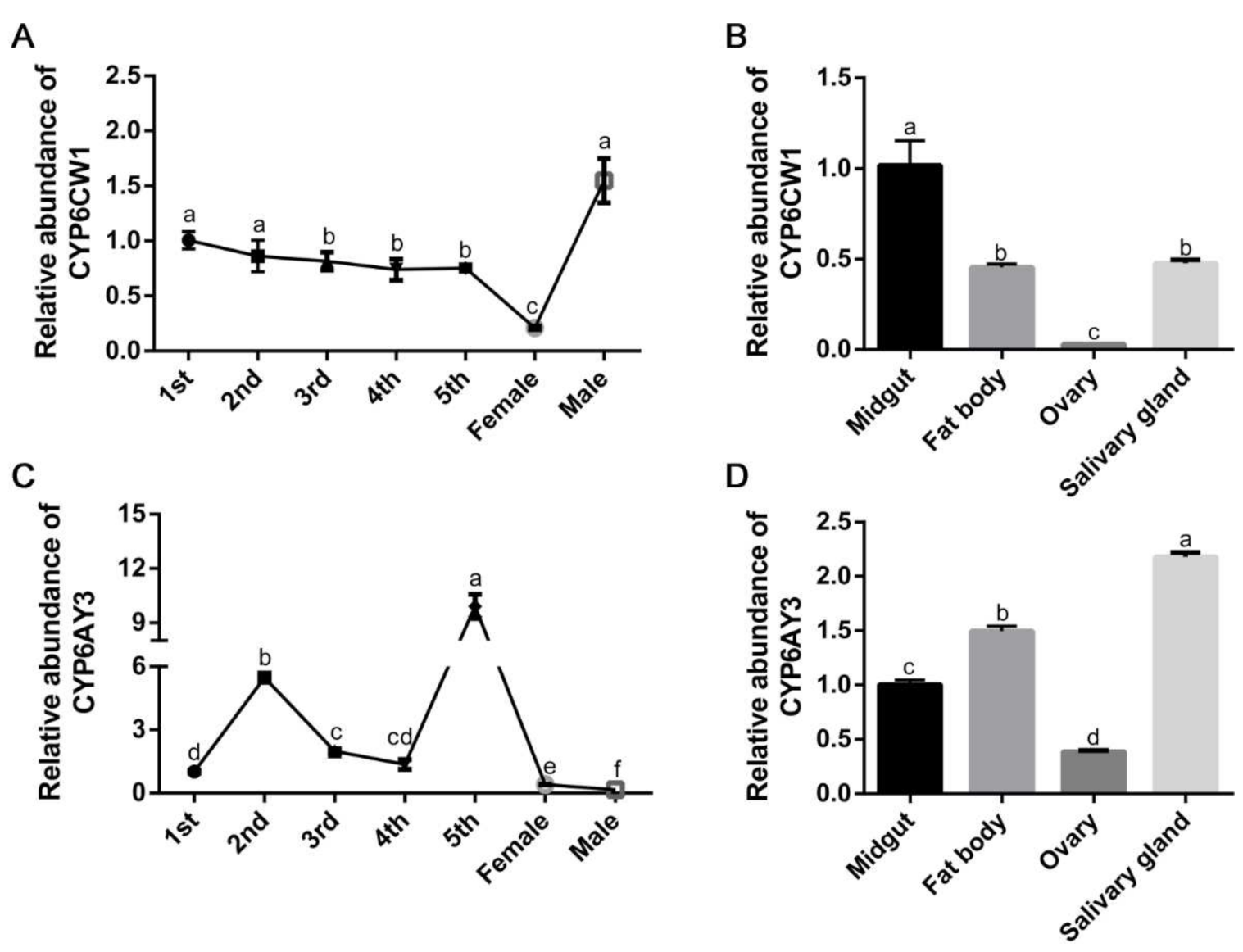
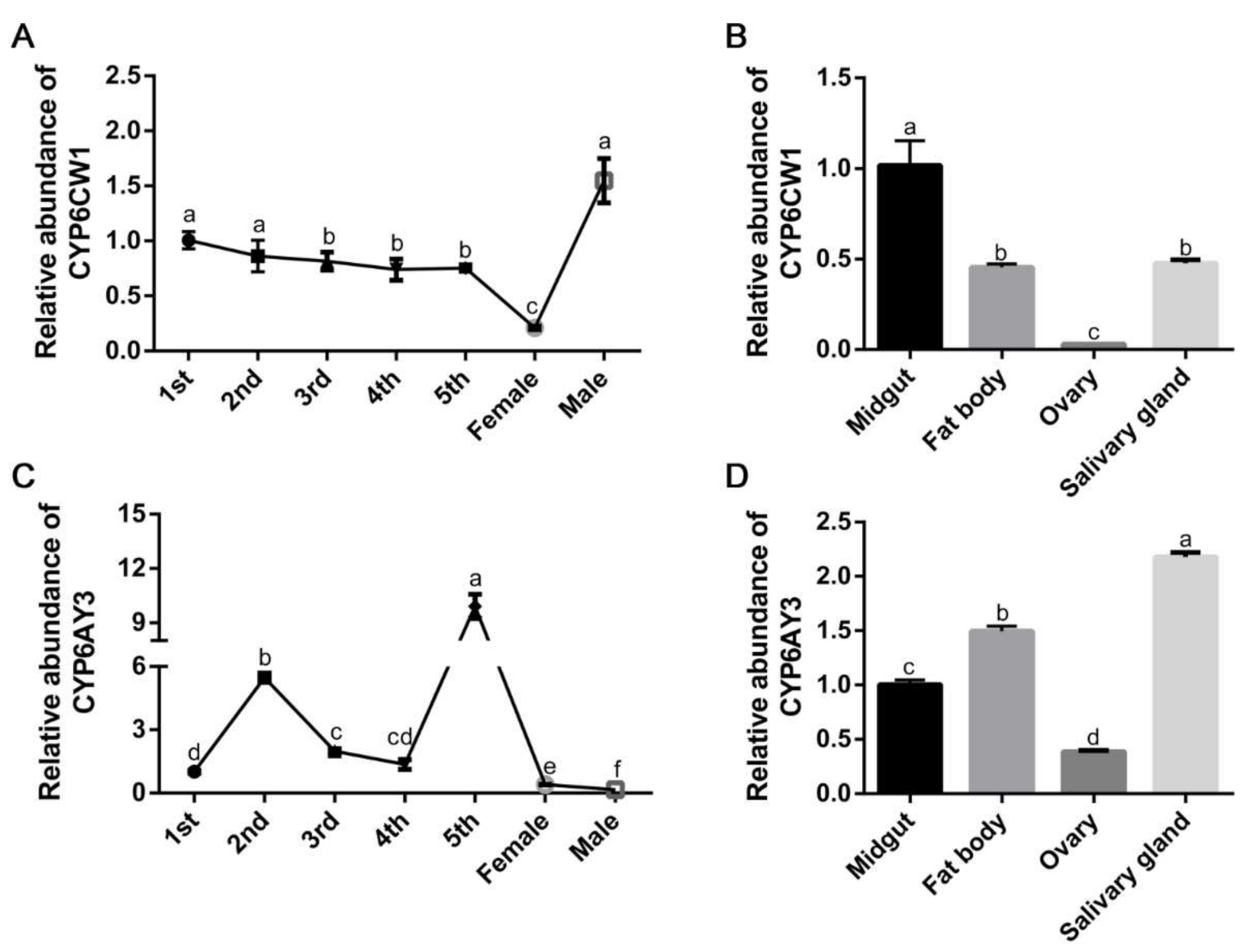
3.4. The Expression Profiles of CYP6CW1 and CYP6AY3 in L. striatellus
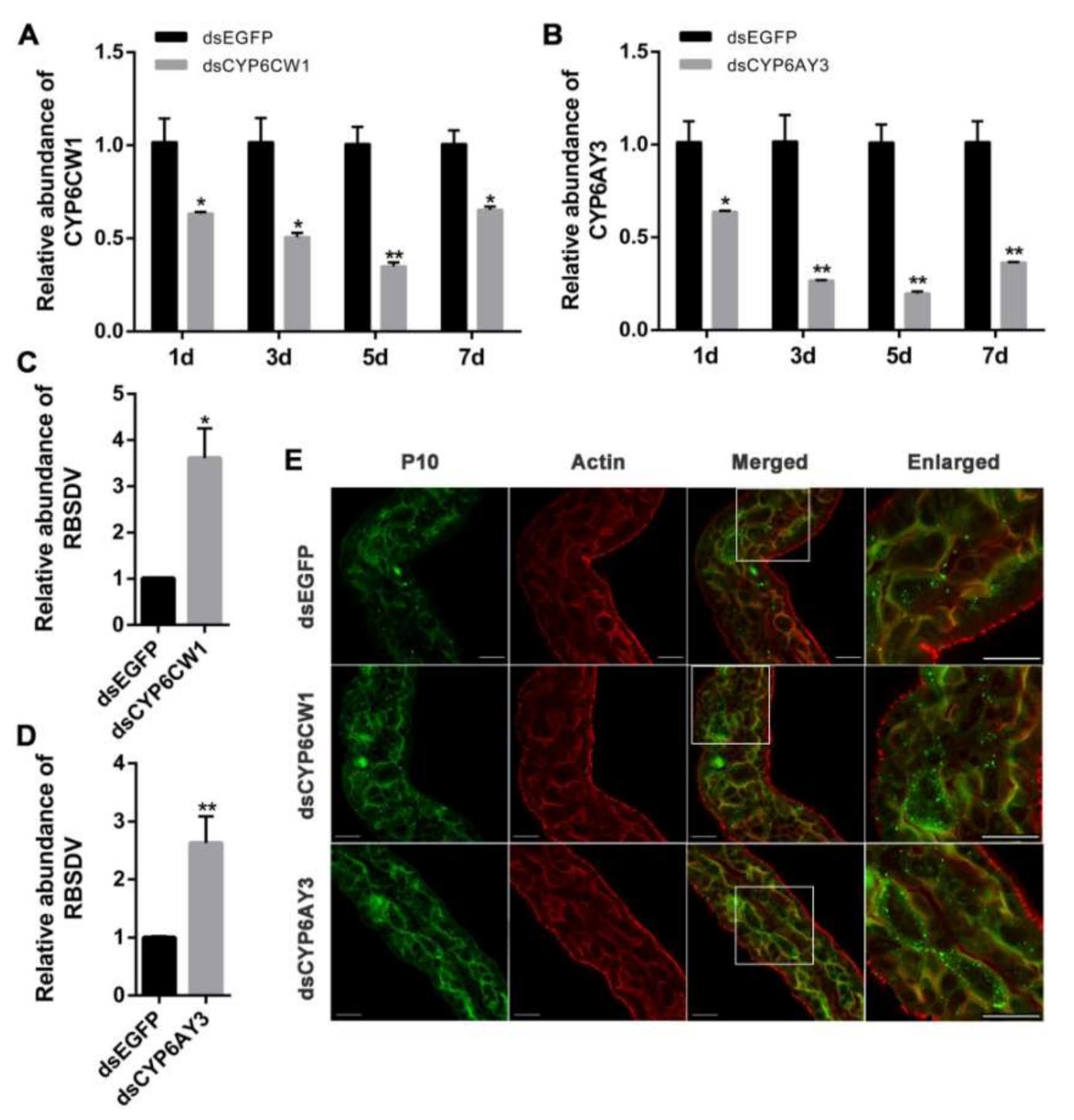
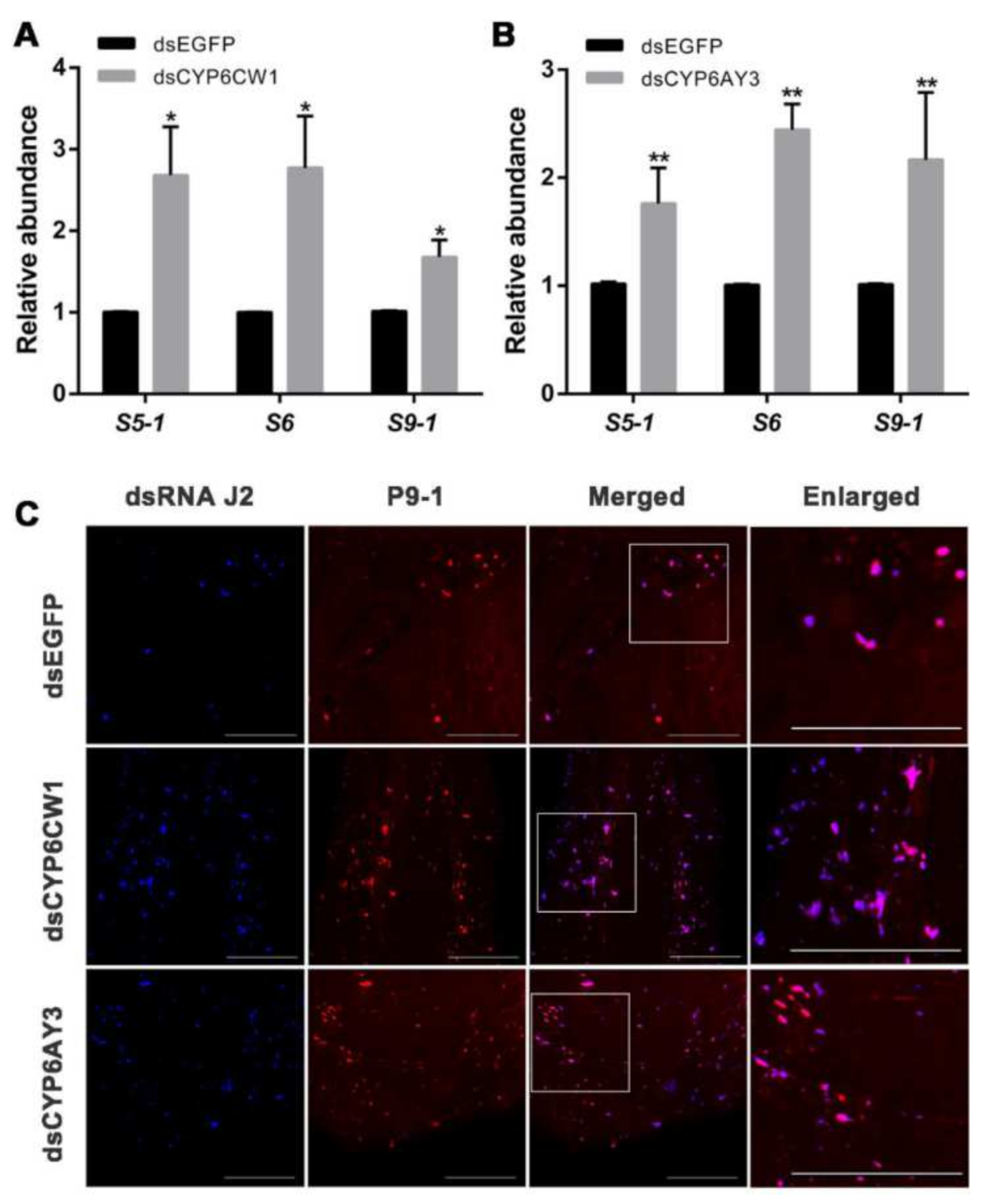
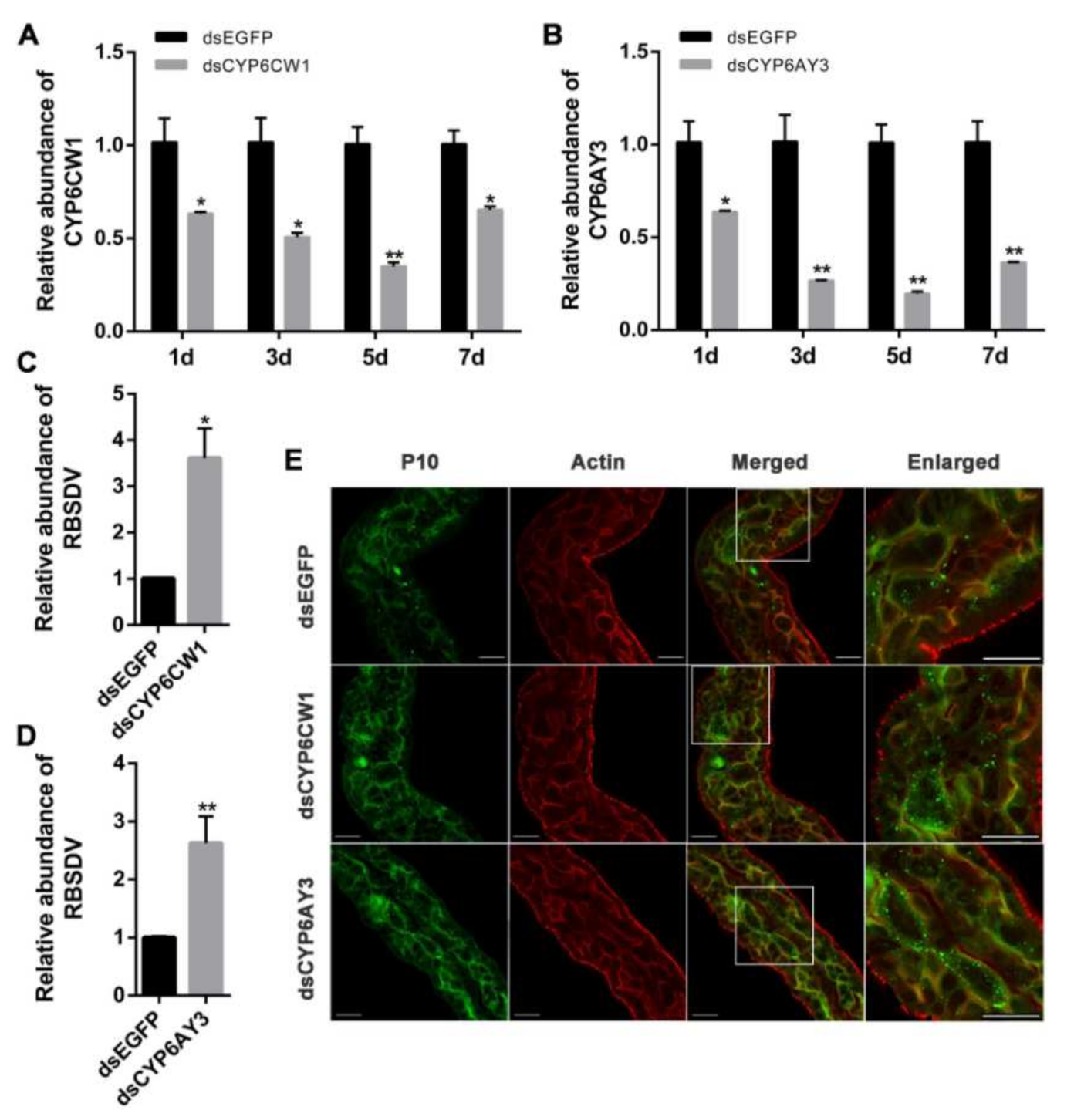
3.5. Knockdown of CYP6CW1 and CYP6AY3 Promotes RBSDV Infection
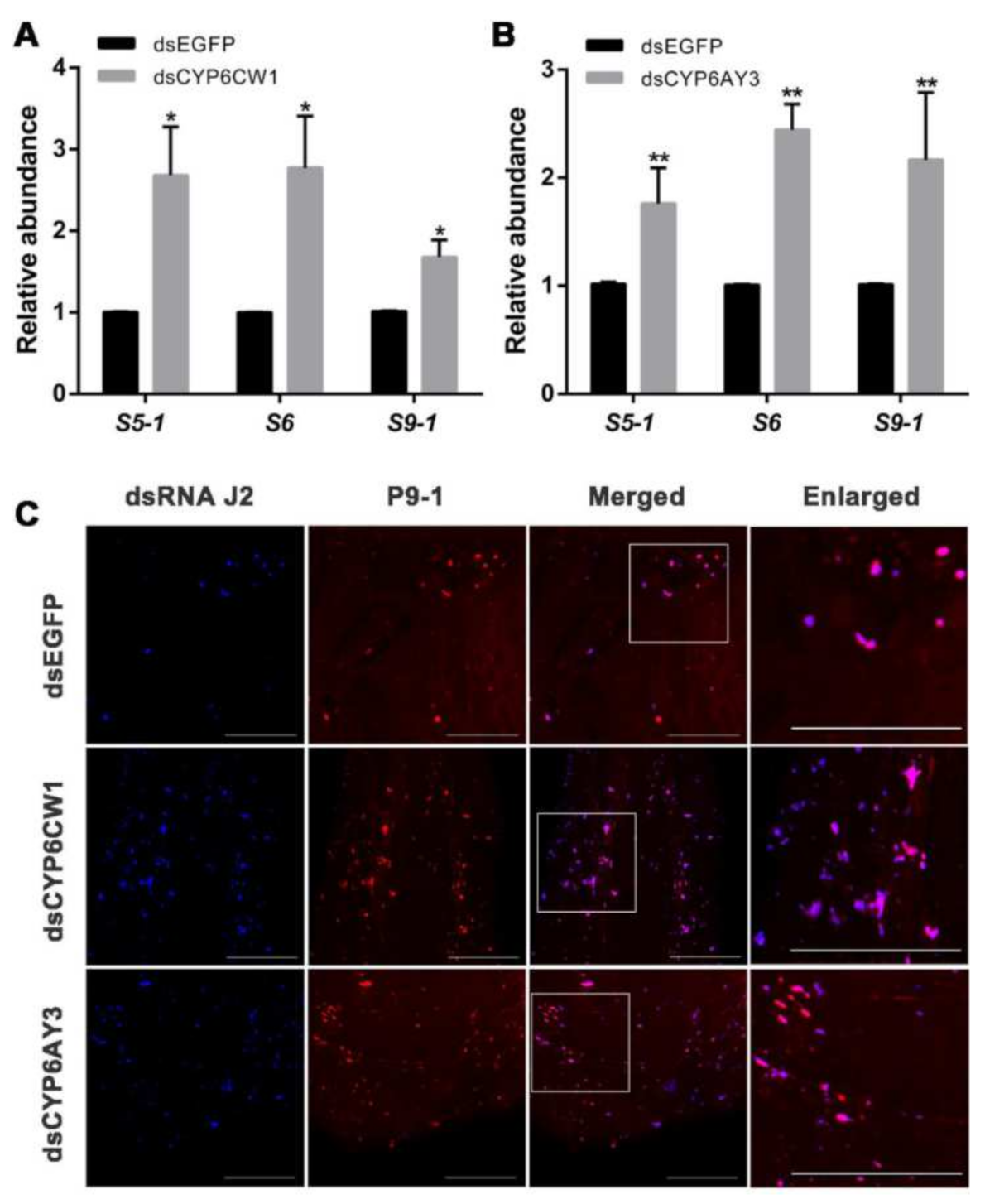
3.6. CYP6CW1 and CYP6AY3 Negatively Regulates the Replication of Viruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pu, L.; Xie, G.; Ji, C.; Ling, B.; Zhang, M.; Xu, D.; Zhou, G. Transmission characteristics of Southern rice black-streaked dwarf virus by rice planthoppers. Crop. Prot. 2012, 41, 71–76. [Google Scholar] [CrossRef]
- Mohammed, N.; Sharoni, A.M.; Kouji, S.; Rezaul, K.M.; Harikrishna, J.A.; Takumi, S.; Kikuchi, S.; Ahmad, A.; Hasan, S.M.Z.; Omura, T.; et al. NAC transcription factor family genes are differentially expressed in rice during infections with Rice dwarf virus, Rice black-streaked dwarf virus, Rice grassy stunt virus, Rice ragged stunt virus, and Rice transitory yellowing virus. Front. Plant Sci. 2015, 6, 676. [Google Scholar]
- Falk, B.W.; Tsai, J.H. Biology and molecular biology of viruses in the genus. Tenuivirus Annu. Rev. Phytopathol. 1998, 36, 139–163. [Google Scholar]
