
Molecular Characterization of Ahp2, a Lytic Bacteriophage of Aeromonas hydrophila


Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Phage Isolation and Host-Range Testing
2.3. Isolation of Phage-Resistant Mutants and Determination of the Lysogenic Activity of Ahp2
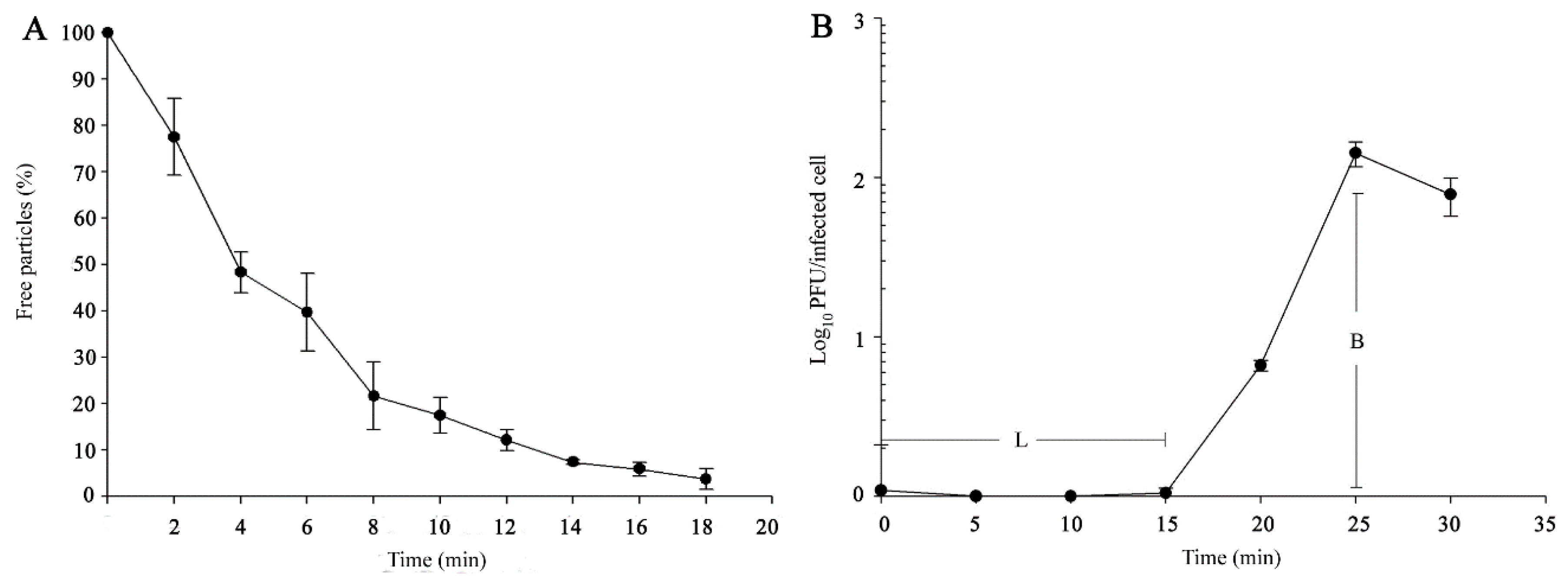
2.4. Phage Adsorption, One-Step Growth, and Purification
2.5. Phage DNA Isolation and Pulsed-Field Gel Electrophoresis
2.6. Protein Analyses
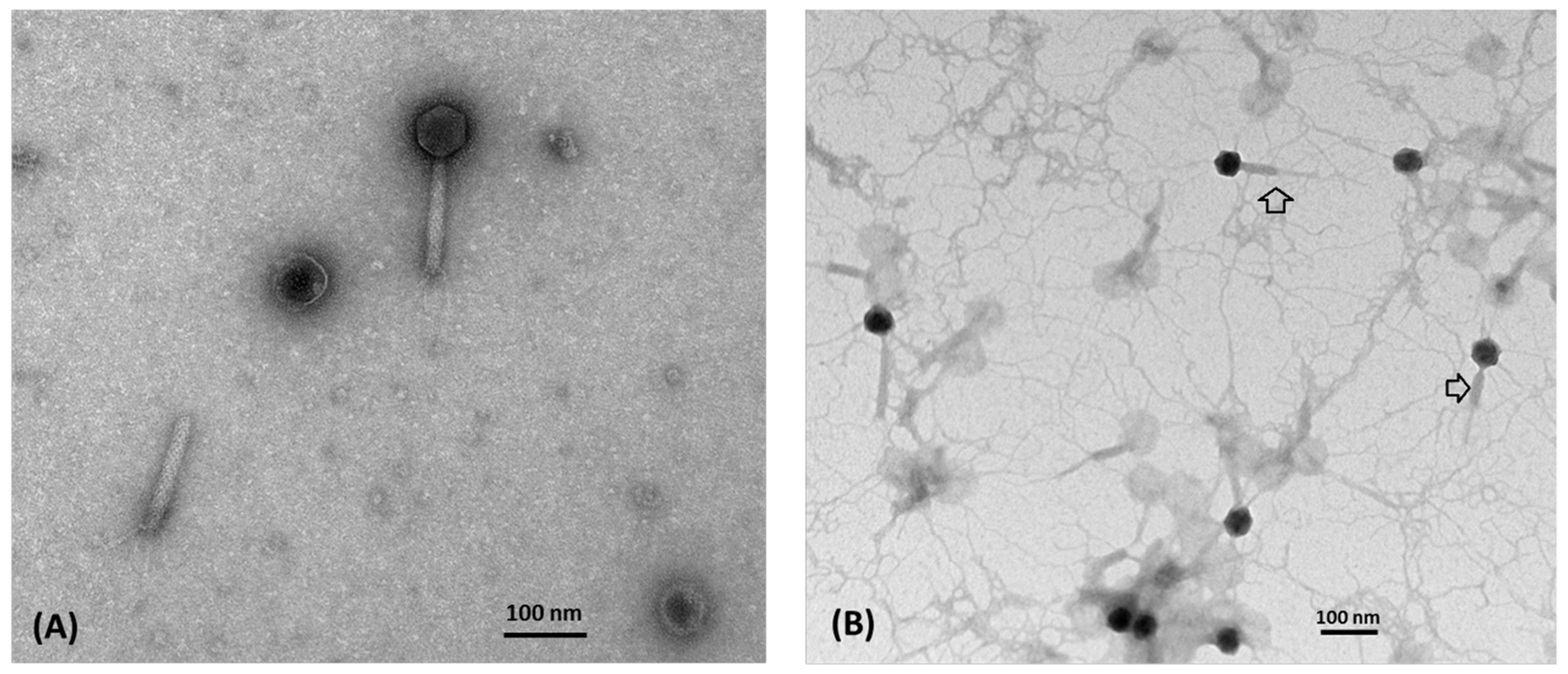
2.7. Electron Microscopy
2.8. Whole-Genome Sequencing and In Silico Analysis
2.9. Nucleotide Sequence Accession Number
3. Results
3.1. Morphology and Biological Properties of Ahp2
3.2. Ahp2 Infects a Wide Range of Environmental A. hydrophila Strains
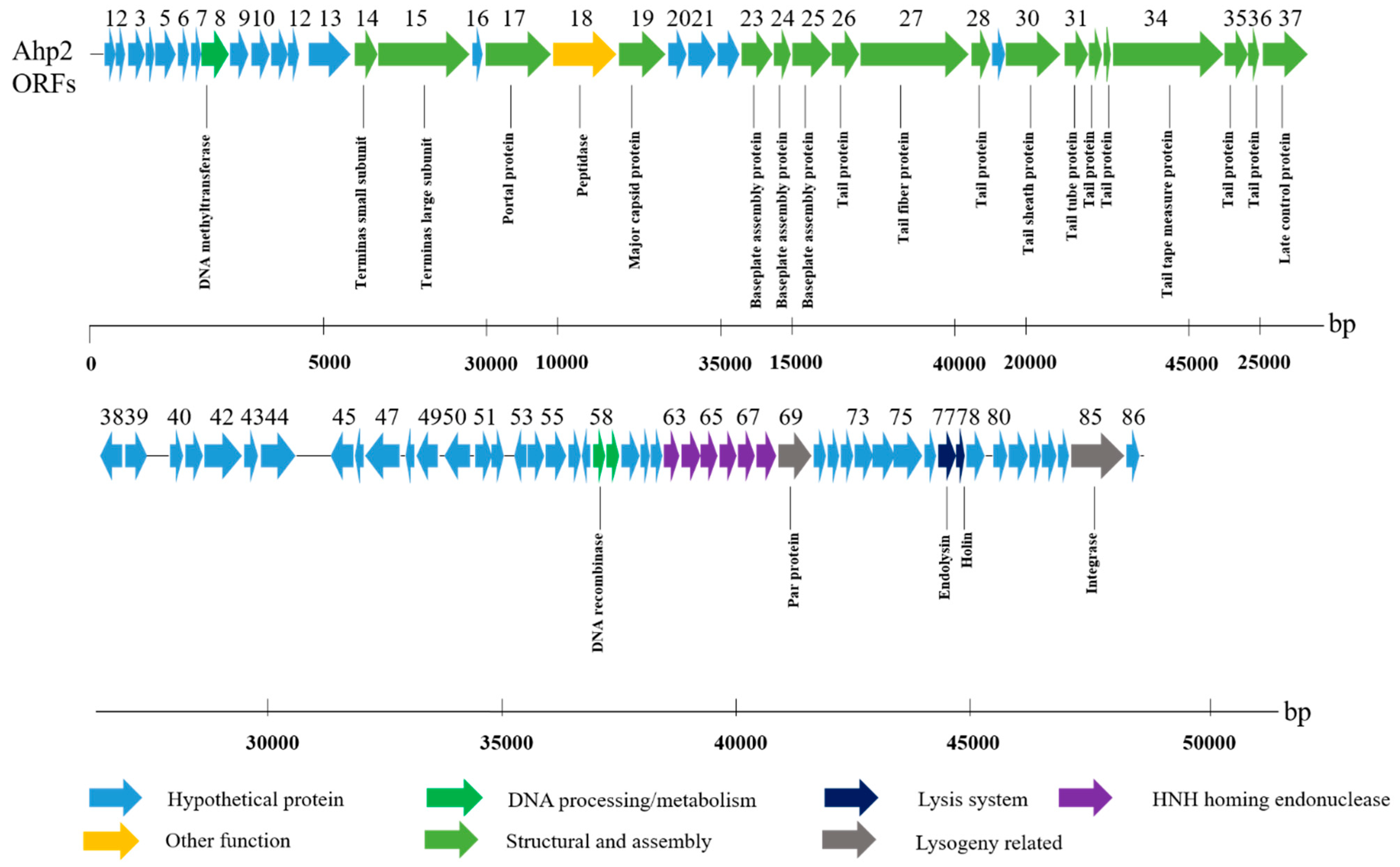
3.3. General Properties of the Ahp2 Genome
3.4. Genome Comparison of Ahp2 and Similar Aeromonas Phages
3.5. Gene Products and Their Functions
3.5.1. Morphogenesis and Structural Genes
3.5.2. Genes Involved in Replication and Recombination
3.5.3. Lysis and Lysogeny Genes
3.5.4. Other Open Reading Frames (ORFs)
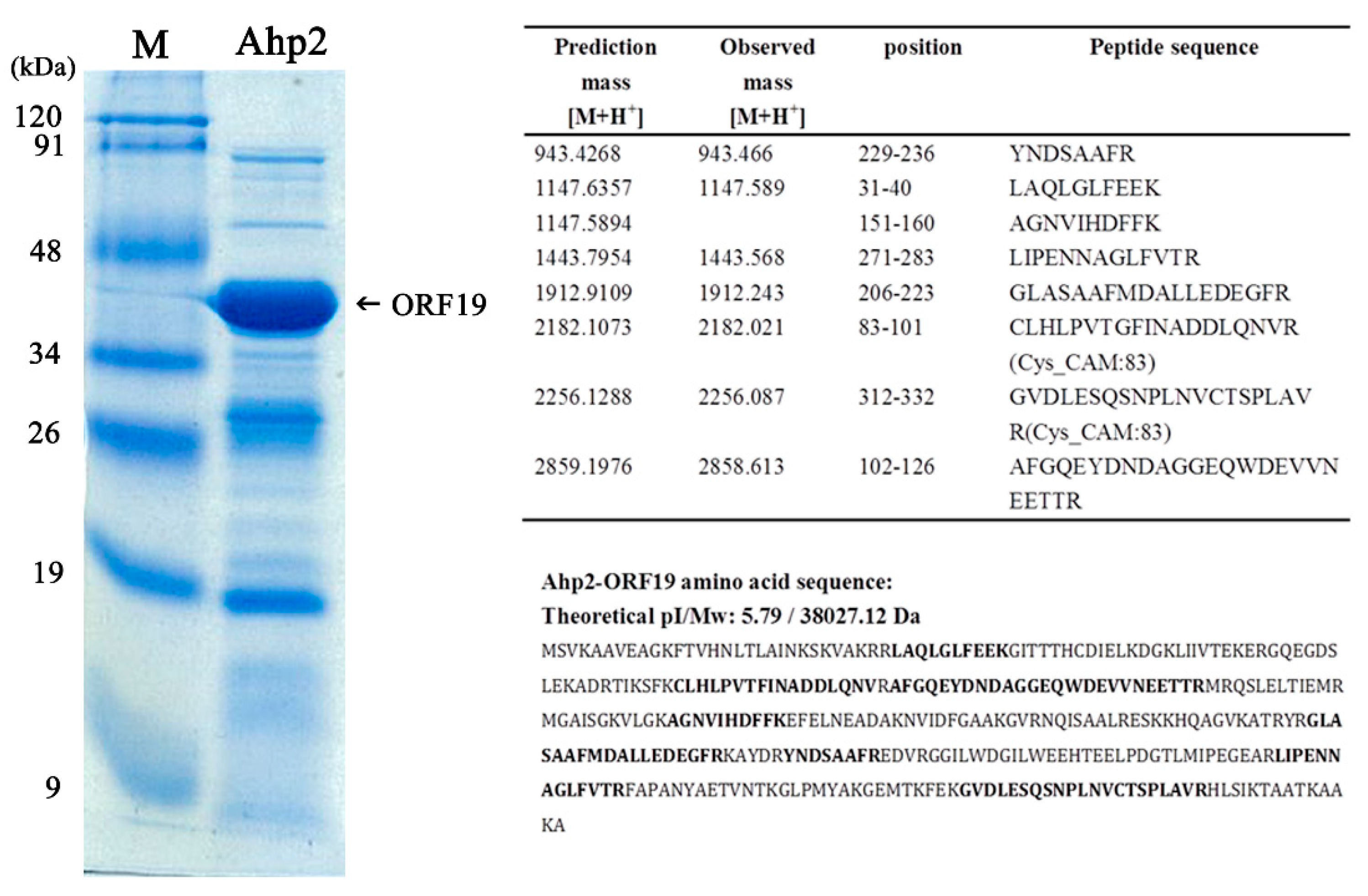
3.6. Characterization of the Most Abundant Protein in the Ahp2 Virion
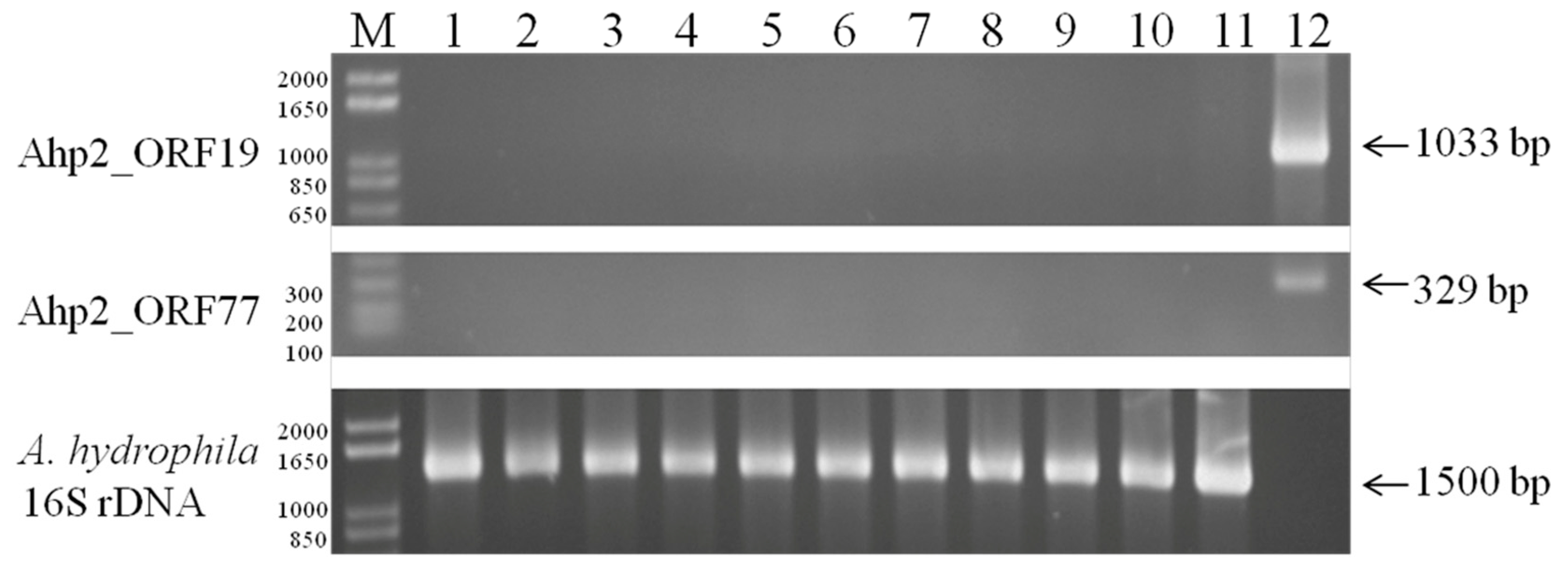
3.7. Detection of Lysogen in Ahp2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Janda, J.M.; Abbott, S.L. The Genus Aeromonas: Taxonomy, Pathogenicity, and Infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.M.; Lai, C.C.; Tang, H.J.; Ko, W.C.; Hsueh, P.R. Skin and Soft-Tissue Infections Caused by Aeromonas species. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.M.; Lai, C.C.; Tsai, H.Y.; Wu, C.J.; Tang, H.J.; Ko, W.C.; Hsueh, P.R. Pneumonia Caused by Aeromonas species in Taiwan, 2004–2011. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Picão, R.C.; Cardoso, J.P.; Campana, E.H.; Nicoletti, A.G.; Petrolini, F.V.B.; Assis, D.M.; Juliano, L.; Gales, A.C. The Route of Antimicrobial Resistance from the Hospital Effluent to the Environment: Focus on the Occurrence of KPC-Producing Aeromonas spp. and Enterobacteriaceae in Sewage. Diagn. Microbiol. Infect. Dis. 2013, 76, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.N.K.; Van, T.T.H.; Nguyen, H.T.; Smooker, P.M.; Shimeta, J.; Coloe, P.J. Molecular Characterization of Antibiotic Resistance in Pseudomonas and Aeromonas Isolates from Catfish of the Mekong Delta, Vietnam. Vet. Microbiol. 2014, 171, 397–405. [Google Scholar] [CrossRef]
