
An Amplicon-Based Approach for the Whole-Genome Sequencing of Human Metapneumovirus

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
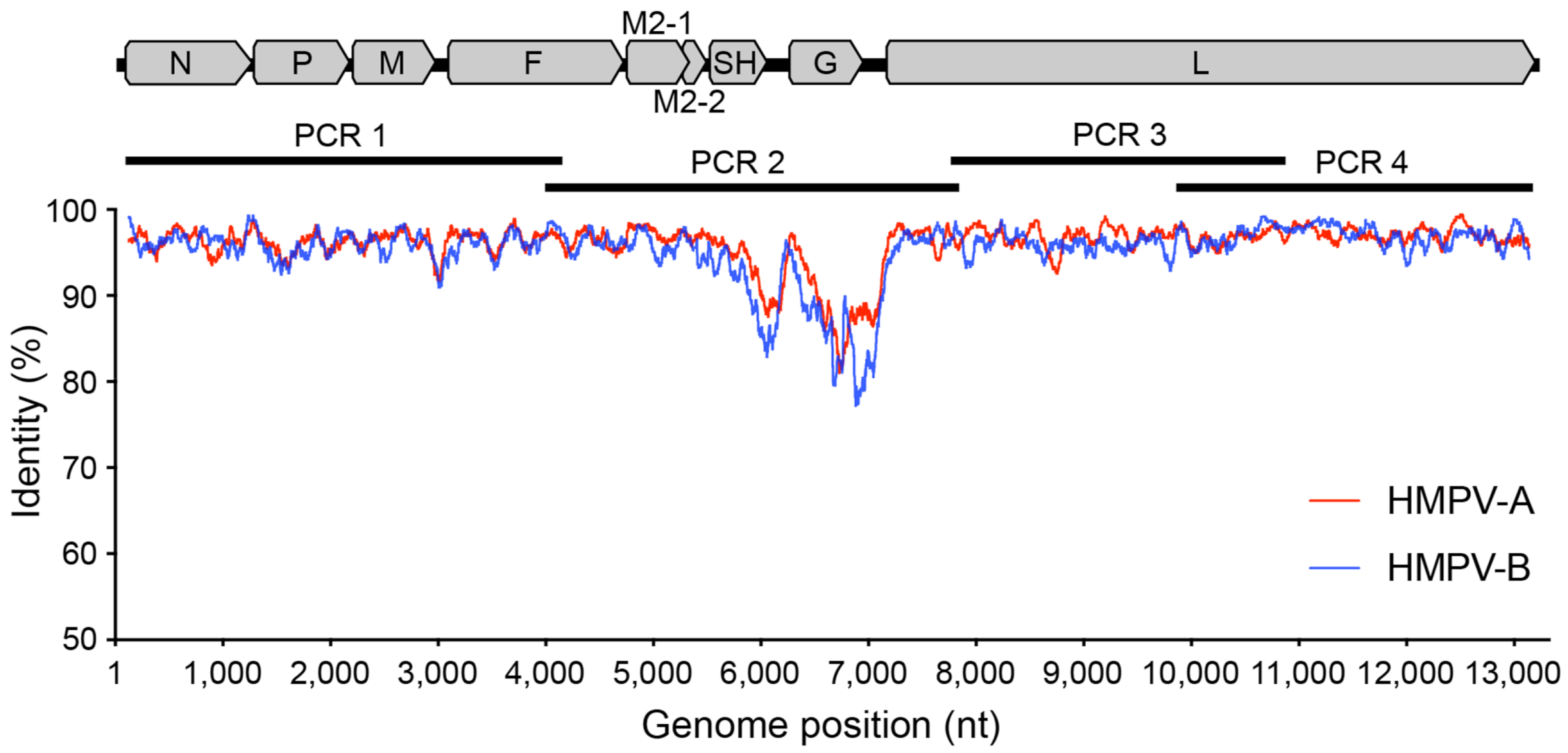
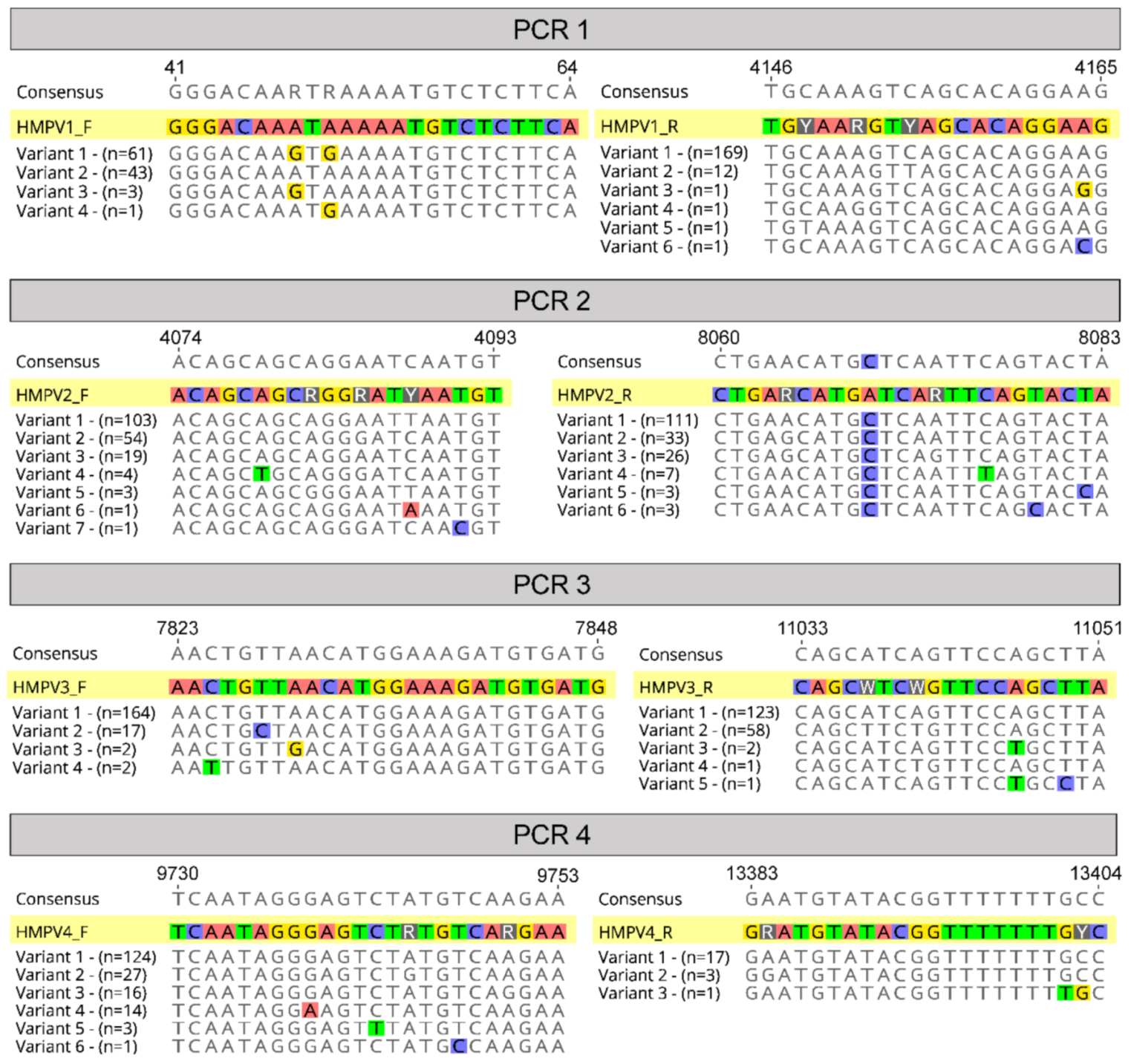
2.1. Primer Design
2.2. Clinical Specimens and HMPV Isolation
2.3. Reverse Transcription
2.4. HMPV Genome Amplification
2.5. Library Preparation and Sequencing
2.6. Viral Assembly
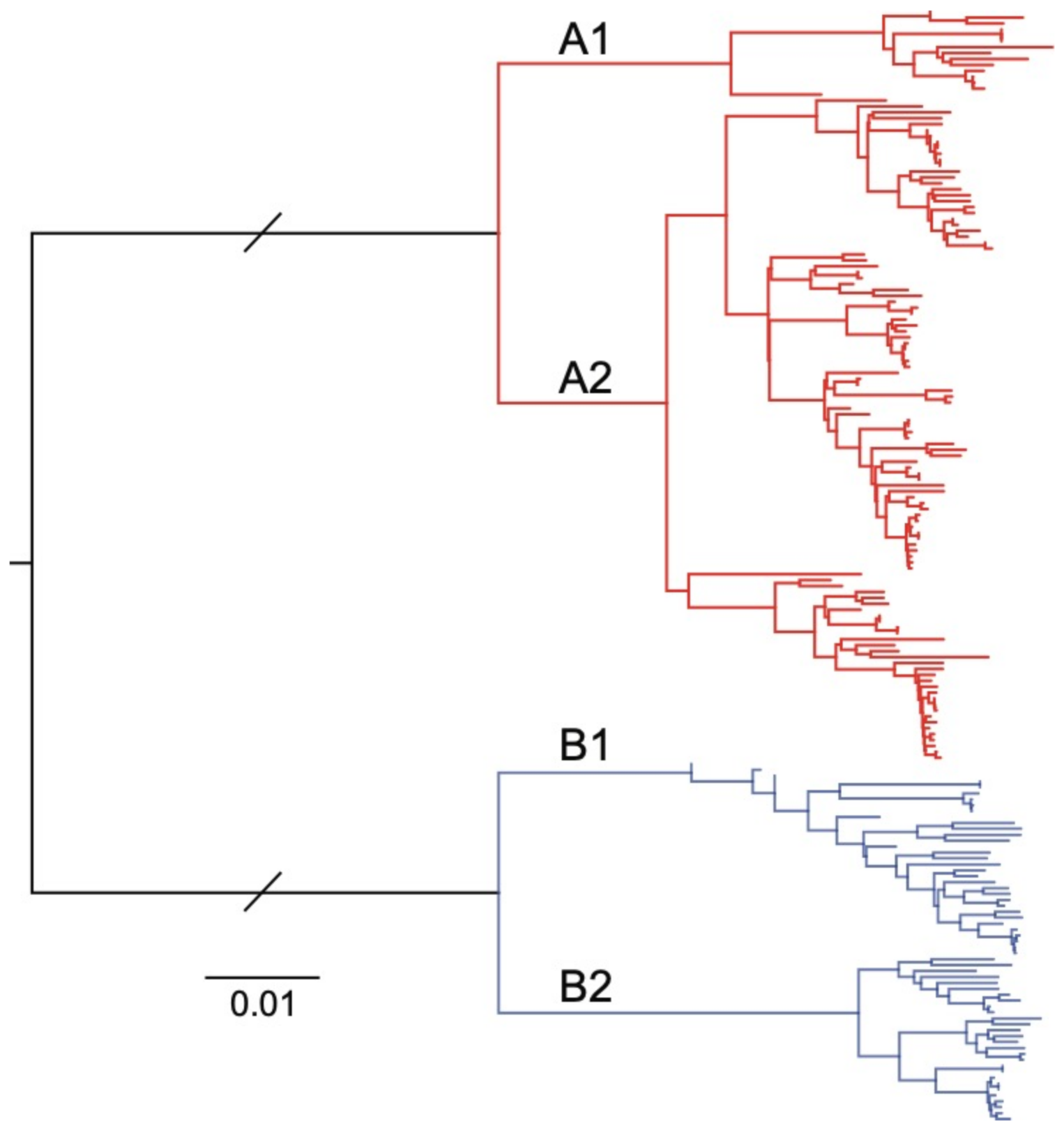
2.7. Phylogenetic Analysis
3. Results and Discussion
3.1. Designing Primers to Amplify the HMPV Genome
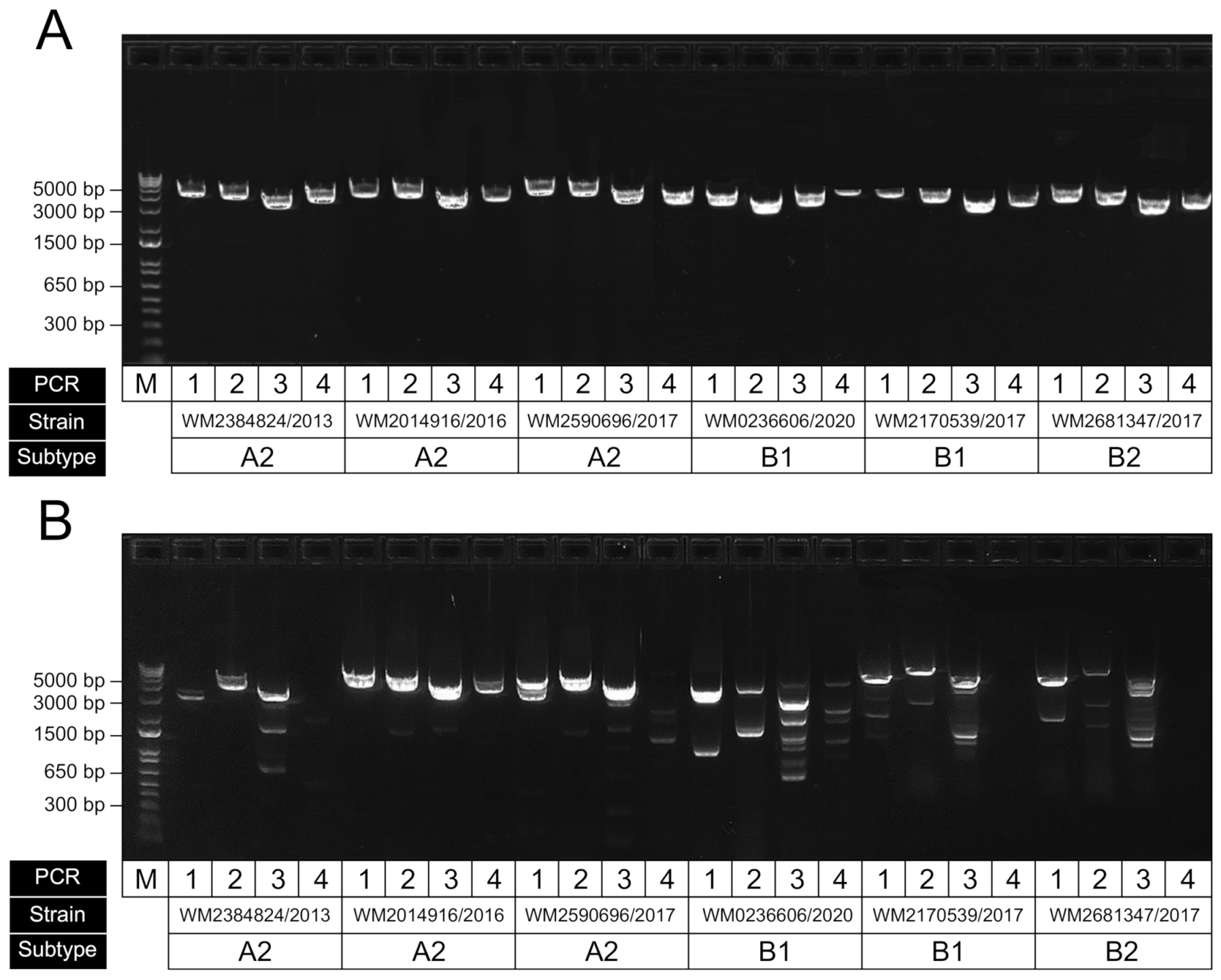
3.2. HMPV Genome RT-PCR Performance
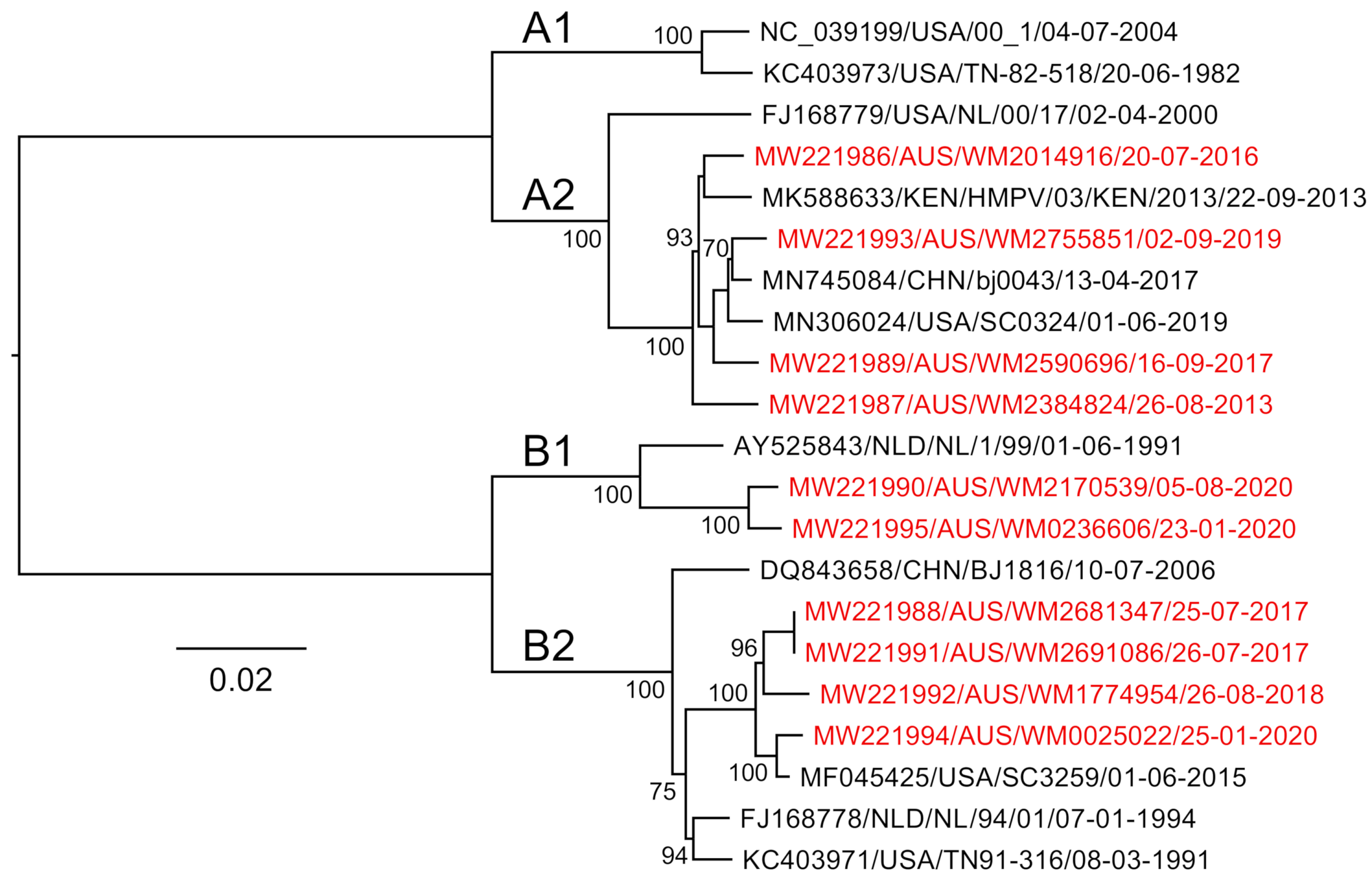
3.3. Genome Sequencing, Assembly and Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoogen, B.G.V.D.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Analysis of the Genomic Sequence of a Human Metapneumovirus. Virology 2002, 295, 119–132. [Google Scholar] [CrossRef]
