
Uniqueness of RNA Coliphage Qβ Display System in Directed Evolutionary Biotechnology

 ,
, 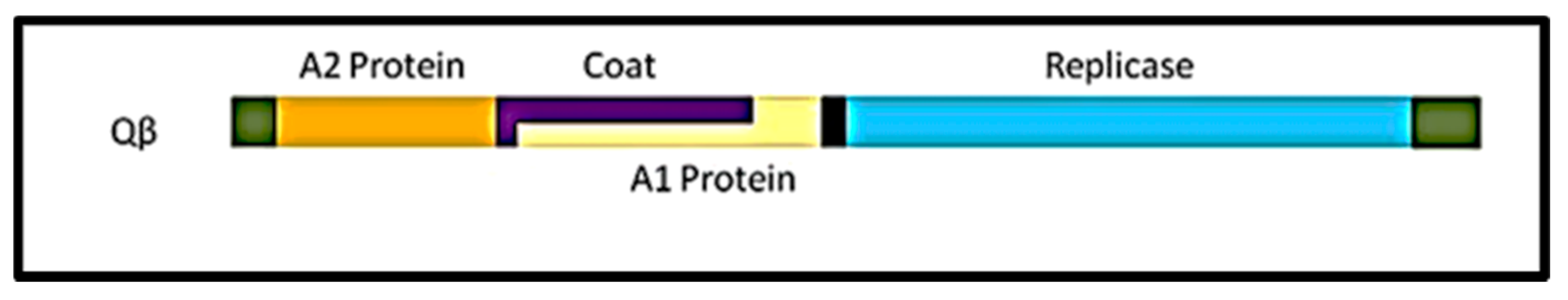{kind=link}
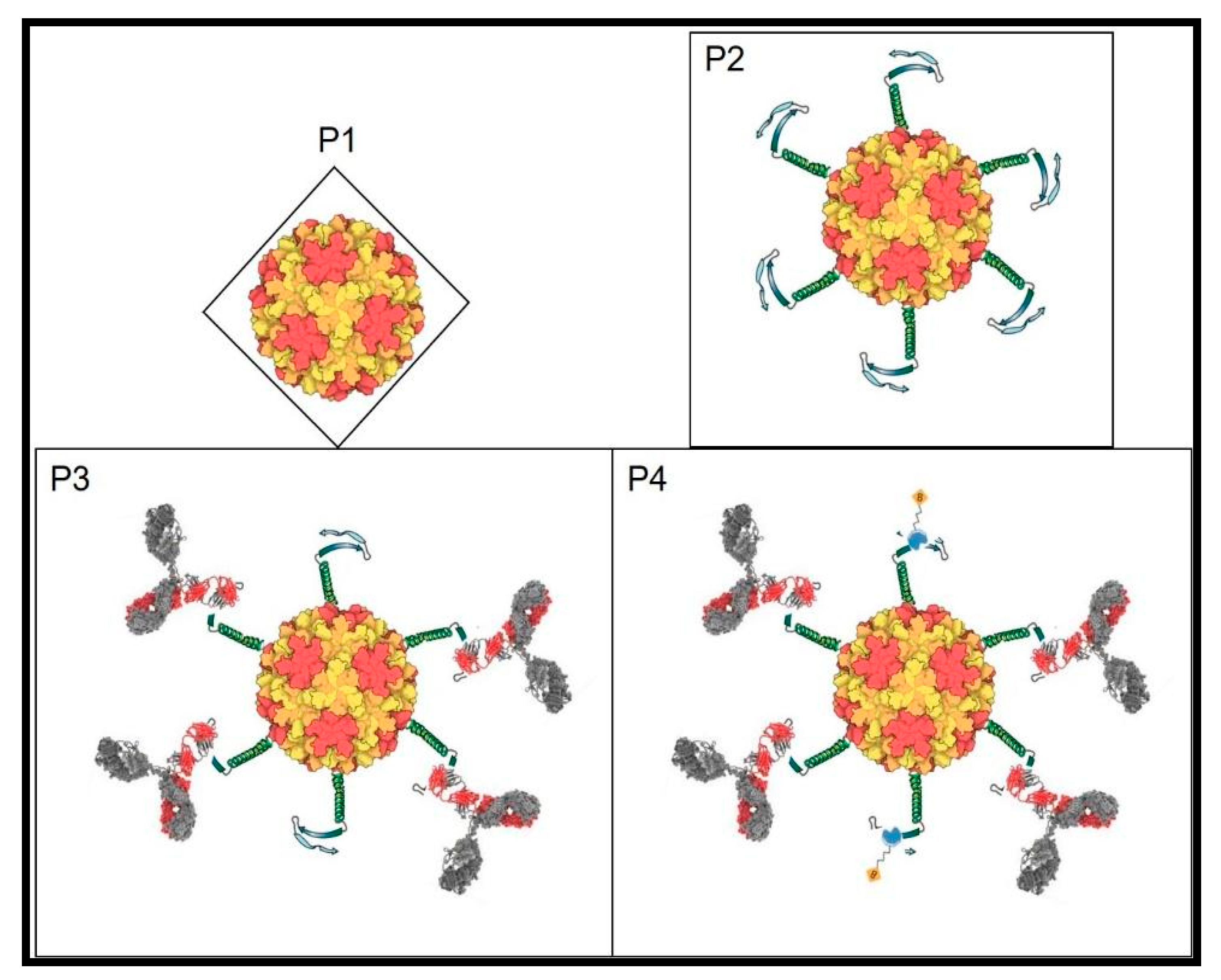{kind=link}
Abstract
1. Introduction
2. RNA Evolution
2.1. Overall Evolution
2.2. Evolution with RNA Viruses
2.3. An Emphasis of Evolution with Coliphage Qβ
3. Uniqueness of the Biology Coliphage Qβ
3.1. Coliphage Qβ Life Cycle
3.1.1. Overall Cycle
3.1.2. Proteins Involved in the Life Cycle and Functions
3.1.3. Unknown or Unclear Part of the Cycle
3.2. Coliphage Qβ A2 Protein
3.2.1. Structure and Domain
3.2.2. Functional Domains
3.2.3. Contribution in Panning for Phage Display
3.3. Coliphage Qβ Coat Protein
3.3.1. Structure and Domain
3.3.2. Function of Domain
3.3.3. Contribution to the RNA Display System
3.4. Coliphage Qβ Read-Through or A1 Protein
3.4.1. Structure and Domain
3.4.2. Functional Domains
3.4.3. Contribution to the Qβ Phage Display Platform
3.5. Coliphage Qβ Replicase Protein
3.5.1. Structure
3.5.2. Subunit Definition
3.5.3. Contribution to Evolution and Phage Display
4. Coliphage Qβ Display System
4.1. Overall Methodology: Features and Emphasis
4.2. Determining Steps in the RNA Qβ Display System
5. Application in Directed Evolution and Conclusion
Application
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef]
- Smith, G.P.; Petrenko, V.A. Phage display. Chem. Rev. 1997, 97, 391–410. [Google Scholar] [CrossRef]
- Fernandez-Gacio, A.; Uguen, M.; Fastrez, J. Phage display as a tool for the directed evolution of enzymes. Trends Biotechnol. 2003, 21, 408–414. [Google Scholar] [CrossRef]
- Citorik, R.J.; Mimee, M.; Lu, T.K. Bacteriophage-based synthetic biology for the study of infectious diseases. Curr. Opin. Microbiol. 2014, 19, 59–69. [Google Scholar] [CrossRef]