- Dobiasova, H.; Kutilova, I.; Piackova, V.; Vesely, T.; Cizek, A.; Dolejska, M. Ornamental Fish as a Source of Plasmid-Mediated Quinolone Resistance Genes and Antibiotic Resistance Plasmids. Vet. Microbiol. 2014, 171, 413–421. [Google Scholar] [CrossRef]
- Fosse, T.; Giraud-Morin, C.; Madinier, I.; Mantoux, F.; Lacour, J.P.; Ortonne, J.P. Aeromonas hydrophila with Plasmid-Borne class A Extended-Spectrum β-Lactamase TEM-24 and Three Chromosomal class B, C, and D β-Lactamases, Isolated from a Patient with Necrotizing Fasciitis. Antimicrob. Agents Chemother. 2004, 48, 2342–2343. [Google Scholar] [CrossRef]
- Chen, P.L.; Ko, W.C.; Wu, C.J. Complexity of β-Lactamases Among Clinical Aeromonas Isolates and Its Clinical Implications. J. Microbiol. Immunol. Infect. 2012, 45, 398–403. [Google Scholar] [CrossRef]
- Esteve, C.; Alcaide, E.; Giménez, M.J. Multidrug-Resistant (MDR) Aeromonas Recovered from the Metropolitan Area of Valencia (Spain): Diseases Spectrum and Prevalence in the Environment. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Hughes, H.Y.; Conlan, S.P.; Lau, A.F.; Dekker, J.P.; Michelin, A.V.; Youn, J.H.; Henderson, D.K.; Frank, K.M.; Segre, J.A.; Palmore, T.N. Detection and Whole-Genome Sequencing of Carbapenemase-Producing Aeromonas hydrophila Isolates from Routine Perirectal Surveillance Culture. J. Clin. Microbiol. 2016, 54, 1167–1170. [Google Scholar] [CrossRef] [PubMed]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G. Bacteriophage Therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C. Where Will New Antibiotics Come From? Nat. Rev. Microbiol. 2003, 1, 65–70. [Google Scholar] [CrossRef]