- Kaslow, R.A.; Stanberry, L.R.; Le Duc, J.W. Viral Infections of Humans Epidemiology and Control, 5th ed.; Springer: Boston, MA, USA, 2014. [Google Scholar]
- Biacchesi, S.; Skiadopoulos, M.H.; Boivin, G.; Hanson, C.T.; Murphy, B.R.; Collins, P.L.; Buchholz, U.J. Genetic diversity between human metapneumovirus subgroups. Virology 2003, 315, 1–9. [Google Scholar] [CrossRef]
- Falsey, A.R.; Erdman, D.; Anderson, L.J.; Walsh, E.E. Human Metapneumovirus Infections in Young and Elderly Adults. J. Infect. Dis. 2003, 187, 785–790. [Google Scholar] [CrossRef]
- Hoogen, B.G.V.D.; De Jong, J.C.; Groen, J.; Kuiken, T.; De Groot, R.; Fouchier, R.A.; Osterhaus, A.D. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat. Med. 2001, 7, 719–724. [Google Scholar] [CrossRef]
- McAdam, A.J.; Hasenbein, M.E.; Feldman, H.A.; Cole, S.E.; Offermann, J.T.; Riley, A.M.; Lieu, T.A. Human Metapneumovirus in Children Tested at a Tertiary-Care Hospital. J. Infect. Dis. 2004, 190, 20–26. [Google Scholar] [CrossRef]