- Wu, N.; Zhang, L.; Ren, Y.D.; Wang, X.F. Rice black-streaked dwarf virus: From multiparty interactions among plant–virus–vector to intermittent epidemics. Mol. Plant Pathol 2020, 21, 1007–1019. [Google Scholar]
- Tanno, F.; Nakatsu, A.; Toriyama, S.; Kojima, M. Complete nucleotide sequence of Northern cereal mosaic virus and its genome organization. Arch. Virol. 2000, 145, 1373–1384. [Google Scholar] [CrossRef]
- Di, D.P.; Zhang, Y.; Yan, C.; Yan, T.; Zhang, A.H.; Yang, F.; Cao, X.L.; Li, D.; Lu, Y.G.; Wang, X.B.; et al. First Report of Barley yellow striate mosaic virus on Wheat in China. Plant Dis. 2014, 98, 1450. [Google Scholar]
- Hajano, J.; Raza, A.; Zhang, L.; Liu, W.; Wang, X. Ribavirin targets sugar transporter 6 to suppress acquisition and transmission of rice stripe tenuivirus by its vector Laodelphax striatellus. Pest Manag. Sci. 2020, 76, 4086–4092. [Google Scholar]
- Guengerich, F.P. Common and Uncommon Cytochrome P450 Reactions Related to Metabolism and Chemical Toxicity. Chem. Res. Toxicol. 2001, 14, 611–650. [Google Scholar]
- Feyereisen, R. Insect P450 enzymes. Annu Rev. Entomol. 1999, 44, 507–533. [Google Scholar] [CrossRef]