- Barbu, E.M.; Cady, K.C.; Hubby, B. Phage therapy in the era of synthetic biology. Cold Spring Harb. Perspect. Biol. 2016, 8, a023879. [Google Scholar] [CrossRef]
- Bertani, G. Lysogenic versus lytic cycle of phage multiplication. In Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1953; Volume 18, pp. 65–70. [Google Scholar]
- Wang, N.; Dykhuizen, D.E.; Slobodkin, L.B. The evolution of phage lysis timing. Evol. Ecol. 1996, 10, 545–558. [Google Scholar] [CrossRef]
- Chibani-Chennoufi, S.; Bruttin, A.; Dillmann, M.L.; Brüssow, H. Phage-host interaction: An ecological perspective. J. Bacteriol. 2004, 186, 3677–3686. [Google Scholar] [CrossRef]
- Böttger, V.; Böttger, A.; Howard, S.F.; Picksley, S.M.; Chène, P.; Garcia-Echeverria, C.; Hochkeppel, H.K.; Lane, D.P. Identification of novel mdm2 binding peptides by phage display. Oncogene 1996, 13, 2141–2147. [Google Scholar]
- Koivunen, E.; Arap, W.; Rajotte, D.; Lahdenranta, J.; Pasqualini, R. Identification of receptor ligands with phage display peptide libraries. J. Nucl. Med. 1999, 40, 883–888. [Google Scholar]
- Li, Y.; Moysey, R.; Molloy, P.E.; Vuidepot, A.L.; Mahon, T.; Baston, E.; Dunn, S.; Liddy, N.; Jacob, J.; Jakobsen, B.K.; et al. Directed evolution of human T-cell receptors with picomolar affinities by phage display. Nat. Biotechnol. 2005, 23, 349–354. [Google Scholar] [CrossRef]
- Bratkovič, T. Progress in phage display: Evolution of the technique and its applications. Cell. Mol. Life Sci. 2010, 67, 749–767. [Google Scholar] [CrossRef]
- Blanco, L.; Bernad, A.; Lázaro, J.M.; Martín, G.; Garmendia, C.; Salas, M. Highly efficient DNA synthesis by the phage ϕ 29 DNA polymerase: Symmetrical mode of DNA replication. J. Biol. Chem. 1989, 264, 8935–8940. [Google Scholar] [CrossRef]