- Hesse, S.; Adhya, S. Phage Therapy in the Twenty-First Century: Facing the Decline of the Antibiotic Era; Is It Finally Time for the Age of the Phage? Annu. Rev. Microbiol. 2019, 73, 155–174. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Yasuda, M.; Nishikawa, H.; Kuroda, M.; Ujihara, T.; Shuin, T.; Shen, Y.; Jin, Z.; Fujimoto, S.; Nasimuzzaman, M.D.; et al. Experimental Protection of Mice Against Lethal Staphylococcus aureus Infection by Novel Bacteriophage Phi MR11. J. Infect. Dis. 2003, 187, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.W.; Kim, J.H.; Shin, S.P.; Han, J.E.; Chai, J.Y.; Park, S.C. Protective Effects of the Aeromonas Phages pAh1-C and pAh6-C Against Mass Mortality of the Cyprinid Loach (Misgurnus anguillicaudatus) Caused by Aeromonas hydrophila. Aquaculture 2013, 416–417, 289–295. [Google Scholar] [CrossRef]
- Soothill, J.S. Bacteriophage Prevents Destruction of Skin Grafts by Pseudomonas aeruginosa. Burns 1994, 20, 209–211. [Google Scholar] [CrossRef]
- Weber-Dabrowska, B.; Mulczyk, M.; Górski, A. Bacteriophages as an Efficient Therapy for Antibiotic-Resistant Septicemia in Man. Transpl. Proc. 2003, 35, 1385–1386. [Google Scholar] [CrossRef]
- Jikia, D.; Chkhaidze, N.; Imedashvili, E.; Mgaloblishvili, I.; Tsitlanadze, G.; Katsarava, R.; Glenn Morris, J.; Sulakvelidze, A. The Use of a Novel Biodegradable Preparation Capable of the Sustained Release of Bacteriophages and Ciprofloxacin, in the Complex Treatment of Multidrug-Resistant Staphylococcus aureus-Infected Local Radiation Injuries Caused by Exposure to Sr90. Clin. Exp. Derm. 2005, 30, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.B.; Lin, N.T.; Tseng, Y.H.; Weng, S.F. Genomic Characterization of the Novel Aeromonas hydrophila Phage Ahp1 Suggests the Derivation of a New Subgroup from phiKMV-Like Family. PLoS ONE 2016, 11, e0162060. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, S.; Wang, D.; Zhao, J.; Xu, L.; Liu, H.; Lu, T.; Mou, Z. Genomic Characterization of a Novel Virulent Phage Infecting the Aeromonas hydrophila Isolated from Rainbow Trout (Oncorhynchus mykiss). Virus Res. 2019, 273, 197764. [Google Scholar] [CrossRef]