- Karunker, I.; Benting, J.; Lueke, B.; Ponge, T.; Nauen, R.; Roditakis, E.; Vontas, J.; Gorman, K.; Denholm, I.; Morin, S. Over-expression of cytochrome P450 CYP6CM1 is associated with high resistance to imidacloprid in the B and Q biotypes of Bemisia tabaci (Hemiptera: Aleyrodidae). Insect Biochem. Mol. Biol. 2008, 38, 634–644. [Google Scholar]
- Zhang, Y.; Yang, Y.; Sun, H.; Liu, Z. Metabolic imidacloprid resistance in the brown planthopper, Nilaparvata lugens, relies on multiple P450 enzymes. Insect Biochem. Mol. Biol. 2016, 79, 50–56. [Google Scholar]
- Elzaki, M.E.A.; Zhang, W.; Feng, A.; Qiou, X.; Zhao, W.; Han, Z. Constitutive overexpression of cytochrome P450 associated with imidacloprid resistance in Laodelphax striatellus (Fallén). Pest Manag. Sci. 2015, 72, 1051–1058. [Google Scholar]
- Wang, R.; Deng, L.; Song, P.; Elzaki, M.E.A.; Han, Z.; Wu, M.; Zhu, Y.; Zhang, H.; Yin, M. Imidacloprid is hydroxylated by Laodelphax striatellus CYP6AY3v2. Insect Mol. Biol. 2017, 26, 543–551. [Google Scholar]
- Zhang, Y.; Han, Y.; Liu, B.; Yang, Q.; Guo, H.; Liu, Z.; Wang, L.; Fang, J. Resistance monitoring and cross-resistance role of CYP6CW1 between buprofezin and pymetrozine in field populations of Laodelphax striatellus (Fallén). Sci. Rep. 2017, 7, 14639. [Google Scholar]
