
Experimental Inoculation of Young Calves with SARS-CoV-2


 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Methods
2.1. Experimental Design and Sample Collection
2.2. Assessing the Presence or Exposure to Other Bovine Respiratory Pathogens
2.3. Cells and Virus
2.4. Cell Susceptibility
2.5. RNA Extraction and Real-Time RT-PCR (RT-rPCR)
2.6. Virus Isolation
2.7. Serological Analysis
2.8. In Situ Hybridization (ISH)
3. Results
3.1. Assessing In Vitro Replication of SARS-CoV-2 in Bovine Cells
3.2. Clinical Parameters and Assessment of Other Bovine Pathogens
3.3. Neutralizing Antibody Response to SARS-CoV-2 Infection
3.4. SARS-CoV-2 Nucleic Acid Detection
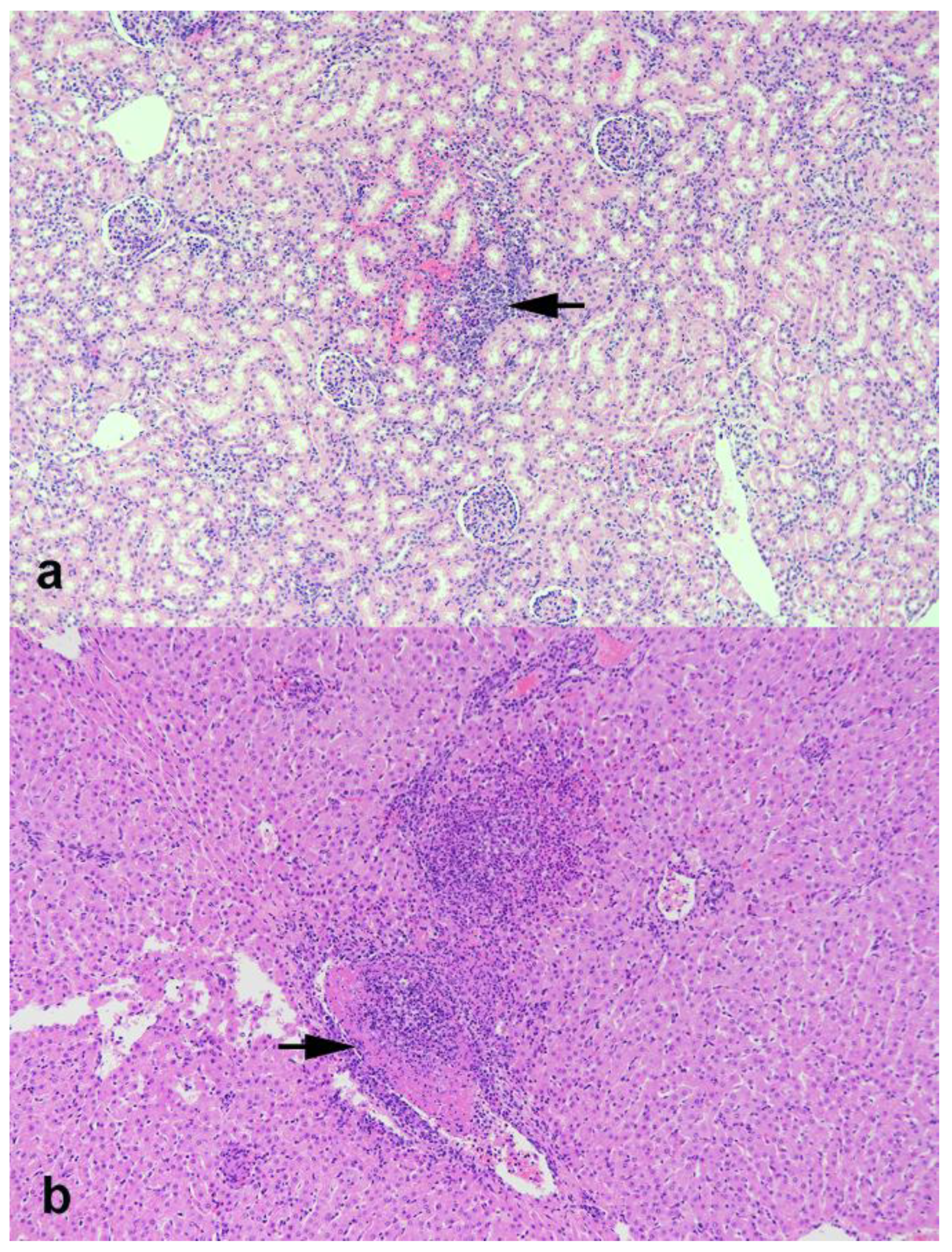
3.5. Gross and Histologic Changes at Necropsy
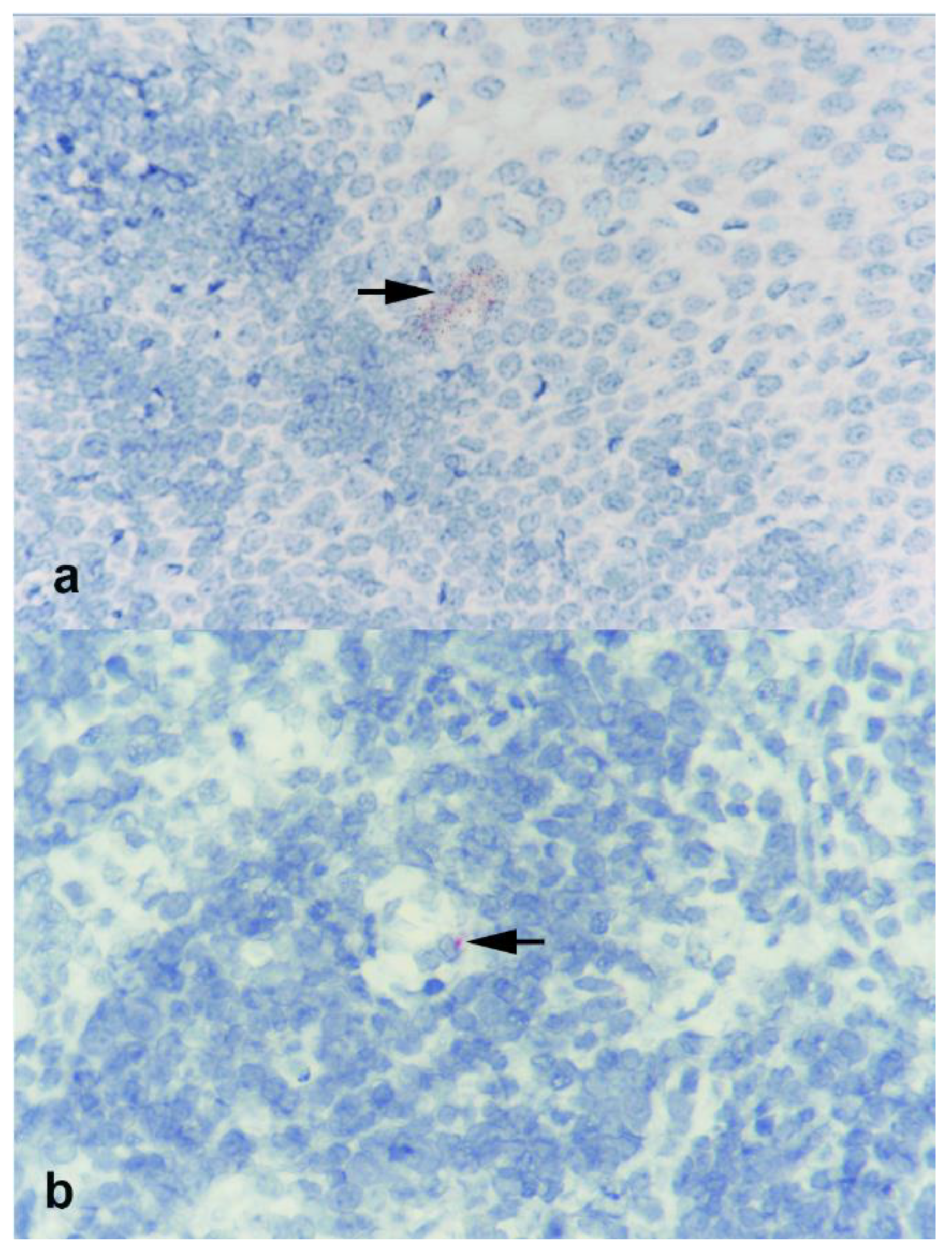
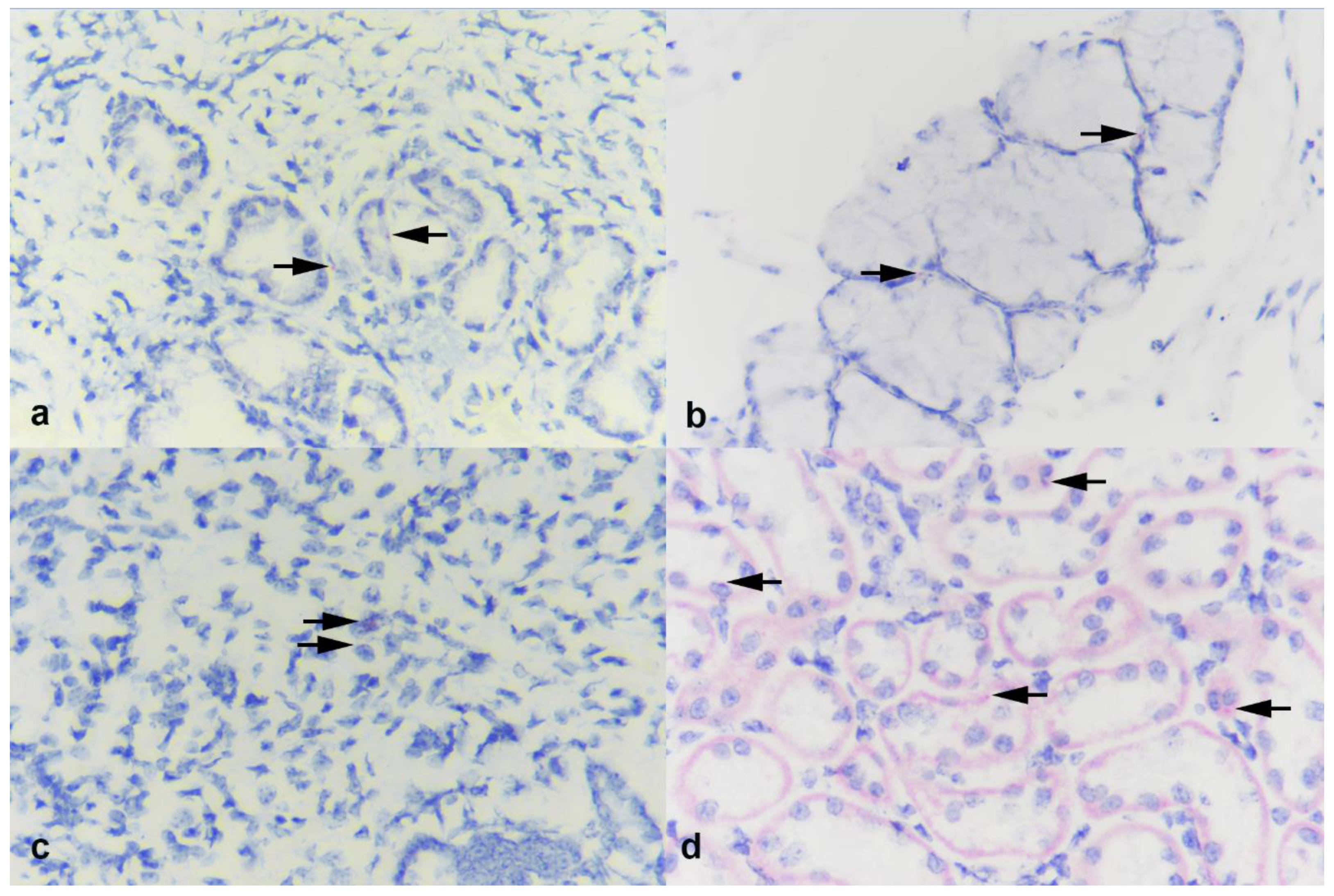
3.6. In Situ Hybridization (ISH)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef]
- McAloose, D.; Laverack, M.; Wang, L.; Killian, M.L.; Caserta, L.C.; Yuan, F.; Mitchell, P.K.; Queen, K.; Mauldin, M.R.; Cronk, B.D. From people to Panthera: Natural SARS-CoV-2 infection in tigers and lions at the Bronx Zoo. Mbio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Paraskevis, D.; Kostaki, E.G.; Magiorkinis, G.; Panayiotakopoulos, G.; Sourvinos, G.; Tsiodras, S. Full-genome evolutionary analysis of the novel corona virus (2019-nCoV) rejects the hypothesis of emergence as a result of a recent recombination event. Infect. Genet. Evol. 2020, 79, 104212. [Google Scholar] [CrossRef]
- Decaro, N.; Lorusso, A. Novel human coronavirus (SARS-CoV-2): A lesson from animal coronaviruses. Vet. Microbiol. 2020, 244, 108693. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor recognition by the novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS coronavirus. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef]
