
An Outbreak of Dengue Virus Serotype 2 Cosmopolitan Genotype in Nepal, 2017

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
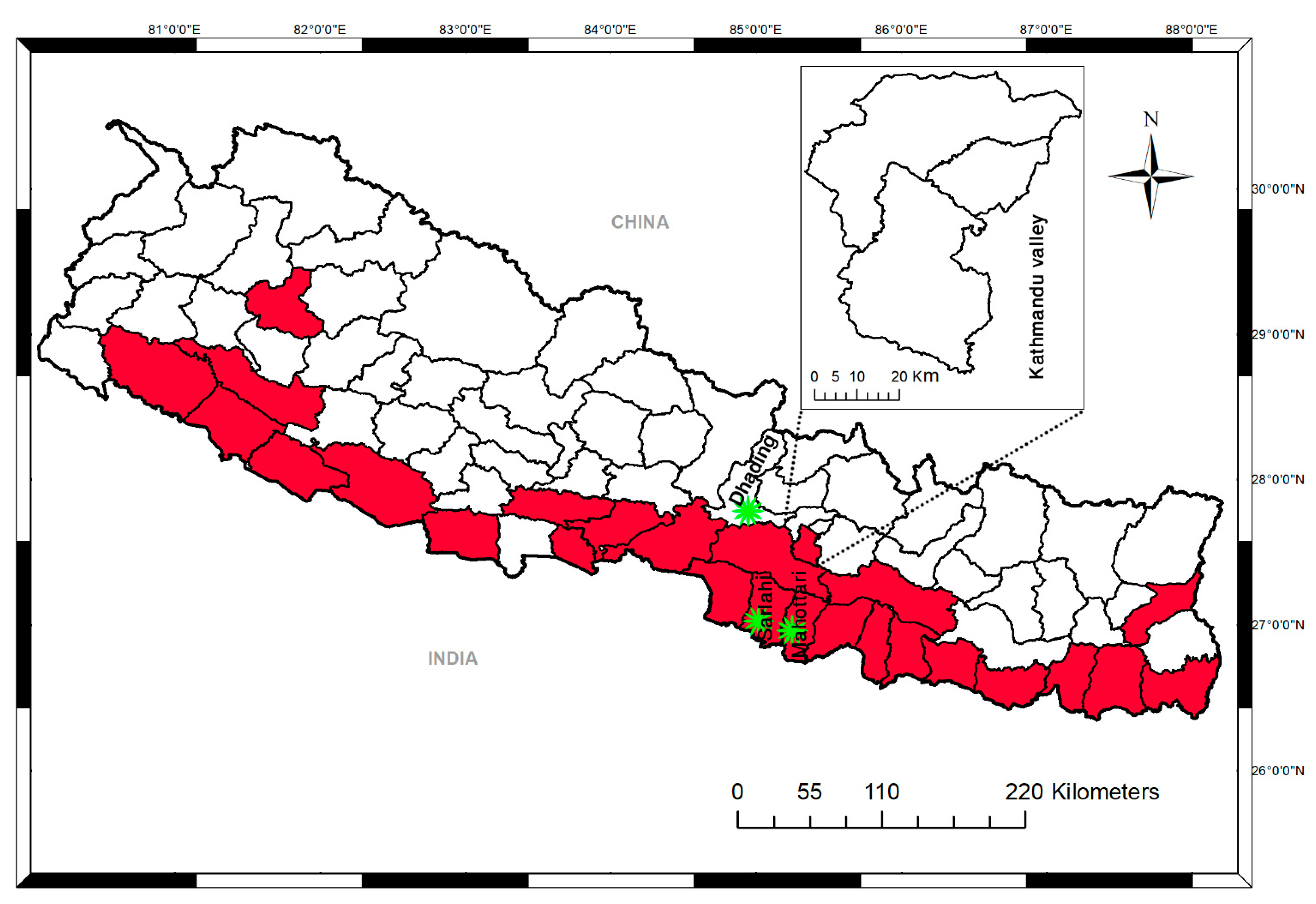
2.1. Study Design and Sites
2.2. Sample Collection
2.3. Serological Examination (Anti-DENV IgM and Anti-DENV IgG)
2.4. Virus Isolation in Cell Culture
2.5. RNA Extraction and Conventional Reverse Transcription (RT)-PCR
2.6. Quantification of DENV Genome Levels by Real-Time RT-PCR
2.7. Whole-Genome Sequencing by Next-Generation Sequencing (NGS) and Phylogenetic Tree Analysis
2.8. Statistical Analysis
3. Results
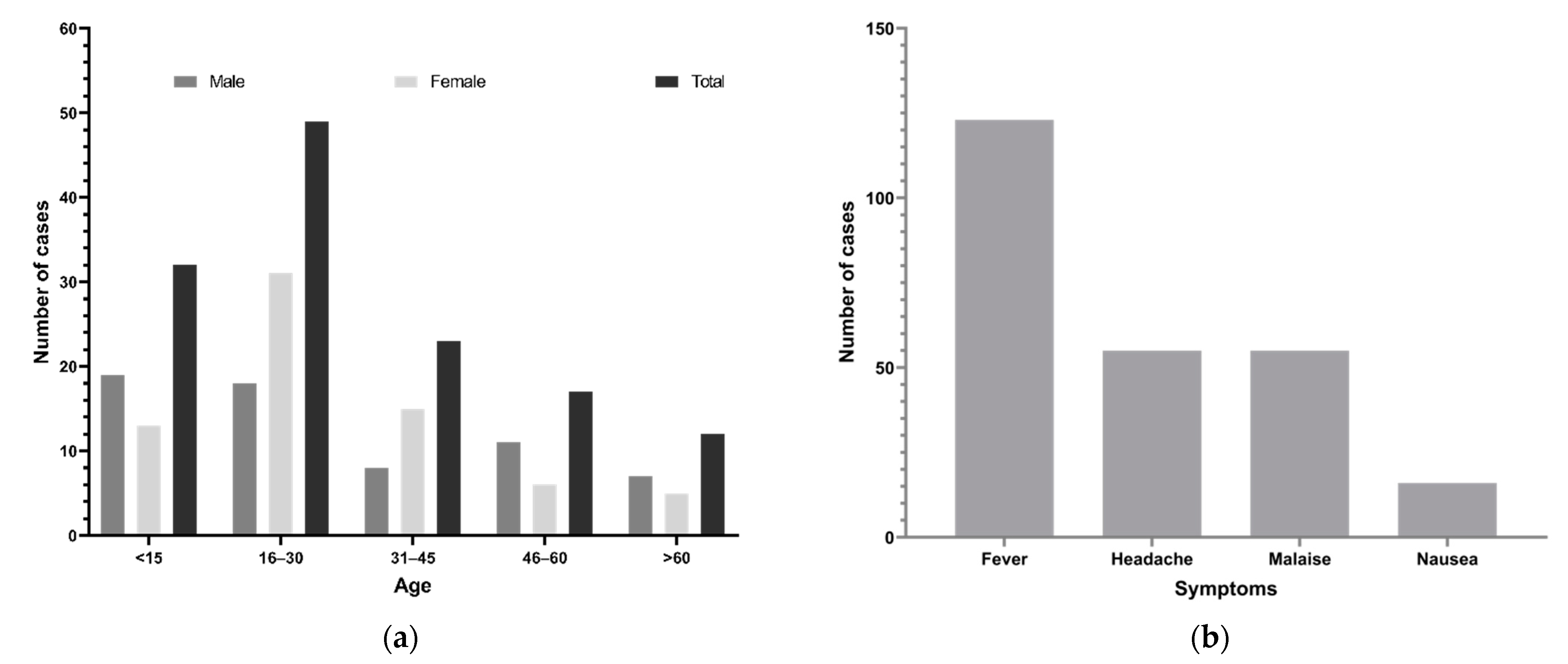
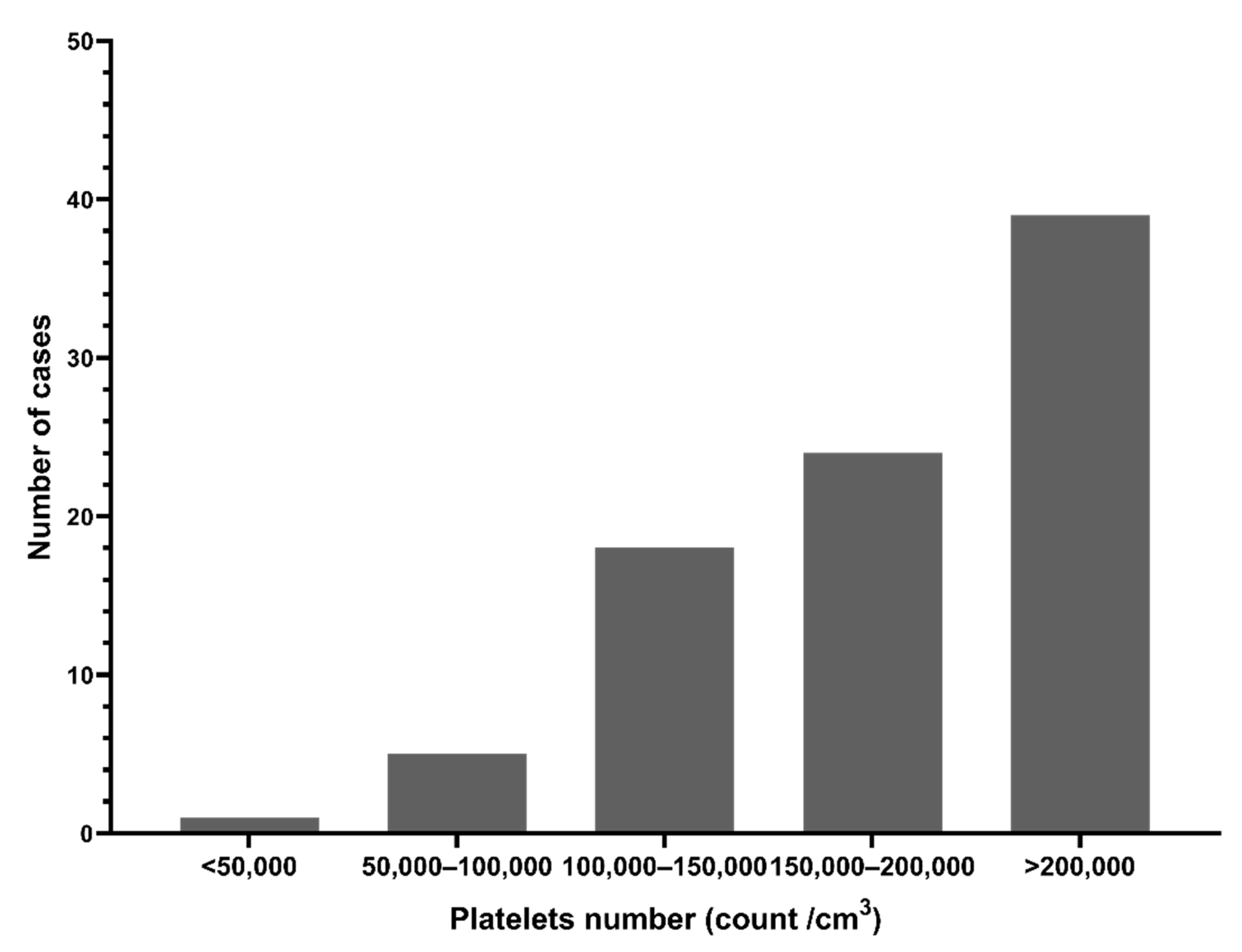
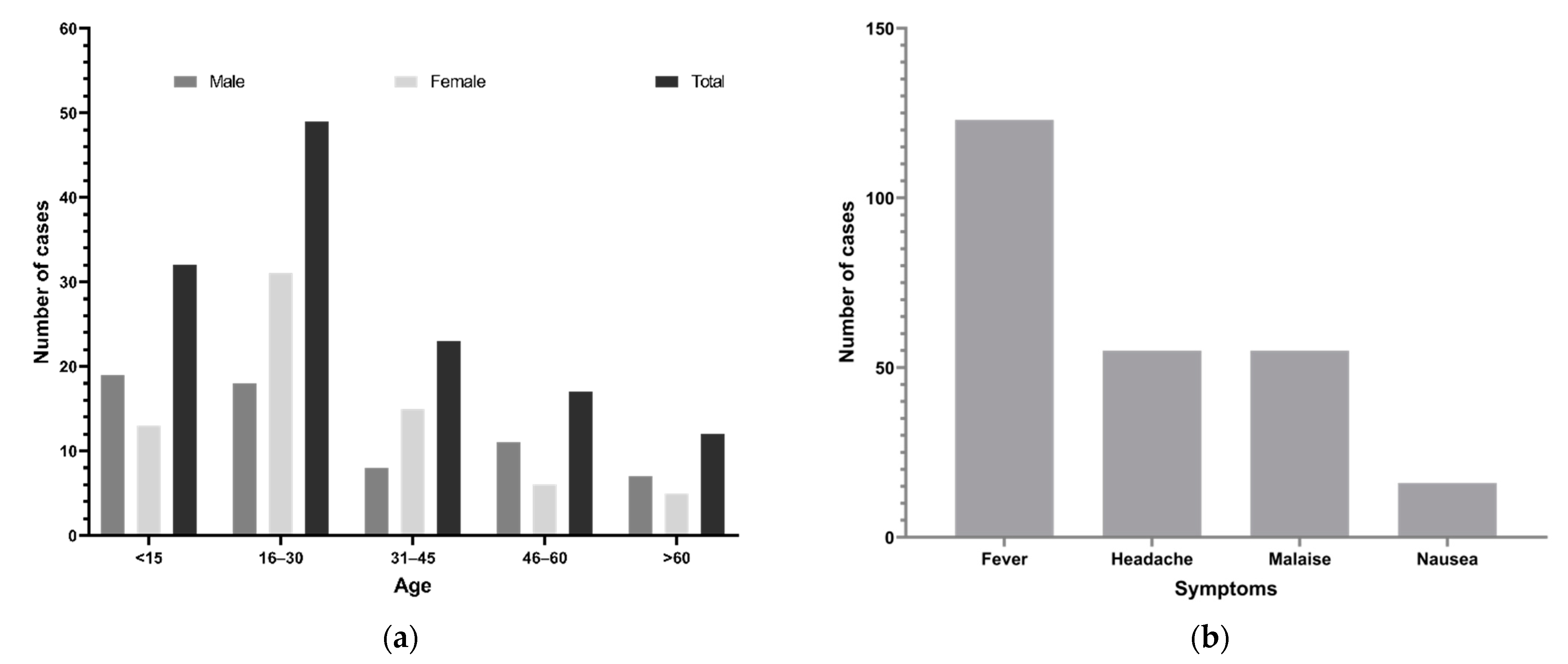
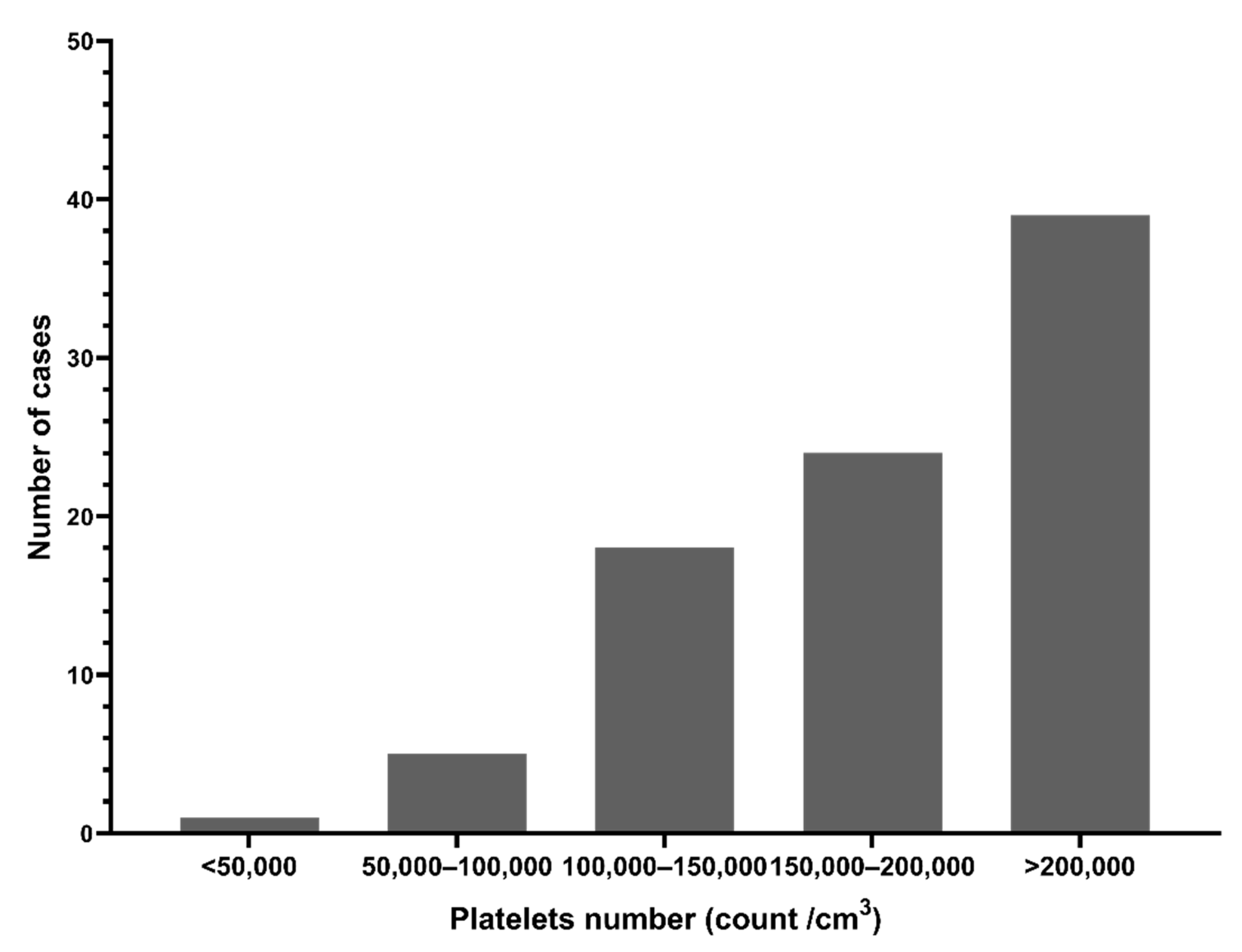
3.1. Demographics and Clinical Findings
3.2. Serological Findings
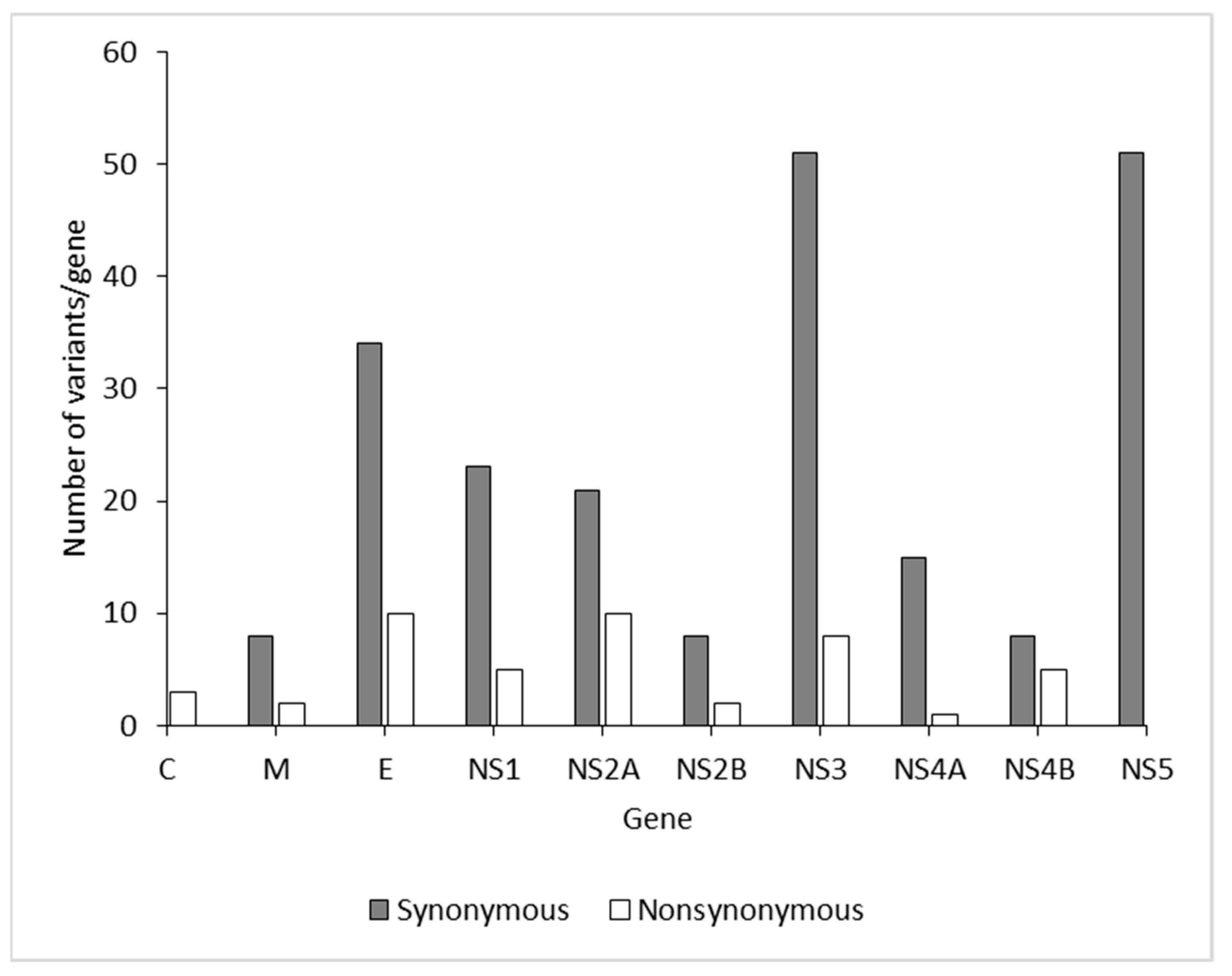
3.3. Molecular Findings
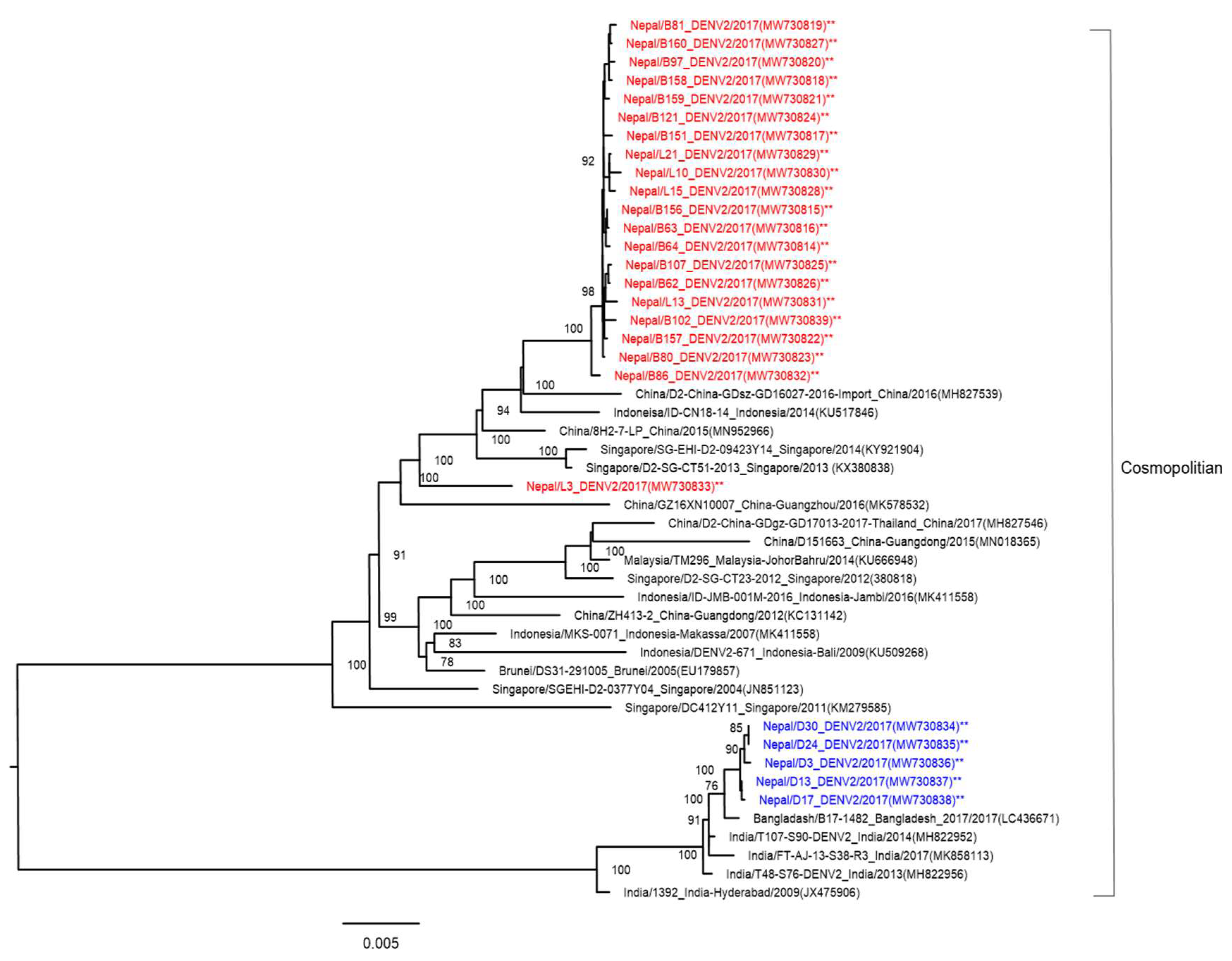
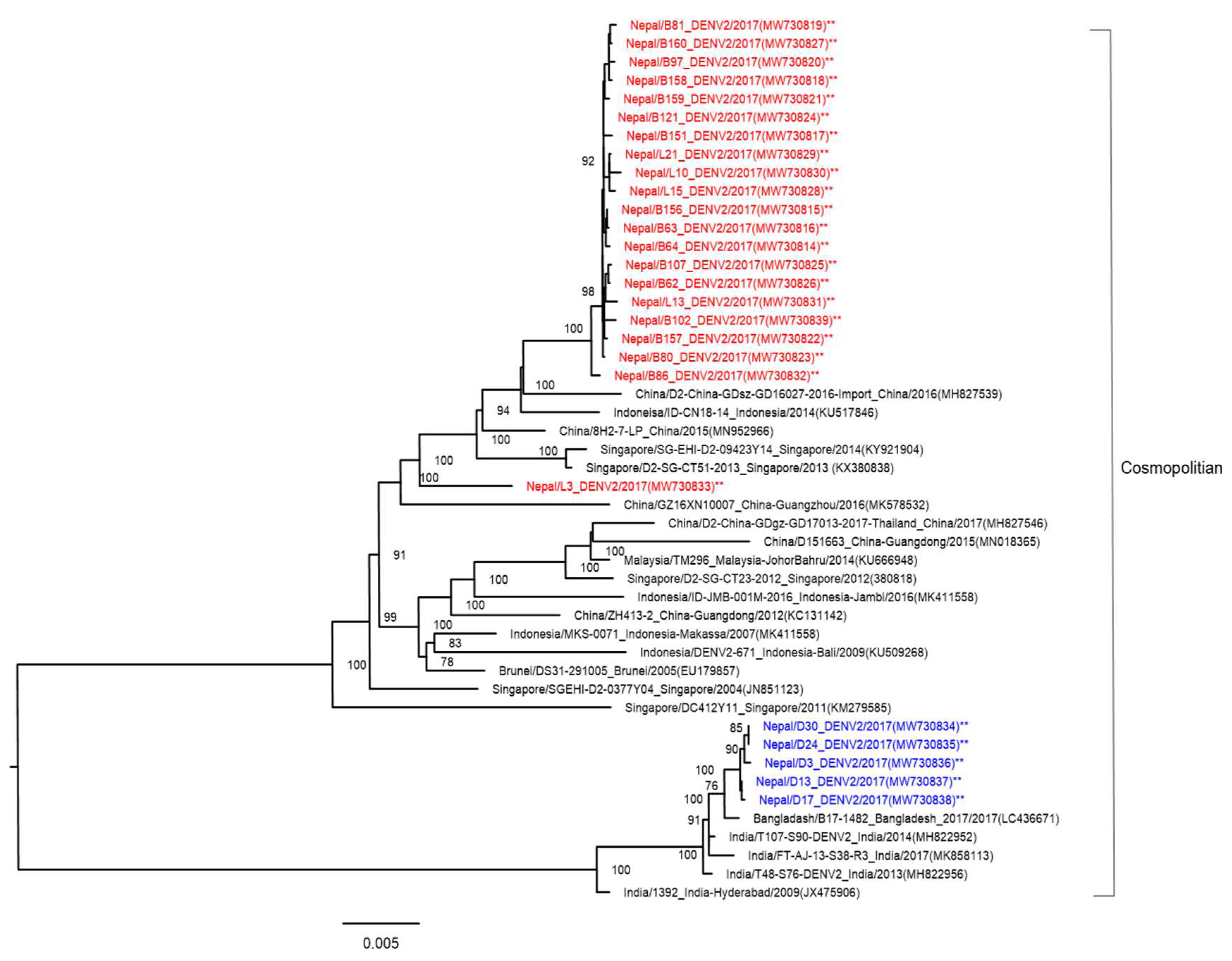
3.4. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. WHO|Dengue Haemorrhagic Fever: Diagnosis, Treatment, Prevention and Control, 2nd ed.; World Health Organization: Geneva, Switzerland, 1997. [Google Scholar]
- World Health Organization. Dengue Guidelines for Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Wilder-Smith, A.; Ooi, E.E.; Vasudevan, S.; Gubler, D.J. Update on Dengue: Epidemiology, Virus Evolution, Antiviral Drugs, and Vaccine Development. Curr. Infect. Dis. Rep. 2010, 12, 157–164. [Google Scholar] [CrossRef]
- Halstead, S.B. Dengue. Lancet 2007, 370, 1644–1652. [Google Scholar] [CrossRef]