- Fattahi, S.; Alivije, M.K.; Babamahmoodi, F.; Bayani, M.; Zavareh, M.S.H.; Asouri, M.; Lotfi, M.; Amirbozorgi, G.; Akhavan-Niaki, H. Cytochrome P450 Genes (CYP2E1 and CYP1A1) Variants and Susceptibility to Chronic Hepatitis B Virus Infection. Indian J. Clin. Biochem. 2017, 33, 467–472. [Google Scholar]
- Zhang, L.; Wu, N.; Ren, Y.; Wang, X. Insights Into Insect Vector Transmission and Epidemiology of Plant-Infecting Fijiviruses. Front. Microbiol. 2021, 12, 628262. [Google Scholar] [CrossRef]
- Sun, L.; Xie, L.; Andika, I.B.; Tan, Z.; Chen, J. Non-structural protein P6 encoded by rice black-streaked dwarf virus is recruited to viral inclusion bodies by binding to the viroplasm matrix protein P9-1. J. Gen. Virol. 2013, 94, 1908–1916. [Google Scholar]
- He, L.; Chen, X.; Yang, J.; Zhang, T.; Li, J.; Zhang, S.; Zhong, K.; Zhang, H.; Chen, J. Rice black-streaked dwarf virus-encoded P5-1 regulates the ubiquitination activity of SCF E3 ligases and inhibits jasmonate signaling to benefit its infection in rice. New Phytol. 2019, 225, 896–912. [Google Scholar]
- Li, J.; Xue, J.; Zhang, H.-M.; Yang, J.; Lv, M.-F.; Xie, L.; Meng, Y.; Li, P.-P.; Chen, J.-P. Interactions between the P6 and P5-1 proteins of southern rice black-streaked dwarf fijivirus in yeast and plant cells. Arch. Virol. 2013, 158, 1649–1659. [Google Scholar]