- Luan, J.; Lu, Y.; Jin, X.; Zhang, L. Spike protein recognition of mammalian ACE2 predicts the host range and an optimized ACE2 for SARS-CoV-2 infection. Biochem. Biophys. Res. Commun. 2020, 526, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; Hughes, G.M.; Keough, K.C.; Painter, C.A.; Persky, N.S.; Corbo, M.; Hiller, M.; Koepfli, K.-P.; Pfenning, A.R.; Zhao, H. Broad Host Range of SARS-CoV-2 Predicted by Comparative and Structural Analysis of ACE2 in Vertebrates. bioRxiv 2020. [Google Scholar] [CrossRef]
- Palmer, M.V.; Martins, M.; Falkenberg, S.; Buckley, A.C.; Caserta, L.C.; Mitchell, P.K.; Cassmann, E.; Rollins, A.; Zylich, N.C.; Renshaw, R.W. Susceptibility of white-tailed deer (Odocoileus virginianus) to SARS-CoV-2. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, H.; Huang, K.; Yang, Y.; Hui, X.; Gao, J.; He, X.; Li, C.; Gong, W.; Zhang, Y. SARS-CoV-2 neutralizing serum antibodies in cats: A serological investigation. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sit, T.H.; Brackman, C.J.; Ip, S.M.; Tam, K.W.; Law, P.Y.; To, E.M.; Yu, V.Y.; Sims, L.D.; Tsang, D.N.; Chu, D.K. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef]
- Oreshkova, N.; Molenaar, R.-J.; Vreman, S.; Harders, F.; Munnink, B.B.O.; Honing, R.W.H.-V.; Gerhards, N.; Tolsma, P.; Bouwstra, R.; Sikkema, R. SARS-CoV2 infection in farmed mink, Netherlands, April 2020. bioRxiv 2020. [Google Scholar] [CrossRef]
- Chandrashekar, A.; Liu, J.; Martinot, A.J.; McMahan, K.; Mercado, N.B.; Peter, L.; Tostanoski, L.H.; Yu, J.; Maliga, Z.; Nekorchuk, M. SARS-CoV-2 infection protects against rechallenge in rhesus macaques. Science 2020, 369, 812–817. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS–coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef]
- Munster, V.J.; Feldmann, F.; Williamson, B.N.; van Doremalen, N.; Pérez-Pérez, L.; Schulz, J.; Meade-White, K.; Okumura, A.; Callison, J.; Brumbaugh, B. Respiratory disease in rhesus macaques inoculated with SARS-CoV-2. Nature 2020, 585, 268–272. [Google Scholar] [CrossRef]