- WHO. Dengue and Severe Dengue 2020. WHO Fact Sheet 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 30 May 2021).
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus genome organization, expression, and replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef]
- Owens, R.J.; Limn, C.; Roy, P. Role of an Arbovirus Nonstructural Protein in Cellular Pathogenesis and Virus Release. J. Virol. 2004, 78, 6649–6656. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Roche, R.; Gould, E.A. Understanding the Dengue Viruses and Progress towards Their Control. BioMed Res. Int. 2013, 2013, 690835. [Google Scholar] [CrossRef]
- Holmes, E.C.; Twiddy, S.S. The origin, emergence and evolutionary genetics of dengue virus. Infect. Genet. Evol. 2003, 3, 19–28. [Google Scholar] [CrossRef]
- WHO. Dengue and Severe Dengue Key Facts; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Pandey, B.D.; Rai, S.K.; Morita, K.; Kurane, I. First case of Dengue virus infection in Nepal. Nepal Med. Coll. J. NMCJ 2004, 6, 157–159. [Google Scholar]
- Pandey, B.D.; Nabeshima, T.; Pandey, K.; Rajendra, S.P.; Shah, Y.; Adhikari, B.R.; Gupta, G.; Gautam, I.; Tun, M.M.N.; Uchida, R.; et al. First Isolation of Dengue Virus from the 2010 Epidemic in Nepal. Trop. Med. Health 2013, 41, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Prajapati, S.; Napit, R.; Bastola, A.; Rauniyar, R.; Shrestha, S.; Lamsal, M.; Adhikari, A.; Bhandari, P.; Yadav, S.R.; Manandhar, K.D. Molecular phylogeny and distribution of dengue virus serotypes circulating in Nepal in 2017. PLoS ONE 2020, 15, e0234929. [Google Scholar] [CrossRef] [PubMed]