- Lu, L.; Wang, Q.; Huang, D.; Xu, Q.; Zhou, X.; Wu, J. Rice black-streaked dwarf virus P10 suppresses protein kinase C in insect vector through changing the subcellular localization of LsRACK1. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180315. [Google Scholar]
- Zhang, H.H.; Tan, X.X.; He, Y.Q.; Xie, K.L.; Li, L.L.; Wang, R.; Sun, Z.; Chen, J.; Yan, F.; Li, J.; et al. Rice black-streaked dwarf virus P10 acts as either a synergistic or antagonistic determinant during superinfection with related or unrelated virus. Mol. Plant. Pathol. 2019, 20, 641–655. [Google Scholar]
- Jia, D.; Chen, Q.; Mao, Q.; Zhang, X.; Wu, W.; Chen, H.; Yu, X.; Wang, Z.; Wei, T. Vector mediated transmission of persistently transmitted plant viruses. Curr. Opin. Virol. 2018, 28, 127–132. [Google Scholar]
- Liao, X.; Xu, P.; Gong, P.; Wan, H.; Li, J. Current susceptibilities of brown planthopper Nilaparvata lugens to triflumezopyrim and other frequently used insecticides in China. Insect Sci. 2020, 28, 115–126. [Google Scholar]
- Xu, L.; Wu, M.; Han, Z. Overexpression of multiple detoxification genes in deltamethrin resistant Laodelphax striatellus (Hemiptera: Delphacidae) in China. PLoS ONE 2013, 8, e79443. [Google Scholar]