- Cowart, M.; Gibson, K.J.; Allen, D.J.; Benkovic, S.J. DNA substrate structural requirements for the exonuclease and polymerase activities of prokaryotic and phage DNA polymerases. Biochemistry 1989, 28, 1975–1983. [Google Scholar] [CrossRef]
- Landers, T.A.; Blumenthal, T.; Weber, K. Function and structure in ribonucleic acid phage Qβ ribonucleic acid replicase: The roles of the different subunits in transcription of synthetic templates. J. Biol. Chem. 1974, 249, 5801–5808. [Google Scholar] [CrossRef]
- Young, R.A.; Blumenthal, T. Phage Q-beta ribonucleic acid replicase. Subunit relationships determined by intramolecular cross-linking. J. Biol. Chem. 1975, 250, 1829–1832. [Google Scholar] [CrossRef]
- Garmendia, C.; Bernad, A.; Esteban, J.A.; Blanco, L.; Salas, M. The bacteriophage phi 29 DNA polymerase, a proofreading enzyme. J. Biol. Chem. 1992, 267, 2594–2599. [Google Scholar] [CrossRef]
- Reddy, M.K.; Weitzel, S.E.; Von Hippel, P.H. Processive proofreading is intrinsic to T4 DNA polymerase. J. Biol. Chem. 1992, 267, 14157–14166. [Google Scholar] [CrossRef]
- Inokuchi, Y.; Hirashima, A.K. Interference with viral infection by defective RNA replicase. J. Virol. 1987, 61, 3946–3949. [Google Scholar] [CrossRef]
- Kajitani, M.; Ishihama, A. Identification and sequence determination of the host factor gene for bacteriophage Qβ. Nucleic Acids Res. 1991, 19, 1063–1066. [Google Scholar] [CrossRef]
- d’Herelle, F.H. Comptes Rendu. Acad. Sci. Paris 1917, 165, 373–375. [Google Scholar]
- d’Herelle, M.F. Sur un microbe invisible antagoniste des bacilles dysentériques. Acta Kravsi 1961, 1, 1. [Google Scholar]
- Stent, G.S. Molecular Biology of Bacterial Viruses. Available online: https://www.cabdirect.org/cabdirect/abstract/19640101695 (accessed on 18 February 2021).