- Dumre, S.P.; Bhandari, R.; Shakya, G.; Shrestha, S.K.; Cherif, M.S.; Ghimire, P.; Klungthong, C.; Yoon, I.-K.; Hirayama, K.; Na-Bangchang, K.; et al. Dengue Virus Serotypes 1 and 2 Responsible for Major Dengue Outbreaks in Nepal: Clinical, Laboratory, and Epidemiological Features. Am. J. Trop. Med. Hyg. 2017, 97, 1062–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudyal, P.; Sharma, K.; Dumre, S.P.; Bastola, A.; Chalise, B.S.; Shrestha, B.; Poudel, A.; Giri, A.; Bhandari, P.; Shah, Y.; et al. Molecular study of 2019 dengue fever outbreaks in Nepal. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Dumre, S.P.; Acharya, D.; Lal, B.K.; Brady, O.J. Dengue virus on the rise in Nepal. Lancet Infect. Dis. 2020, 20, 889–890. [Google Scholar] [CrossRef]
- Wilder-Smith, A.; Murray, N.E.A.; Quam, M. Epidemiology of dengue: Past, present and future prospects. Clin. Epidemiol. 2013, 5, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bundo, K.; Igarashi, A. Antibody-capture ELISA for detection of immunoglobulin M antibodies in sera from japanese encephalitis and dengue hemorrhagic fever patients. J. Virol. Methods 1985, 11, 15–22. [Google Scholar] [CrossRef]
- Tun, M.M.N.; Thant, K.Z.; Inoue, S.; Kurosawa, Y.; Lwin, Y.Y.; Lin, S.; Aye, K.T.; Khin, P.T.; Myint, T.; Htwe, K.; et al. Serological characterization of dengue virus infections observed among dengue hemorrhagic fever/dengue shock syndrome cases in upper Myanmar. J. Med. Virol. 2013, 85, 1258–1266. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Alonzo, M.T.G.; Kurosawa, Y.; Mapua, C.A.; Reyes, J.D.; Dimaano, E.M.; Alera, M.T.P.; Saito, M.; Oishi, K.; Hasebe, F.; et al. Evaluation of a Dengue IgG Indirect Enzyme-Linked Immunosorbent Assay and a Japanese Encephalitis IgG Indirect Enzyme-Linked Immunosorbent Assay for Diagnosis of Secondary Dengue Virus Infection. Vector-Borne Zoonotic Dis. 2010, 10, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Tun, M.M.N.; Kyaw, A.K.; Makki, N.; Muthugala, R.; Nabeshima, T.; Inoue, S.; Hayasaka, D.; Moi, M.L.; Buerano, C.C.; Thwe, S.M.; et al. Characterization of the 2013 dengue epidemic in Myanmar with dengue virus 1 as the dominant serotype. Infect. Genet. Evol. 2016, 43, 31–37. [Google Scholar] [CrossRef]
- Tun, M.M.N.; Muthugala, R.; Nabeshima, T.; Rajamanthri, L.; Jayawardana, D.; Attanayake, S.; Soe, A.M.; Dumre, S.P.; Ando, T.; Hayasaka, D.; et al. Unusual, neurological and severe dengue manifestations during the outbreak in Sri Lanka, 2017. J. Clin. Virol. 2020, 125, 104304. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Morita, K.; Tanaka, M.; Igarashi, A. Rapid identification of dengue virus serotypes by using polymerase chain reaction. J. Clin. Microbiol. 1991, 29, 2107–2110. [Google Scholar] [CrossRef] [Green Version]
- Kyaw, A.K.; Tun, M.M.N.; Moi, M.L.; Nabeshima, T.; Soe, K.T.; Thwe, S.M.; Myint, A.A.; Maung, K.T.T.; Aung, W.; Hayasaka, D.; et al. Clinical, virological and epidemiological characterization of dengue outbreak in Myanmar, 2015. Epidemiol. Infect. 2017, 145, 1886–1897. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.S.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3. 