- Kazimierczak, J.; Wójcik, E.A.; Witaszewska, J.; Guziński, A.; Górecka, E.; Stańczyk, M.; Kaczorek, E.; Siwicki, A.K.; Dastych, J. Complete Genome Sequences of Aeromonas and Pseudomonas Phages as a Supportive Tool for Development of Antibacterial Treatment in Aquaculture. Virol. J. 2019, 16, 4. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Chen, L.; Liu, Q.; Zhou, Y.; Yang, J.; Deng, D.; Li, H.; Ma, Y. Characterization and Genomic Analyses of Aeromonas hydrophila Phages AhSzq-1 and AhSzw-1, Isolates Representing New Species Within the T5virus Genus. Arch. Virol. 2018, 163, 1985–1988. [Google Scholar] [CrossRef] [PubMed]
- Akmal, M.; Rahimi-Midani, A.; Hafeez-Ur-Rehman, M.; Hussain, A.; Choi, T.J. Isolation, Characterization, and Application of a Bacteriophage Infecting the Fish Pathogen Aeromonas hydrophila. Pathogens 2020, 9, 215. [Google Scholar] [CrossRef]
- Merino, S.; Camprubi, S.; Tomás, J.M. Isolation and Characterization of Bacteriophage PM3 from Aeromonas hydrophila the Bacterial Receptor for Which Is the Monopolar Flagellum. Fems Microbiol. Lett. 1990, 57, 277–282. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wu, J.L.; Lin, H.M.; Jan, L.; Hsu, Y.L.; Chang, L.H. Biological Control of Fish Bacterial Pathogen, Aeromonas hydrophila, by Bacteriophage AH1. Fish Pathol. 1981, 1, 271–276. [Google Scholar] [CrossRef]
- Chow, M.S.; Rouf, M.A. Isolation and Partial Characterization of Two Aeromonas hydrophila Bacteriophages. Appl. Environ. Microbiol. 1983, 45, 1670–1676. [Google Scholar] [CrossRef]
- Merino, S.; Camprubi, S.; Tomás, J.M. Isolation and Characterization of Bacteriophage PM2 from Aeromonas hydrophila. Fems Microbiol. Lett. 1990, 56, 239–244. [Google Scholar] [CrossRef]
- Jun, J.W.; Kim, H.J.; Yun, S.K.; Chai, J.Y.; Park, S.C. Genomic Structure of the Aeromonas Bacteriophage pAh6-C and Its Comparative Genomic Analysis. Arch. Virol. 2015, 160, 561–564. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Gao, D.G.; Xia, Y.S.; Wang, Z.Y.; Bai, M.; Luo, K.J.; Cui, X.L.; Wang, Y.G.; Zhang, S.Y.; Xiao, W. Characterization of Novel Bacteriophage AhyVDH1 and Its Lytic Activity Against Aeromonas hydrophila. Curr. Microbiol. 2021, 78, 329–337. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S Ribosomal DNA Amplification for Phylogenetic Study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on Transformation of Escherichia coli with Plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Yang, B.Y.; Tsai, H.F.; Tseng, Y.H. Broad Host Range Cosmid pLAFR1 and Non-Mucoid Mutant XCP20 Provide a Suitable Vector-Host System for Cloning Genes in Xanthomonas campestris pv. campestris. Chin. J. Microbiol. Immunol. 1988, 2, 40–49. [Google Scholar]