- Summers, W.C. The strange history of phage therapy. Bacteriophage 2012, 2, 130–133. [Google Scholar] [CrossRef]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef]
- Salmond, G.P.; Fineran, P.C. A century of the phage: Past, present and future. Nat. Rev. Microbiol. 2015, 13, 777–786. [Google Scholar] [CrossRef]
- Chibani, C.M.; Farr, A.; Klama, S.; Dietrich, S.; Liesegang, H. Classifying the unclassified: A phage classification method. Viruses 2019, 11, 195. [Google Scholar] [CrossRef] [PubMed]
- Moineau, S.; Borkaev, M.; Holler, B.J.; Walker, S.A.; Kondo, J.K.; Vedamuthu, E.R.; Vandenbergh, P.A. Isolation and characterization of lactococcal bacteriophages from cultured buttermilk plants in the United States. J. Dairy Sci. 1996, 79, 2104–2111. [Google Scholar] [CrossRef]
- Ackermann, H.W. Phage classification and characterization. In Bacteriophages; Humana Press: Totowa, NJ, USA, 2009; pp. 127–140. [Google Scholar]
- Salivar, W.O.; Tzagoloff, H.; Pratt, D. Some physical-chemical and biological properties of the rod-shaped coliphage M13. Virology 1964, 24, 359–371. [Google Scholar] [CrossRef]
- Tzagoloff, H.; Pratt, D. The initial steps in infection with coliphage M13. Virology 1964, 24, 372–380. [Google Scholar] [CrossRef]
- Messing, J.; Gronenborn, B.; Müller-Hill, B.; Hopschneider, P.H. Filamentous coliphage M13 as a cloning vehicle: Insertion of a HindII fragment of the lac regulatory region in M13 replicative form in vitro. Proc. Natl. Acad. Sci. USA 1977, 74, 3642–3646. [Google Scholar] [CrossRef]
- Messing, J. New M13 vectors for cloning. Methods Enzymol. 1983, 101, 20–78. [Google Scholar] [PubMed]
- Goldsmith, M.E.; Konigsberg, W.H. Adsorption protein of the bacteriophage fd: Isolation, molecular properties, and location in the virus. Biochemistry 1977, 16, 2686–2694. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; Webster, R.E. Morphogenesis of filamentous bacteriophage f1: Orientation of extrusion and production of polyphage. Virology 1983, 127, 177–193. [Google Scholar] [CrossRef]
- Rakonjac, J.; Model, P. Roles of pIII in filamentous phage assembly. J. Mol. Biol. 1998, 282, 25–41. [Google Scholar] [CrossRef]
- Rakonjac, J.; Feng, J.N.; Model, P. Filamentous phage are released from the bacterial membrane by a two-step mechanism involving a short C-terminal fragment of pIII. J. Mol. Biol. 1999, 289, 1253–1265. [Google Scholar] [CrossRef]
- Palasingam, K.; Shaklee, P.N. Reversion of Q beta RNA phage mutants by homologous RNA recombination. J. Virol. 1992, 66, 2435–2442. [Google Scholar] [CrossRef]
- Malpica, J.M.; Fraile, A.; Moreno, I.; Obies, C.I.; Drake, J.W.; García-Arenal, F. The rate and character of spontaneous mutation in an RNA virus. Genetics 2002, 162, 1505–1511. [Google Scholar]
- Moya, A.; Elena, S.F.; Bracho, A.; Miralles, R.; Barrio, E. The evolution of RNA viruses: A population genetics view. Proc. Natl. Acad. Sci. USA 2000, 97, 6967–6973. [Google Scholar] [CrossRef]
- Elena, S.F.; Bedhomme, S.; Carrasco, P.; Cuevas, J.M.; de la Iglesia, F.; Lafforgue, G.; Lalić, J.; Pròsper, À.; Tromas, N.; Zwart, M.P. The evolutionary genetics of emerging plant RNA viruses. Mol. Plant Microbe Interact. 2011, 24, 287–293. [Google Scholar] [CrossRef]