- Zhang, J.; Zhang, Y.; Wang, Y.; Yang, Y.; Cang, X.; Liu, Z. Expression induction of P450 genes by imidacloprid in Nilaparvata lugens: A genome-scale analysis. Pestic. Biochem. Physiol. 2016, 132, 59–64. [Google Scholar]
- Wu, W.; Liu, H.Q.; Dong, Y.; Zhang, Y.; Wong, S.M.; Wang, C.C.; Zhou, Y.J.; Xu, Q. Determination of suitable RT-qPCR reference genes for studies of gene functions in Laodelphax striatellus (Fallen). Genes 2019, 10, 887. [Google Scholar]
- Zhu, J.; Jiang, F.; Wang, X.; Yang, P.; Bao, Y.; Zhao, W.; Wang, W.; Lu, H.; Wang, Q.; Cui, N.; et al. Genome sequence of the small brown planthopper, Laodelphax striatellus. GigaScience 2017, 6, gix109. [Google Scholar]
- Maluta, N.K.P.; Lopes, J.R.S.; Fiallo-Olivé, E.; Navas-Castillo, J.; Lourenção, A.L. Foliar application of systemic insecticides disrupts feeding behavior of the whitefly Bemisia tabaci MEAM1 and the transmission of tomato chlorosis virus in potato plants. J. Pest Sci. 2021, 894, 1–12. [Google Scholar] [CrossRef]
- Wan, Y.; Zheng, X.; Xu, B.; Xie, W.; Wang, S.; Zhang, Y.; Zhou, X.; Wu, Q. Insecticide resistance increases the vector competence: A case study in Frankliniella occidentalis. J. Pest Sci. 2020, 94, 83–91. [Google Scholar]