- Lin, N.T.; Chiou, P.Y.; Chang, K.C.; Chen, L.K.; Lai, M.J. Isolation and Characterization of Phi AB2: A Novel Bacteriophage of Acinetobacter baumannii. Res. Microbiol. 2010, 161, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Kawato, Y.; Yasuike, M.; Nakamura, Y.; Shigenobu, Y.; Fujiwara, A.; Sano, M.; Nakai, T. Complete Genome Sequence Analysis of Two Pseudomonas plecoglossicida Phages, Potential Therapeutic Agents. Appl. Environ. Microbiol. 2015, 81, 874–881. [Google Scholar] [CrossRef]
- Chang, H.C.; Chen, C.R.; Lin, J.W.; Shen, G.H.; Chang, K.M.; Tseng, Y.H.; Weng, S.F. Isolation and Characterization of Novel Giant Stenotrophomonas maltophilia Phage phiSMA5. Appl. Environ. Microbiol. 2005, 71, 1387–1393. [Google Scholar] [CrossRef]
- Tseng, Y.H.; Choy, K.T.; Hung, C.H.; Lin, N.T.; Liu, J.Y.; Lou, C.H.; Yang, B.Y.; Wen, F.S.; Weng, S.F.; Wu, J.R. Chromosome Map of Xanthomonas campestris pv. campestris 17 with Locations of Genes Involved in Xanthan Gum Synthesis and Yellow Pigmentation. J. Bacteriol. 1999, 181, 117–125. [Google Scholar] [CrossRef]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A Self-Training Method for Prediction of Gene Starts in Microbial Genomes. Implications for Finding Sequence Motifs in Regulatory Regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [PubMed]
- Alva, V.; Nam, S.Z.; Söding, J.; Lupas, A.N. The MPI Bioinformatics Toolkit as an Integrative Platform for Advanced Protein Sequence and Structure Analysis. Nucleic Acids Res. 2016, 44, W410–W415. [Google Scholar] [CrossRef]
- Hildebrand, A.; Remmert, M.; Biegert, A.; Söding, J. Fast and Accurate Automatic Structure Prediction with HHpred. Proteins 2009, 77, 128–132. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete genomes11Edited. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Vincent, A.T.; Paquet, V.E.; Bernatchez, A.; Tremblay, D.M.; Moineau, S.; Charette, S.J. Characterization and Diversity of Phages Infecting Aeromonas salmonicida subsp. salmonicida. Sci. Rep. 2017, 7, 7054. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.B.; Feiss, M. The Bacteriophage DNA Packaging Motor. Annu. Rev. Genet. 2008, 42, 647–681. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, C.; King, J. The DNA Translocating Vertex of dsDNA Bacteriophage. Annu. Rev. Microbiol. 1985, 39, 109–129. [Google Scholar] [CrossRef]
- Young, R. Bacteriophage Lysis: Mechanism and Regulation. Microbiol. Rev. 1992, 56, 430–481. [Google Scholar] [CrossRef]
- Borysowski, J.; Weber-Dąbrowska, B.; Górski, A. Bacteriophage Endolysins as a Novel Class of Antibacterial Agents. Exp. Biol. Med. 2006, 231, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.N.; Smith, D.L.; Young, R. Holins: The Protein Clocks of Bacteriophage Infections. Annu. Rev. Microbiol. 2000, 54, 799–825. [Google Scholar] [CrossRef]
- Sergueev, K.; Dabrazhynetskaya, A.; Austin, S. Plasmid Partition System of the P1par Family from the pWR100 Virulence Plasmid of Shigella flexneri. J. Bacteriol. 2005, 187, 3369–3373. [Google Scholar] [CrossRef]
- Walker, J.E.; Saraste, M.; Runswick, M.J.; Gay, N.J. Distantly Related Sequences in the Alpha- and Beta-Subunits of ATP Synthase, Myosin, Kinases and Other ATP-Requiring Enzymes and a Common Nucleotide Binding Fold. Embo J. 1982, 1, 945–951. [Google Scholar] [CrossRef]
- Nobrega, F.L.; Vlot, M.; de Jonge, P.A.; Dreesens, L.L.; Beaumont, H.J.E.; Lavigne, R.; Dutilh, B.E.; Brouns, S.J.J. Targeting mechanisms of tailed bacteriophages. Nat. Rev. Microbiol. 2018, 16, 760–773. [Google Scholar] [CrossRef]
- Grose, J.H.; Casjens, S.R. Understanding the enormous diversity of bacteriophages: The tailed phages that infect the bacterial family Enterobacteriaceae. Virology 2014, 468–470, 421–443. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, C.; Caumont-Sarcos, A.; Comeau, A.M.; Krisch, H.M. Isolation and genomic characterization of the first phage infecting Iodobacteria: ϕPLPE, a myovirus having a novel set of features. Environ. Microbiol. Rep. 2009, 1, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Adesanya, O.; Oduselu, T.; Akin-Ajani, O.; Adewumi, O.M.; Ademowo, O.G. An exegesis of bacteriophage therapy: An emerging player in the fight against anti-microbial resistance. AIMS Microbiol. 2020, 6, 204–230. [Google Scholar] [CrossRef]