- Domingo, E.; Escarmís, C.; Sevilla, N.; Moya, A.; Elena, S.F.; Quer, J.; Novella, I.S.; Holland, J.J. Basic concepts in RNA virus evolution. FASEB J. 1996, 10, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C.; Wallace, A.R. Evolution by natural selection. Evol. Nat. Sel. 1958, 1, 1. [Google Scholar]
- Mills, D.R.; Peterson, R.L.; Spiegelman, S. An extracellular Darwinian experiment with a self-duplicating nucleic acid molecule. Proc. Natl. Acad. Sci. USA 1967, 58, 217. [Google Scholar] [CrossRef] [PubMed]
- Popper, K. Natural selection and the emergence of mind. Dialectica 1978, 32, 339–355. [Google Scholar] [CrossRef]
- Tsimring, L.S.; Levine, H.; Kessler, D.A. RNA virus evolution via a fitness-space model. Phys. Rev. Lett. 1996, 76, 4440. [Google Scholar] [CrossRef] [PubMed]
- Burch, C.L.; Chao, L. Evolution by small steps and rugged landscapes in the RNA virus ϕ6. Genetics 1999, 151, 921–927. [Google Scholar]
- Domingo, E.; Holland, J.J. RNA virus mutations and fitness for survival. Annu. Rev. Microbiol. 1997, 51, 151–178. [Google Scholar] [CrossRef]
- Eigen, M. On the nature of virus quasispecies. Trends Microbiol. 1996, 4, 216–218. [Google Scholar] [CrossRef]
- Eigen, M. Viral quasispecies. Sci. Am. 1993, 269, 42–49. [Google Scholar] [CrossRef]
- Domingo, E. Quasispecies theory in virology. J. Virol. 2002, 76, 463–465. [Google Scholar] [CrossRef]
- Solé, R.V.; Ferrer, R.; González-García, I.; Quer, J.; Domingo, E. Red queen dynamics, competition and critical points in a model of RNA virus quasispecies. J. Theor. Biol. 1999, 198, 47–59. [Google Scholar] [CrossRef]
- Domingo, E.; Holland, J.J.; Biebricher, C.; Eigen, M. Quasispecies: The concept and the word. Mol. Basis Virus Evol. 1995, 1, 171–180. [Google Scholar]
- Waffo, A.B. Evolutionary RNA-coliphage Qβ [Q-beta] Display: Expression Biopathway in E. Coli; Cuvillier Verlag: Goettingen, Germany, 2004. [Google Scholar]
- Skamel, C.; Aller, S.G.; Waffo, A.B. In vitro evolution and affinity-maturation with coliphage Qβ display. PLoS ONE 2014, 9, e113069. [Google Scholar] [CrossRef]
- Källberg, M.; Wang, H.; Wang, S.; Peng, J.; Wang, Z.; Lu, H.; Xu, J. Template-based protein structure modeling using the RaptorX web server. Nature Protocols 2012, 7, 1511–1522. [Google Scholar] [CrossRef]
- Lissom, A.; Ouambo, H.F.; Megnekou, R.; Okeke, M.I.; Ngu, L.N.; Netongo, P.M.; Ngoh, A.A.; Sanders, C.A.; Bawage, S.; Tchouangueu, T.F.; et al. Comparative analysis of igg responses to recombinant Qβ phage displayed MSP3 and UB05 in dual HIV-malaria infected adults living in areas differing in malaria transmission intensities. bioRxiv 2018. [Google Scholar] [CrossRef]
- Waffo, A.B.; Lissom, A.; Ouambo, H.F.; Ngu, L.N.; Ngoh, A.A. Surface Engineering of the RNA Coliphage Qβ to Display Plasmodium Falciparum Derived Asexual Blood Stage Antigens UB05 and Merozoite Surface Protein 3. Clin. Microbiol. 2018, 7, 2. [Google Scholar] [CrossRef]