0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Pandey, B.D.; Pandey, K.; Neupane, B.; Shah, Y.; Adhikary, K.P.; Gautam, I.; Hagge, D.A.; Morita, K. Persistent dengue emergence: The seven years surrounding the 2010 epidemic in Nepal. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Shah, Y.; Katuwal, A.; Pun, R.; Pant, K.; Sherchand, S.P.; Pandey, K.; Joshi, D.D.; Pandey, B.D. Dengue in western Terai region of Nepal. J. Nepal Health Res. Counc. 2012, 10, 152–155. [Google Scholar] [PubMed]
- Neupane, B.; Sherchand, J.B.; Pandey, B.D. Clinical observations among patients with Dengue Fever in Nepal. J. Inst. Med. 2014, 36, 92–94. [Google Scholar]
- Dumre, S.P.; Fernandez, S.; Shakya, G.; Nisalak, A.; Upreti, S.R.; Kc, K.; Gibbons, R.V.; Na-Bangchang, K.; Grams, R.; Ghimire, P.; et al. Dengue Virus and Japanese Encephalitis Virus Epidemiological Shifts in Nepal: A Case of Opposing Trends. Am. J. Trop. Med. Hyg. 2013, 88, 677–680. [Google Scholar] [CrossRef] [Green Version]
- Gupta, B.P.; Tuladhar, R.; Kurmi, R.; Manandhar, K. Das Dengue periodic outbreaks and epidemiological trends in Nepal. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 6. [Google Scholar] [CrossRef] [Green Version]
- Malla, S.; Thakur, G.D.; Shrestha, S.K.; Banjeree, M.K.; Thapa, L.B.; Gongal, G.; Ghimire, P.; Upadhyay, B.P.; Gautam, P.; Khanal, S.; et al. Identification of All Dengue Serotypes in Nepal. Emerg. Infect. Dis. 2008, 14, 1669–1670. [Google Scholar] [CrossRef] [PubMed]