- Cisek, A.A.; Dabrowska, I.; Gregorczyk, K.P.; Wyzewski, Z. Phage Therapy in Bacterial Infections Treatment: One Hundred Years After the Discovery of Bacteriophages. Curr. Microbiol. 2017, 74, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Jault, P.; Leclerc, T.; Jennes, S.; Pirnay, J.P.; Que, Y.A.; Resch, G.; Rousseau, A.F.; Ravat, F.; Carsin, H.; Le Floch, R.; et al. Efficacy and tolerability of a cocktail of bacteriophages to treat burn wounds infected by Pseudomonas aeruginosa (PhagoBurn): A randomised, controlled, double-blind phase 1/2 trial. Lancet Infect. Dis. 2019, 19, 35–45. [Google Scholar] [CrossRef]
- Wright, A.; Hawkins, C.H.; Änggård, E.E.; Harper, D.R. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; A preliminary report of efficacy. Clin. Otolaryngol. 2009, 34, 349–357. [Google Scholar] [CrossRef]
- Carlton, R.M.; Noordman, W.H.; Biswas, B.; De Meester, E.D.; Loessner, M.J. Bacteriophage P100 forcontrol of Listeria monocytogenes in foods: Genome sequence, bioinformatic analyses, oral toxicity study, and application. Regul. Toxicol. Pharmacol. 2005, 43, 301–312. [Google Scholar] [CrossRef]
- Kowalska, J.D.; Kazimierczak, J.; Sowinska, P.M.; Wojcik, E.A.; Siwicki, A.K.; Dastych, J. Growing Trend of Fighting Infections in Aquaculture Environment-Opportunities and Challenges of Phage Therapy. Antibiotics 2020, 9, 301. [Google Scholar] [CrossRef]
- Hoang, A.H.; Tran, T.T.X.; Le, P.N.; Dang, T.H.O. Selection of Phages to Control Aeromonas hydrophila—An Infectious Agent in Striped Catfish. Biocontrol Sci. 2019, 24, 23–28. [Google Scholar] [CrossRef]
- Chen, L.; Yuan, S.; Liu, Q.; Mai, G.; Yang, J.; Deng, D.; Zhang, B.; Liu, C.; Ma, Y. In Vitro Design and Evaluation of Phage Cocktails Against Aeromonas salmonicida. Front Microbiol. 2018, 6, 1476. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Descriptions | Reference or Source |
|---|---|---|
| A. hydrophila | ||
| ATCC7966 | Type strain, Ap r | ATCC |
| ATCC43414 | Type strain, Ap r | ATCC |
| AH19288 | Clinical isolate from Buddhist Tzu Chi General Hospital, Ap r | [19] |