- Priano, C.; Arora, R.; Butke, J.; Mills, D.R. A complete plasmid-based complementation system for RNA coliphage Qβ: Three proteins of bacteriophages Qβ (group III) and SP (group IV) can be interchanged. J. Mol. Biol. 1995, 249, 283–297. [Google Scholar] [CrossRef]
- Witherell, G.W.; Uhlenbeck, O.C. Specific RNA binding by Q. beta. coat protein. Biochemistry 1989, 28, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Gorzelnik, K.V.; Cui, Z.; Reed, C.A.; Jakana, J.; Young, R.; Zhang, J. Asymmetric cryo-EM structure of the canonical Allolevivirus Qβ reveals a single maturation protein and the genomic ssRNA in situ. Proc. Natl. Acad. Sci. USA 2016, 113, 11519–11524. [Google Scholar] [CrossRef] [PubMed]
- Crawford, E.M.; Gesteland, R.F. The adsorption of bacteriophage R-17. Virology 1964, 22, 165–167. [Google Scholar] [CrossRef]
- Poot, R.A.; Tsareva, N.V.; Boni, I.V.; Van Duin, J. RNA folding kinetics regulates translation of phage MS2 maturation gene. Proc. Natl. Acad. Sci. USA 1997, 94, 10110–10115. [Google Scholar] [CrossRef]
- Reed, C.A. Characterization of A2: The Lysis Protein of SsRNA Phage Qbeta. Ph.D. Thesis, Texas A & M University, College Station, TX, USA, 2012. [Google Scholar]
- Klovins, J.; Van Duin, J. A long-range pseudoknot in Qβ RNA is essential for replication. J. Mol. Biol. 1999, 294, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Rumnieks, J.; Tars, K. Crystal structure of the maturation protein from bacteriophage Qβ. J. Mol. Biol. 2017, 429, 688–696. [Google Scholar] [CrossRef]
- Rūmnieks, J.; Tārs, K. Protein-RNA interactions in the single-stranded RNA bacteriophages. Virus Protein Nucl. Complexes 2018, 88, 281–303. [Google Scholar]
- Tars, K. ssRNA Phages: Life Cycle, Structure and Applications. In Biocommunication of Phages; Springer: Cham, Switzerland, 2020; pp. 261–292. [Google Scholar]
- Reed, C.A.; Langlais, C.; Kuznetsov, V.; Young, R. Inhibitory mechanism of the Qβ lysis protein A2. Mol. Microbiol. 2012, 86, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, D.; Tomita, K. Assembly of Qβ viral RNA polymerase with host translational elongation factors EF-Tu and-Ts. Proc. Natl. Acad. Sci. USA 2010, 107, 15733–15738. [Google Scholar] [CrossRef]
- Morais, M.C. Breaking the symmetry of a viral capsid. Proc. Natl. Acad. Sci. USA 2016, 113, 11390–11392. [Google Scholar] [CrossRef]
- Chamakura, K.R.; Tran, J.S.; O’Leary, C.; Lisciandro, H.G.; Antillon, S.F.; Garza, K.D.; Tran, E.; Min, L.; Young, R. Rapid de novo evolution of lysis genes in single-stranded RNA phages. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Serwer, P.; Wright, E.T.; Demeler, B.; Jiang, W. States of phage T3/T7 capsids: Buoyant density centrifugation and cryo-EM. Biophys. Rev. 2018, 10, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Golmohammadi, R.; Fridborg, K.; Bundule, M.; Valegård, K.; Liljas, L. The crystal structure of bacteriophage Qβ at 3.5 Å resolution. Structure 1996, 4, 543–554. [Google Scholar] [CrossRef]
- Bernhardt, T.G.; Wang, N.; Struck, D.K.; Young, R. Breaking free:“protein antibiotics” and phage lysis. Res. Microbiol. 2002, 153, 493–501. [Google Scholar] [CrossRef]