- Williams, J.V.; Harris, P.A.; Tollefson, S.J.; Halburnt-Rush, L.L.; Pingsterhaus, J.M.; Edwards, K.M.; Wright, P.F.; Crowe, J.E. Human Metapneumovirus and Lower Respiratory Tract Disease in Otherwise Healthy Infants and Children. N. Engl. J. Med. 2004, 350, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Widmer, K.; Zhu, Y.; Williams, J.V.; Griffin, M.R.; Edwards, K.M.; Talbot, H.K. Rates of Hospitalizations for Respiratory Syncytial Virus, Human Metapneumovirus, and Influenza Virus in Older Adults. J. Infect. Dis. 2012, 206, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, K.A.; Collins, P.L.; Peeples, M.E. Synergistic Effects of Gene-End Signal Mutations and the M2-1 Protein on Transcription Termination by Respiratory Syncytial Virus. Virology 2001, 288, 295–307. [Google Scholar] [CrossRef] [PubMed]
- El Najjar, F.; Cifuentes-Muñoz, N.; Chen, J.; Zhu, H.; Buchholz, U.J.; Moncman, C.L.; Dutch, R.E. Human metapneumovirus Induces Reorganization of the Actin Cytoskeleton for Direct Cell-to-Cell Spread. PLoS Pathog. 