- Lustig, Y.; Wolf, D.; Halutz, O.; Schwartz, E. An outbreak of dengue virus (DENV) type 2 Cosmopolitan genotype in Israeli travellers returning from the Seychelles, April 2017. Eurosurveillance 2017, 22, 30563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascalis, H.; Turpin, J.; Roche, M.; Krejbich, P.; Gadea, G.; Nten, C.A.; Desprès, P.; Mavingui, P. The epidemic of Dengue virus type-2 Cosmopolitan genotype on Reunion Island relates to its active circulation in the Southwestern Indian Ocean neighboring islands. Heliyon 2019, 5, e01455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phadungsombat, J.; Lin, M.Y.-C.; Srimark, N.; Yamanaka, A.; Nakayama, E.E.; Moolasart, V.; Suttha, P.; Shioda, T.; Uttayamakul, S. Emergence of genotype Cosmopolitan of dengue virus type 2 and genotype III of dengue virus type 3 in Thailand. PLoS ONE 2018, 13, e0207220. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Phadungsombat, J.; Nakayama, E.E.; Saito, A.; Egawa, A.; Sato, T.; Rahim, R.; Hasan, A.; Lin, M.Y.-C.; Takasaki, T.; et al. Genotype replacement of dengue virus type 3 and clade replacement of dengue virus type 2 genotype Cosmopolitan in Dhaka, Bangladesh in 2017. Infect. Genet. Evol. 2019, 75, 103977. [Google Scholar] [CrossRef]
- Yu, H.; Kong, Q.; Wang, J.; Qiu, X.; Wen, Y.; Yu, X.; Liu, M.; Wang, H.; Pan, J.; Sun, Z. Multiple Lineages of Dengue Virus Serotype 2 Cosmopolitan Genotype Caused a Local Dengue Outbreak in Hangzhou, Zhejiang Province, China, in 2017. Sci. Rep. 2019, 9, 7345. [Google Scholar] [CrossRef] [PubMed]
- Government of Nepal. Nepal Tourism Statistics; Ministry of Culture, Tourism & Civil Aviation, Singha Durbar: Kathmandu, Nepal, 2019. Available online: https://www.tourism.gov.np/files/NOTICE%20MANAGER_FILES/Nepal_%20tourism_statics_2019.pdf (accessed on 16 July 2021).