- Karnik, S.; Billeter, M. The lysis function of RNA bacteriophage Qbeta is mediated by the maturation (A2) protein. EMBO J. 1983, 2, 1521–1526. [Google Scholar] [CrossRef]
- Mills, D.R.; Priano, C.; Merz, P.A.; Binderow, B.D. Q beta RNA bacteriophage: Mapping cis-acting elements within an RNA genome. J. Virol. 1990, 64, 3872–3881. [Google Scholar] [CrossRef]
- Taniguchi, T.; Palmieri, M.; Weissmann, C. Qβ DNA-containing hybrid plasmids giving rise to Qβ phage formation in the bacterial host. Nature 1978, 274, 223–228. [Google Scholar] [CrossRef]
- Klovins, J.; Berzins, V.; Van Duin, J. A long-range interaction in Qbeta RNA that bridges the thousand nucleotides between the M-site and the 3’end is required for replication. RNA 1998, 4, 948–957. [Google Scholar] [CrossRef]
- Lázaro, E.; Arribas, M.; Cabanillas, L.; Román, I.; Acosta, E. Evolutionary adaptation of an RNA bacteriophage to the simultaneous increase in the within-host and extracellular temperatures. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Stoll, E.; Wilson, K.J.; Reiser, J.; Weissmann, C. Revised amino acid sequence of Qbeta coat protein between positions 1 and 60. J. Biol. Chem. 1977, 252, 990–993. [Google Scholar] [CrossRef]
- Ni, C.Z.; White, C.A.; Mitchell, R.S.; Wickersham, J.; Kodandapani, R.; Peabody, D.S.; Ely, K.R. Crystal structure of the coat protein from the GA bacteriophage: Model of the unassembled dimer. Protein Sci. 1996, 5, 2485–2493. [Google Scholar] [CrossRef] [PubMed]
- Lim, F.; Peabody, D.S. RNA recognition site of PP7 coat protein. Nucleic Acids Res. 2002, 30, 4138–4144. [Google Scholar] [CrossRef] [PubMed]
- Shishovs, M.; Rumnieks, J.; Diebolder, C.; Jaudzems, K.; Andreas, L.B.; Stanek, J.; Kazaks, A.; Kotelovica, S.; Akopjana, I.; Pintacuda, G.; et al. Structure of AP205 coat protein reveals circular permutation in ssRNA bacteriophages. J. Mol. Biol. 2016, 428, 4267–4279. [Google Scholar] [CrossRef]
- Rumnieks, J.; Tars, K. Crystal structure of the read-through domain from bacteriophage Qβ A1 protein. Protein Sci. 2011, 20, 1707–1712. [Google Scholar] [CrossRef]
- Axelrod, V.D.; Brown, E.; Priano, C.; Mills, D.R. Coliphage Qβ RNA replication: RNA catalytic for single-strand release. Virology 1991, 184, 595–608. [Google Scholar] [CrossRef]
- Barrett, R.W.; Cwirla, S.E.; Ackerman, M.S.; Olson, A.M.; Peters, E.A.; Dower, W.J. Selective enrichment and characterization of high affinity ligands from collections of random peptides on filamentous phage. Anal. Biochem. 1992, 204, 357–364. [Google Scholar] [CrossRef]
- Adzhubei, A.A.; Sternberg, M.J.; Makarov, A.A. Polyproline-II helix in proteins: Structure and function. J. Mol. Biol. 2013, 425, 2100–2132. [Google Scholar] [CrossRef]
- Katz, M.J.; Gándara, L.; Ezcurra, A.D.L.; Wappner, P. Hydroxylation and translational adaptation to stress: Some answers lie beyond the STOP codon. Cell. Mol. Life Sci. 2016, 73, 1881–1893. [Google Scholar] [CrossRef]
- Brown, D.; Gold, L. RNA replication by Q beta replicase: A working model. Proc. Natl. Acad. Sci. USA 1996, 93, 11558–11562. [Google Scholar] [CrossRef]