| AH60114, AH300206 | Clinical isolates from Hualien Armed Forces General Hospital, Ap r | [19] |
| Hua-1, Hua-2 | Fish isolates from Hualien Animal and Plant Disease Control Center, Ap r | [19] |
| H1 to H35 | Environmental isolates, Ap r | [19] |
| Acinetobacter baumannii | ||
| ATCC17978 | Type strain, Ap r | ATCC |
| Escherichia coli | ||
| DH5α | F− φ80dlacZΔM15Δ(lacZYA-argF) U169 recA1 endA1 hsdR17 (rk−, mk+) phoA supE44 λ− thi-1 gyrA96 relA1 | [31] |
| Klebsiella pneumonia | ||
| Kp-6 | Clinical isolate, Ap r | N. T. Lin a |
| Staphylococcus aureus | ||
| NCTC8325 | Type strain, Ap r | NCTC (National Collection of Type Cultures) |
| Vibrio parahaemolyticus | ||
| VP93 | Clinical isolate, Ap r | M. S. Yu a |
| Vibrio harveyi | ||
| BAA-1117 | luxN::tn5Kan | ATCC |
| Xanthomonas campestris pv. campestris | ||
| P20H | Nonmucoid mutant, Ap r | [32] |
| Genes | Primer Name | Primer Sequence | Product Size (bp) | Reference |
|---|---|---|---|---|
| 16S rDNA | 27F | 5′-AGAGTTTGATCMTGGCTCAG | 1500 | [30] |
| 1492R | 5′-TACGGYTACCTTGTTACGACTT | |||
| Ahp2_ORF19 (coat protein) | MCP_F | 5′-GCGTAAAAGCTGCCGTAGAA | 1033 | This study |
| MCP_R | 5′-GCTTTAGCTGCCTTGGTTGC | |||
| Ahp2_ORF77 (endopeptidase) | 77_F | 5′-AAGACGTAAAGCTGCGCTGC | 329 | This study |
| 77_R | 5′-TTAGTAGTCAAAAACGATGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.-B.; Yu, M.-S.; Tseng, T.-T.; Lin, L.-C. Molecular Characterization of Ahp2, a Lytic Bacteriophage of Aeromonas hydrophila. Viruses 2021, 13, 477. https://doi.org/10.3390/v13030477
Wang J-B, Yu M-S, Tseng T-T, Lin L-C. Molecular Characterization of Ahp2, a Lytic Bacteriophage of Aeromonas hydrophila. Viruses. 2021; 13(3):477. https://doi.org/10.3390/v13030477
Chicago/Turabian StyleWang, Jian-Bin, Mei-Shiuan Yu, Tsai-Tien Tseng, and Ling-Chun Lin. 2021. "Molecular Characterization of Ahp2, a Lytic Bacteriophage of Aeromonas hydrophila" Viruses 13, no. 3: 477. https://doi.org/10.3390/v13030477
APA StyleWang, J.-B., Yu, M.-S., Tseng, T.-T., & Lin, L.-C. (2021). Molecular Characterization of Ahp2, a Lytic Bacteriophage of Aeromonas hydrophila. Viruses, 13(3), 477. https://doi.org/10.3390/v13030477