2016, 12, e1005922. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.K.; Kweon, O.J.; Kim, H.R.; Kim, T.-H.; Lee, M.-K. Clinical Features, Epidemiology, and Climatic Impact of Genotype-specific Human Metapneumovirus Infections: Long-term Surveillance of Hospitalized Patients in South Korea. Clin. Infect. Dis. 2019, 70, 2683–2694. [Google Scholar] [CrossRef]
- Jain, B.; Singh, A.K.; Dangi, T.; Agarwal, A.; Verma, A.K.; Dwivedi, M.; Singh, K.P.; Jain, A. High prevalence of human metapneumovirus subtype B in cases presenting as severe acute respiratory illness: An experience at tertiary care hospital. Clin. Respir. J. 2013, 8, 225–233. [Google Scholar] [CrossRef]
- Reiche, J.; Jacobsen, S.; Neubauer, K.; Hafemann, S.; Nitsche, A.; Milde, J.; Wolff, T.; Schweiger, B. Human Metapneumovirus: Insights from a Ten-Year Molecular and Epidemiological Analysis in Germany. PLoS ONE 2014, 9, e88342. [Google Scholar] [CrossRef]
- Gardy, J.L.; Naus, M.; Amlani, A.; Chung, W.; Kim, H.; Tan, M.; Severini, A.; Krajden, M.; Puddicombe, D.; Sahni, V.; et al. Whole-Genome Sequencing of Measles Virus Genotypes H1 and D8 During Outbreaks of Infection Following the 2010 Olympic Winter Games Reveals Viral Transmission Routes. J. Infect. Dis. 2015, 212, 1574–1578. [Google Scholar] [CrossRef] [PubMed]
- Houlihan, C.F.; Frampton, D.; Ferns, R.B.; Raffle, J.; Grant, P.; Reidy, M.; Hail, L.; Thomson, K.; Mattes, F.; Kozlakidis, Z.; et al. Use of Whole-Genome Sequencing in the Investigation of a Nosocomial Influenza Virus Outbreak. J. Infect. Dis. 2018, 218, 1485–1489. [Google Scholar] [CrossRef]
- Gymoese, P.; Sørensen, G.; Litrup, E.; Olsen, J.E.; Nielsen, E.M.; Torpdahl, M. Investigation of Outbreaks ofSalmonella entericaSerovar Typhimurium and Its Monophasic Variants Using Whole-Genome Sequencing, Denmark. Emerg. Infect. Dis. 2017, 23, 1631–1639. [Google Scholar] [CrossRef]