- Dang, T.T.; Pham, M.H.; Bui, H.V.; Van Le, D. First Full-Length Genome Sequence of Dengue Virus Serotype 2 Circulating in Vietnam in 2017. Infect. Drug Resist. 2020, 13, 4061–4068. [Google Scholar] [CrossRef] [PubMed]
- Rico-Hesse, R.; Harrison, L.M.; Salas, R.A.; Tovarb, D.; Nisalakc, A.; Ramosd, C.; Boshelle, J.; de Mesa, M.T.R.; Nogueira, R.M.; da Rosa, A.T. Origins of Dengue Type 2 Viruses Associated with Increased Pathogenicity in the Americas. Virology 1997, 230, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Cui, G.; Fang, D.; Yan, H.; Zhou, J.; Si, L.; Wu, D.; Jiang, L. Origin and Phylogenetic Characteristics of Dengue Virus Isolated from the Large Outbreak of Dengue in Guangdong Province in 2014. J. Sun Yat-sen Univ. Sci. 2017, 38, 21–28. [Google Scholar]
- Finol, E. Are viral small RNA regulating Dengue virus replication beyond serotype 2? Proc. Natl. Acad. Sci. USA 2014, 111, E2915–E2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkataraman, S.; Prasad, B.V.L.S.; Selvarajan, R. RNA Dependent RNA Polymerases: Insights from Structure, Function and Evolution. Viruses 2018, 10, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
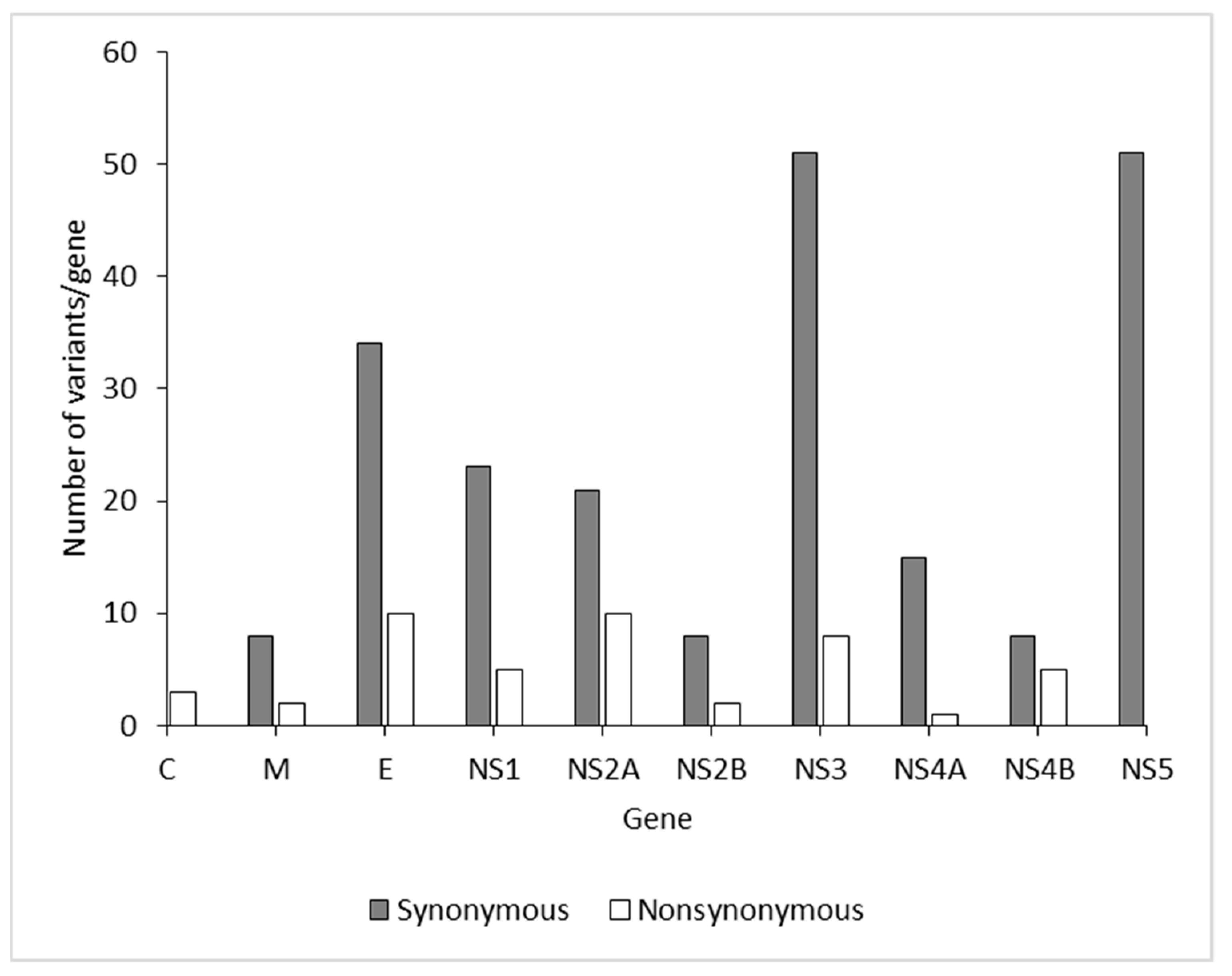{kind=link}
| Sample No. | Patient Code | Location | Age | Sex | Days of Fever | IgM Ratio | IgG Titer | Serum Viremia Level (log10 Copies/mL) | Virus Isolation Serotype |
|---|---|---|---|---|---|---|---|---|---|
| 1 | B-102 | Bardibas | 12 | M | 2 | 0.9 | 178 | 8.0 | DENV-2 |
| 2 | B-107 | Bardibas | 62 | M | 2 | 0.7 | 324 | 7.0 | DENV-2 |
| 3 | B-57 | Bardibas | 5 | F | N/A | 1.0 | 282 | 7.6 | DENV-2 |
| 4 | B-151 | Bardibas | 36 | F | N/A | 2.3 | 13,036 | 5.7 | DENV-2 |
| 5 | B-156 | Bardibas | N/A | N/A | N/A | 5.0 | 1702 | 4.9 | DENV-2 |
| 6 | B-157 | Bardibas | N/A | N/A | N/A | 0.8 | 1674 | 5.7 | DENV-2 |
| 7 | B-158 | Bardibas | N/A | N/A | N/A | 1.0 | 278 | 5.9 | DENV-2 |
| 8 | B-159 | Bardibas | N/A | N/A | N/A | 1.7 | 203 | 4.9 | DENV-2 |
| 9 | B-160 | Bardibas | N/A | N/A | N/A | 3.2 | 117 | 9.2 | DENV-2 |
| 10 | B-62 | Bardibas | 35 | M | N/A | 0.7 | 212 | 6.8 | DENV-2 |
| 11 | B-63 | Bardibas | 20 | M | N/A | 1.0 | 404 | 7.5 | DENV-2 |
| 12 | B-64 | Bardibas | 32 | F | N/A | 0.7 | 849 | 7.6 | DENV-2 |
| 13 | B-80 | Bardibas | 22 | F | 3 | 1.5 | 223 | 5.6 | DENV-2 |
| 14 | B-81 | Bardibas | 22 | M | 2 | 0.8 | 200 | 7.3 | DENV-2 |
| 15 | B-86 | Bardibas | 50 | F | N/A | 6.5 | 6112 | 7.3 | DENV-2 |
| 16 | B-97 | Bardibas | 48 | M | 2 | 0.9 | 395 | 7.4 | DENV-2 |
| 17 | D-13 | Nilkantha | 40 | M | 5 | 0.2 | 217 | 8.7 | DENV-2 |
| 18 | D-17 | Nilkantha | 21 | M | 2 | 0.6 | 219 | 6.1 | DENV-2 |
| 19 | D-24 | Nilkantha | 42 | F | 3 | 0.5 | 305 | 4.1 | DENV-2 |
| 20 | D-3 | Nilkantha | 15 | F | 3 | 2.2 | 135 | 4.5 | DENV-2 |
| 21 | D-30 | Nilkantha | 20 | F | 3 | 0.6 | 409 | 5.0 | DENV-2 |
| 22 | D-40 | Nilkantha | 30 | F | 3 | 0.3 | 342 | 5.8 | DENV-2 |
| 23 | L-10 | Lalbandi | 46 | F | 5 | 3.9 | 997 | 5.5 | DENV-2 |
| 24 | L-13 | Hariwon | 26 | M | 7 | 2.9 | 23,375 | 5.5 | DENV-2 |
| 25 | L-15 | Lalbandi | 30 | F | 3 | 6.8 | 9629 | 7.7 | DENV-2 |
| 26 | L-21 | Lalbandi | 18 | M | 5 | 9.4 | 221 | 5.5 | DENV-2 |
| 27 | L-3 | Lalbandi | 30 | F | 5 | 25.6 | 28,016 | 7.0 | DENV-2 |
| Sample No. | Feature | Nucleotide Position | Amino Acid Position | Reference Allele | Alternate Allele | Frequency (%) | Amino Acid Change |
|---|---|---|---|---|---|---|---|
| 20 | C | 418 | 108 | C | A | 7 | L-M |
| 20 | C | 431 | 112 | C | T | 7 | A-V |
| 20 | C | 435 | 113 | A | G | 7 | I-M |
| 1, 2, 3, 4, 5, 6, 7, 8, 9, 11, 13, 14, 17, 18, 19, 23, 24, 25, 26 | M | 704 | 203 | A | G | 22–62 | E-G |
| 21 | M | 817 | 241 | A | G | 1 | I-V |