- Hartman, A.L.; Nambulli, S.; McMillen, C.M.; White, A.G.; Tilston-Lunel, N.L.; Albe, J.R.; Cottle, E.; Dunn, M.D.; Frye, L.J.; Gilliland, T.H. SARS-CoV-2 infection of African green monkeys results in mild respiratory disease discernible by PET/CT imaging and shedding of infectious virus from both respiratory and gastrointestinal tracts. PLoS Pathog. 2020, 16, e1008903. [Google Scholar] [CrossRef] [PubMed]
- Shuai, L.; Zhong, G.; Yuan, Q.; Wen, Z.; Wang, C.; He, X.; Liu, R.; Wang, J.; Zhao, Q.; Liu, Y. Replication, pathogenicity, and transmission of SARS-CoV-2 in minks. Natl. Sci. Rev. 2020. [Google Scholar] [CrossRef]
- Freuling, C.; Breithaupt, A.; Müller, T.; Sehl, J.; Balkema-Buschmann, A.; Rissmann, M.; Klein, A.; Wylezich, C.; Höper, D.; Wernike, K. Susceptibility of Raccoon Dogs for Experimental SARS-CoV-2 Infection. Emerg. Infect. Dis. 2020, 26, 2982–2985. [Google Scholar] [CrossRef]
- Griffin, B.D.; Chan, M.; Tailor, N.; Mendoza, E.J.; Leung, A.; Warner, B.M.; Duggan, A.T.; Moffat, E.; He, S.; Garnett, L. North American deer mice are susceptible to SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ulrich, L.; Wernike, K.; Hoffmann, D.; Mettenleiter, T.C.; Beer, M. Experimental infection of cattle with SARS-CoV-2. Emerg. Infect. Dis. 2020, 26, 2979. [Google Scholar] [CrossRef]
- Pickering, B.; Smith, G.; Pinette, M.; Embury-Hyatt, C.; Moffat, E.; Marszal, P.; Lewis, C.E. Susceptibility of domestic swine to experimental infection with SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Suarez, D.L.; Pantin-Jackwood, M.J.; Swayne, D.E.; Lee, S.A.; Deblois, S.M.; Spackman, E. Lack of susceptibility of poultry to SARS-CoV-2 and MERS-CoV. Emerg. Infect. Dis. 2021, 26, 3074–3076. [Google Scholar] [CrossRef]