- Wang, H.; Liu, Y.; Mo, L.; Huo, C.; Wang, Z.; Zhong, P.; Jia, D.; Zhang, X.; Chen, Q.; Chen, H.; et al. A neuron-specific antiviral mechanism modulates the persistent infection of rice Rhabdoviruses in leafhopper vectors. Front. Microbiol. 2020, 11, 513. [Google Scholar]
- Gourmet, C.; Kolb, F.L.; Smyth, C.A.; Pedersen, W.L. Use of imidacloprid as a seed-treatment insecticide to control barley yellow dwarf virus (BYDV) in oat and wheat. Plant Dis. 1996, 80, 136–141. [Google Scholar]
- Yang, Y.-X.; Lin, R.-H.; Li, Z.; Wang, A.-Y.; Xue, C.; Duan, A.-L.; Zhao, M.; Zhang, J.-H. Function Analysis of P450 and GST Genes to Imidacloprid in Aphis craccivora (Koch). Front. Physiol. 2021, 11, 624287. [Google Scholar] [CrossRef]
- Yang, T.; Liu, N. Genome analysis of cytochrome p450s and their expression profiles in insecticide resistant mosquitoes, Culex quinquefasciatus. PLoS ONE 2011, 6, e29418. [Google Scholar]
- Ding, T.-B.; Li, J.; Chen, E.-H.; Niu, J.-Z.; Chu, D. Transcriptome profiling of the whitefly Bemisia tabaci med in response to single infection of Tomato yellow leaf curl virus, Tomato chlorosis virus, and Their Co-infection. Front. Physiol. 2019, 10, 302. [Google Scholar]