| 23 | E | 954 | 286 | A | G | 52 | I-M |
| 17, 19 | E | 1046 | 317 | C | A | 28–55 | T-N |
| 9 | E | 1481 | 462 | C | T | 5 | T-M |
| 27 | E | 1649 | 518 | T | A | 1 | V-D |
| 12 | E | 1688 | 531 | T | C | 14 | V-A |
| 12 | E | 1691 | 532 | T | C | 13 | V-A |
| 27 | E | 1960 | 622 | T | A | 1 | L-M |
| 5, 6, 9, 10, 12, 14, 16, 24, 25, 26 | E | 1964 | 623 | A | G | 22–100 | E-G |
| 26 | E | 2009 | 638 | T | C | 13 | V-A |
| 20 | E | 2386 | 764 | G | A | 6 | V-I |
| 20 | NS1 | 2452 | 786 | A | G | 1 | K-E |
| 15 | NS1 | 2482 | 796 | A | G | 1 | I-V |
| 16 | NS1 | 2726 | 877 | G | A | 6 | R-Q |
| 20 | NS1 | 3236 | 1047 | A | G | 7 | K-R |
| 20 | NS1 | 3289 | 1065 | A | G | 12 | N-D |
| 21, 20 | NS2A | 3574 | 1160 | T | A | 20–31 | L-I |
| 21, 20 | NS2A | 3598 | 1168 | A | G | 36–37 | M-V |
| 20 | NS2A | 3598 | 1168 | G | A | 36 | V-M |
| 20 | NS2A | 3664 | 1190 | G | A | 22 | A-T |
| 9 | NS2A | 3736 | 1214 | T | C | 6 | F-L |
| 12 | NS2A | 3799 | 1235 | A | G | 10 | I-V |
| 23 | NS2A | 3803 | 1236 | T | C | 16 | V-A |
| 4 | NS2A | 3886 | 1264 | G | A | 100 | V-I |
| 19 | NS2A | 4126 | 1344 | A | G | 1 | K-E |
| 20 | NS2A | 4127 | 1344 | A | G | 1 | K-R |
| 21 | NS2B | 4387 | 1431 | A | G | 1 | I-V |
| 15 | NS2B | 4439 | 1448 | T | G | 1 | L-R |
| 20 | NS3 | 4612 | 1506 | C | T | 11 | L-F |
| 11, 17, 21 | NS3 | 4857 | 1587 | A | C | 1–21 | K-N |
| 11, 17, 19, 21 | NS3 | 4859 | 1588 | C | A | 1–17 | P-H |
| 15 | NS3 | 4954 | 1620 | A | G | 1 | K-E |
| 8 | NS3 | 5032 | 1646 | G | A | 97 | G-S |
| 20 | NS3 | 5078 | 1661 | G | A | 13 | R-K |
| 20 | NS3 | 5902 | 1936 | A | G | 5 | I-V |
| 27 | NS3 | 6071 | 1992 | A | G | 50 | D-G |
| 10, 12, 23 | NS4A | 6712 | 2206 | T | C | 1 | F-L |
| 19, 20 | NS4B | 6845 | 2250 | A | G | 25–27 | E-G |
| 18 | NS4B | 6850 | 2252 | C | A | 37 | P-T |
| 7 | NS4B | 6853 | 2253 | A | G | 10 | K-E |
| 18 | NS4B | 7252 | 2386 | A | G | 4 | K-E |
| 4, 5, 24 | NS4B | 7256 | 2387 | C | A | 90–97 | T-N |
| 15 | NS5 | 7609 | 2505 | A | C | 1 | K-Q |
| 12 | NS5 | 7966 | 2624 | T | C | 20 | F-L |
| 27 | NS5 | 8107 | 2671 | A | G | 1 | I-V |
| 20 | NS5 | 8423 | 2776 | G | A | 100 | R-K |
| 20 | NS5 | 8500 | 2802 | A | G | 1 | T-A |
| 8 | NS5 | 8731 | 2879 | A | G | 13 | K-E |
| 27 | NS5 | 8888 | 2931 | A | G | 1 | N-S |
| 18 | NS5 | 8947 | 2951 | A | G | 33 | K-E |
| 2 | NS5 | 9157 | 3021 | A | G | 5 | M-V |
| 21, 20, 27 | NS5 | 9210 | 3038 | A | T | 1–100 | L-F |
| 3 | NS5 | 9503 | 3136 | C | A | 63 | T-N |
| 3 | NS5 | 9518 | 3141 | T | C | 24 | V-A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngwe Tun, M.M.; Pandey, K.; Nabeshima, T.; Kyaw, A.K.; Adhikari, M.; Raini, S.K.; Inoue, S.; Dumre, S.P.; Pandey, B.D.; Morita, K. An Outbreak of Dengue Virus Serotype 2 Cosmopolitan Genotype in Nepal, 2017. Viruses 2021, 13, 1444. https://doi.org/10.3390/v13081444
Ngwe Tun MM, Pandey K, Nabeshima T, Kyaw AK, Adhikari M, Raini SK, Inoue S, Dumre SP, Pandey BD, Morita K. An Outbreak of Dengue Virus Serotype 2 Cosmopolitan Genotype in Nepal, 2017. Viruses. 2021; 13(8):1444. https://doi.org/10.3390/v13081444
Chicago/Turabian StyleNgwe Tun, Mya Myat, Kishor Pandey, Takeshi Nabeshima, Aung Kyaw Kyaw, Mandira Adhikari, Sandra Kendra Raini, Shingo Inoue, Shyam Prakash Dumre, Basu Dev Pandey, and Kouichi Morita. 2021. "An Outbreak of Dengue Virus Serotype 2 Cosmopolitan Genotype in Nepal, 2017" Viruses 13, no. 8: 1444. https://doi.org/10.3390/v13081444
APA StyleNgwe Tun, M. M., Pandey, K., Nabeshima, T., Kyaw, A. K., Adhikari, M., Raini, S. K., Inoue, S., Dumre, S. P., Pandey, B. D., & Morita, K. (2021). An Outbreak of Dengue Virus Serotype 2 Cosmopolitan Genotype in Nepal, 2017. Viruses, 13(8), 1444. https://doi.org/10.3390/v13081444