- Schlottau, K.; Rissmann, M.; Graaf, A.; Schön, J.; Sehl, J.; Wylezich, C.; Höper, D.; Mettenleiter, T.C.; Balkema-Buschmann, A.; Harder, T. SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: An experimental transmission study. Lancet Microbe 2020, 1, e218–e225. [Google Scholar] [CrossRef]
- Meekins, D.A.; Morozov, I.; Trujillo, J.D.; Gaudreault, N.N.; Bold, D.; Carossino, M.; Artiaga, B.L.; Indran, S.V.; Kwon, T.; Balaraman, V. Susceptibility of swine cells and domestic pigs to SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 2278–2288. [Google Scholar] [CrossRef]
- Di Teodoro, G.; Valleriani, F.; Puglia, I.; Monaco, F.; di Pancrazio, C.; Luciani, M.; Krasteva, I.; Petrini, A.; Marcacci, M.; D’Alterio, N. SARS-CoV-2 replicates in respiratory ex vivo organ cultures of domestic ruminant species. Vet. Microbiol. 2021, 252, 108933. [Google Scholar] [CrossRef]
- Sun, K.; Gu, L.; Ma, L.; Duan, Y. Atlas of ACE2 gene expression in mammals reveals novel insights in transmission of SARS-Cov-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ridpath, J.F.; Falkenberg, S.M.; Bauermann, F.V.; VanderLey, B.L.; Do, Y.; Flores, E.F.; Rodman, D.M.; Neill, J.D. Comparison of acute infection of calves exposed to a high-virulence or low-virulence bovine viral diarrhea virus or a HoBi-like virus. Am. J. Vet. Res. 2013, 74, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Falkenberg, S.; Ridpath, J.; Ley, B.V.; Bauermann, F.; Sanchez, N.B.; Carroll, J. Comparison of temperature fluctuations at multiple anatomical locations in cattle during exposure to bovine viral diarrhea virus. Livest. Sci. 2014, 164, 159–167. [Google Scholar] [CrossRef]
- Ridpath, J.F.; Fulton, R.W.; Bauermann, F.V.; Falkenberg, S.M.; Welch, J.; Confer, A.W. Sequential exposure to bovine viral diarrhea virus and bovine coronavirus results in increased respiratory disease lesions: Clinical, immunologic, pathologic, and immunohistochemical findings. J. Vet. Diagn. Investig. 