- Eden, J.-S.; Rockett, R.; Carter, I.; Rahman, H.; De Ligt, J.; Hadfield, J.; Storey, M.; Ren, X.; Tulloch, R.; Basile, K.; et al. An emergent clade of SARS-CoV-2 linked to returned travellers from Iran. Virus Evol. 2020, 6, veaa027. [Google Scholar] [CrossRef]
- Freed, N.E.; Vlková, M.; Faisal, M.B.; Silander, O.K. Rapid and Inexpensive Whole-Genome Sequencing of SARS-CoV-2 using 1200 bp Tiled Amplicons and Oxford Nanopore Rapid Barcoding. Biol. Methods Protoc. 2020, 5, bpaa014. [Google Scholar] [CrossRef] [PubMed]
- Tyson, J.R.; James, P.; Stoddart, D.; Sparks, N.; Wickenhagen, A.; Hall, G.; Choi, J.H.; Lapointe, H.; Kamelian, K.; Smith, A.D.; et al. Improvements to the ARTIC multiplex PCR method for SARS-CoV-2 genome sequencing us-ing nanopore. bioRxiv 2020. [Google Scholar] [CrossRef]
- Di Giallonardo, F.; Kok, J.; Fernandez, M.; Carter, I.; Geoghegan, J.L.; Dwyer, D.E.; Holmes, E.C.; Eden, J.-S. Evolution of Human Respiratory Syncytial Virus (RSV) over Multiple Seasons in New South Wales, Australia. Viruses 2018, 10, 476. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Chuang, L.-Y.; Cheng, Y.-H.; Yang, C.-H. Specific primer design for the polymerase chain reaction. Biotechnol. Lett. 2013, 35, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Pollett, S.; Trovão, N.S.; Tan, Y.; Eden, J.-S.; Halpin, R.A.; Bera, J.; Das, S.R.; Wentworth, D.; Ocaña, V.; Mendocilla, S.M.; et al. The transmission dynamics and diversity of human metapneumovirus in Peru. Influ. Other Respir. Viruses 2018, 12, 508–513. [Google Scholar] [CrossRef]
- Brown, J.W.; Pirrung, M.; McCue, L.A. FQC Dashboard: Integrates FastQC results into a web-based, interactive, and extensible FASTQ quality control tool. Bioinformatics 2017, 33, 3137–3139. [Google Scholar] [CrossRef] [PubMed]
- Bushnell. BBMap Short-Read Aligner, and Other Bioinformatics Tools. 2016. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 8 January 2020).