- Bycroft, M.; Hubbard, T.J.; Proctor, M.; Freund, S.M.; Murzin, A.G. The solution structure of the S1 RNA binding domain: A member of an ancient nucleic acid–binding fold. Cell 1997, 88, 235–242. [Google Scholar] [CrossRef]
- Takeshita, D.; Tomita, K. Molecular basis for RNA polymerization by Qβ replicase. Nat. Struct. Mol. Biol. 2012, 19, 229. [Google Scholar] [CrossRef]
- Kashiwagi, A.; Yomo, T. Ongoing phenotypic and genomic changes in experimental coevolution of RNA bacteriophage Qβ and Escherichia coli. PLoS Genet. 2011, 7, e1002188. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, D.; Yamashita, S.; Tomita, K. Molecular insights into replication initiation by Qβ replicase using ribosomal protein S1. Nucleic Acids Res. 2014, 42, 10809–10822. [Google Scholar] [CrossRef] [PubMed]
- Biebricher, C.K.; Luce, R. In vitro recombination and terminal elongation of RNA by Q beta replicase. EMBO J. 1992, 11, 5129–5135. [Google Scholar] [CrossRef]
- Blumenthal, T.; Carmichael, G.G. RNA replication: Function and structure of Qbeta-replicase. Annu. Rev. Biochem. 1979, 48, 525–548. [Google Scholar] [CrossRef] [PubMed]
- Muffler, A.; Fischer, D.; Hengge-Aronis, R. The RNA-binding protein HF-I, known as a host factor for phage Qbeta RNA replication, is essential for rpoS translation in Escherichia coli. Genes Dev. 1996, 10, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Hajnsdorf, E.; Boni, I.V. Multiple activities of RNA-binding proteins S1 and Hfq. Biochimie 2012, 94, 1544–1553. [Google Scholar] [CrossRef] [PubMed]
- Sumper, M.; Luce, R. Evidence for de novo production of self-replicating and environmentally adapted RNA structures by bacteriophage Qbeta replicase. Proc. Natl. Acad. Sci. USA 1975, 72, 162–166. [Google Scholar] [CrossRef]
- Yoshinari, S.; Nagy, P.D.; Simon, A.E.; Dreher, T.W. CCA initiation boxes without unique promoter elements support in vitro transcription by three viral RNA-dependent RNA polymerases. RNA 2000, 6, 698–707. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bernhardt, T.G.; Wang, N.; Struck, D.K.; Young, R. A protein antibiotic in the phage Qβ virion: Diversity in lysis targets. Science 2001, 292, 2326–2329. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nchinda, G.W.; Al-Atoom, N.; Coats, M.T.; Cameron, J.M.; Waffo, A.B. Uniqueness of RNA Coliphage Qβ Display System in Directed Evolutionary Biotechnology. Viruses 2021, 13, 568. https://doi.org/10.3390/v13040568
Nchinda GW, Al-Atoom N, Coats MT, Cameron JM, Waffo AB. Uniqueness of RNA Coliphage Qβ Display System in Directed Evolutionary Biotechnology. Viruses. 2021; 13(4):568. https://doi.org/10.3390/v13040568
Chicago/Turabian StyleNchinda, Godwin W., Nadia Al-Atoom, Mamie T. Coats, Jacqueline M. Cameron, and Alain B. Waffo. 2021. "Uniqueness of RNA Coliphage Qβ Display System in Directed Evolutionary Biotechnology" Viruses 13, no. 4: 568. https://doi.org/10.3390/v13040568
APA StyleNchinda, G. W., Al-Atoom, N., Coats, M. T., Cameron, J. M., & Waffo, A. B. (2021). Uniqueness of RNA Coliphage Qβ Display System in Directed Evolutionary Biotechnology. Viruses, 13(4), 568. https://doi.org/10.3390/v13040568