2020, 32, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Nao, N.; Shirato, K.; Kawase, M.; Saito, S.; Takayama, I.; Nagata, N.; Sekizuka, T.; Katoh, H.; Kato, F.; et al. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. USA 2020, 117, 7001–7003. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.-W.; Xu, D.; Zhang, H.; Zhou, W.; Wang, L.-H.; Cui, X.-G. Identification of a potential mechanism of acute kidney injury during the COVID-19 outbreak: A study based on single-cell transcriptome analysis. Intensive Care Med. 2020, 46, 1114–1116. [Google Scholar] [CrossRef]
- Batlle, D.; Soler, M.J.; Sparks, M.A.; Hiremath, S.; South, A.M.; Welling, P.A.; Swaminathan, S. Acute kidney injury in COVID-19: Emerging evidence of a distinct pathophysiology. J. Am. Soc. Nephrol. 2020, 31, 1380–1383. [Google Scholar] [CrossRef]
- Doi, K.; Leelahavanichkul, A.; Yuen, P.S.; Star, R.A. Animal models of sepsis and sepsis-induced kidney injury. J. Clin. Investig. 2009, 119, 2868–2878. [Google Scholar] [CrossRef] [PubMed]
- Buntyn, J.; Carroll, J.; Smith, T.; Rivera, J.; Sanchez, N.B.; Broadway, P.; Falkenberg, S.; Schmidt, T. Evaluation of the innate immune response of Angus heifers with genetic marker variation for intramuscular fat deposition following a lipopolysaccharide challenge. Livest. Sci. 2016, 188, 103–110. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Route | Intratracheal Inoculation Group | Intravenous Inoculation Group | ||||
|---|---|---|---|---|---|---|
| Calf ID | 7441 | 7432 | 7435 | 7436 | 7439 | 7434 |
| Sample a day | ||||||
| −3 | - | - | - | - | - | - |
| −2 | - | - | - | - | - | - |
| −1 | - | - | - | - | - | - |
| 0 | - | - | - | - | - | - |
| 1 | - | - | - | - | - | - |
| 2 | - | - | - | - | - | - |
| 3 | + * | - | - | - | - | - |
| 4 | - | - | - | - | - | - |
| 5 | - | - | - | - | - | - |
| 6 | - | - | - | - | - | - |
| 7 | - | - | - | - | - | - |
| 8 | - | - | - | - | - | - |
| 9 | - | - | - | - | - | - |