- Zhang, Y.; Guo, H.; Yang, Q.; Li, S.; Wang, L.; Zhang, G.; Fang, J. Overexpression of a P450 gene (CYP6CW1) in buprofezin-resistant Laodelphax striatellus (Fallén). Pestic. Biochem. Physiol. 2012, 104, 277–282. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.-H.; Zhao, M.; Zhou, Y.-J.; Xu, Q.-F.; Yang, Y.-X. Cytochrome P450 Monooxygenases CYP6AY3 and CYP6CW1 Regulate Rice Black-Streaked Dwarf Virus Replication in Laodelphax striatellus (Fallén). Viruses 2021, 13, 1576. https://doi.org/10.3390/v13081576
Zhang J-H, Zhao M, Zhou Y-J, Xu Q-F, Yang Y-X. Cytochrome P450 Monooxygenases CYP6AY3 and CYP6CW1 Regulate Rice Black-Streaked Dwarf Virus Replication in Laodelphax striatellus (Fallén). Viruses. 2021; 13(8):1576. https://doi.org/10.3390/v13081576
Chicago/Turabian StyleZhang, Jian-Hua, Ming Zhao, Yi-Jun Zhou, Qiu-Fang Xu, and Yuan-Xue Yang. 2021. "Cytochrome P450 Monooxygenases CYP6AY3 and CYP6CW1 Regulate Rice Black-Streaked Dwarf Virus Replication in Laodelphax striatellus (Fallén)" Viruses 13, no. 8: 1576. https://doi.org/10.3390/v13081576
APA StyleZhang, J.-H., Zhao, M., Zhou, Y.-J., Xu, Q.-F., & Yang, Y.-X. (2021). Cytochrome P450 Monooxygenases CYP6AY3 and CYP6CW1 Regulate Rice Black-Streaked Dwarf Virus Replication in Laodelphax striatellus (Fallén). Viruses, 13(8), 1576. https://doi.org/10.3390/v13081576