- Li, D.; Liu, C.-M.; Luo, R.; Sadakane, K.; Lam, T.-W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Da Silva, J.O.; Orellana, E.T.V.; Torres, M.; Delgado, M.X.T. Development of a Parallel Version of PhyML 3.0 Using Shared Memory. IEEE Lat. Am. Trans. 2017, 15, 959–967. [Google Scholar] [CrossRef]
- Cotten, M.; Petrova, V.; Phan, M.V.T.; Rabaa, M.A.; Watson, S.J.; Ong, S.H.; Kellam, P.; Baker, S. Deep Sequencing of Norovirus Genomes Defines Evolutionary Patterns in an Urban Tropical Setting. J. Virol. 2014, 88, 11056–11069. [Google Scholar] [CrossRef]
- Sloots, T.P.; Mackay, I.M.; Bialasiewicz, S.; Jacob, K.C.; McQueen, E.; Harnett, G.B.; Siebert, D.J.; Masters, B.I.; Young, P.R.; Nissen, M.D. Human metapneumovirus, Australia, 2001–2004. Emerg. Infect. Dis. 2006, 12, 1263–1266. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Zou, L.; Peng, J.; Yu, J.; Song, Y.; Liang, L.; Guo, Q.; Kang, M.; Ke, C.; Song, T.; et al. Epidemiology, evolution and transmission of human metapneumovirus in Guangzhou China, 2013–2017. Sci. Rep. 2019, 9, 14022. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Primer Name | Sequence (5’-3’) | Position (nt) * | PCR Amplicon Size (bp) |
|---|---|---|---|---|
| PCR1 | HMPV1_F | GGGACAAATAAAAATGTCTCTTCA | 41 | 4125 |
| HMPV1_R | CTTCCTGTGCTRACYTTRCA | 4165 | ||
| PCR2 | HMPV2_F | ACAGCAGCRGGRATYAATGT | 4074 | 4010 |
| HMPV2_R | TAGTACTGAAYTGAGCATGYTCAG | 8083 | ||
| PCR3 | HMPV3_F | AACTGTTAACATGGAAAGATGTGATG | 7823 | 3229 |
| HMPV3_R | TAAGCTGGAACWGAWGCTG | 11,051 | ||
| PCR4 | HMPV4_F | TCAATAGGGAGTCTRTGTCARGAA | 9730 | 3675 |
| HMPV4_R | GRCAAAAAAACCGTATACATYC | 13,404 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tulloch, R.L.; Kok, J.; Carter, I.; Dwyer, D.E.; Eden, J.-S. An Amplicon-Based Approach for the Whole-Genome Sequencing of Human Metapneumovirus. Viruses 2021, 13, 499. https://doi.org/10.3390/v13030499
Tulloch RL, Kok J, Carter I, Dwyer DE, Eden J-S. An Amplicon-Based Approach for the Whole-Genome Sequencing of Human Metapneumovirus. Viruses. 2021; 13(3):499. https://doi.org/10.3390/v13030499
Chicago/Turabian StyleTulloch, Rachel L., Jen Kok, Ian Carter, Dominic E. Dwyer, and John-Sebastian Eden. 2021. "An Amplicon-Based Approach for the Whole-Genome Sequencing of Human Metapneumovirus" Viruses 13, no. 3: 499. https://doi.org/10.3390/v13030499
APA StyleTulloch, R. L., Kok, J., Carter, I., Dwyer, D. E., & Eden, J.-S. (2021). An Amplicon-Based Approach for the Whole-Genome Sequencing of Human Metapneumovirus. Viruses, 13(3), 499. https://doi.org/10.3390/v13030499