| 10 | - | - | - | + * | ||
| 11 | - | - | - | - | ||
| 12 | - | - | - | - | ||
| 13 | - | - | - | - | ||
| 14 | - | - | - | - | ||
| 15 | - | - | - | - | ||
| 16 | - | - | - | - | ||
| 17 | - | - | ||||
| 18 | - | - | ||||
| 19 | - | - | ||||
| 20 | - | - | ||||
| 21 | - | - | ||||
| Intratracheal Inoculation Group | Intravenous Inoculation Group | |||||
|---|---|---|---|---|---|---|
| Calf ID | 7441 | 7432 | 7435 | 7436 | 7439 | 7434 |
| Necropsy day | 9 | 16 | 21 | 9 | 16 | 21 |
| adrenals | - | - | - | - | ||
| bifurcation trachea | - | - | - | - | - | - |
| bronchi | - | - | - | - | - | - |
| bronchioles | - | - | - | - | - | - |
| cerebrum | - | - | - | - | - | - |
| colon | - | - | - | - | - | - |
| heart | - | - | - | - | - | - |
| hepatic LN | - | |||||
| ICE junction | - | - | - | - | ||
| kidney | - | - | - | - | - | - |
| left cerebellum | - | - | - | - | - | - |
| liver | - | - | - | - | - | |
| lung | - | - | - | - | - | - |
| lung lesion | - | - | - | - | - | - |
| lung lavage | - | - | - | - | - | - |
| mediastinal LN | - | - | ||||
| mesenteric LN | - | - | - | - | - | - |
| mid trachea | - | - | - | - | - | - |
| muscle | - | -- | - | - | ||
| Peyer’s patch | - | - | - | - | ||
| olfactory bulb | - | - | ||||
| rectum | - | - | - | - | ||
| retropharyngeal LN | - | - | - | - | - | - |
| right cerebellum | - | - | - | - | - | - |
| seminal vesicles | - | - | ||||
| sm/lg intestines | - | - | - | - | - | |
| spleen | - | - | - | - | - | |
| testicle | - | - | - | - | ||
| thymus | - | - | - | - | ||
| tonsil | - | - | - | - | - | - |
| tracheal wash | - | - | - | - | - | - |
| tracheobronchial LN | + | - | - | - | - | - |
| turbinates | - | - | - | - | - | - |
| urinary bladder | - | - | - | - | ||
| vas deferens | - | - | - | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falkenberg, S.; Buckley, A.; Laverack, M.; Martins, M.; Palmer, M.V.; Lager, K.; Diel, D.G. Experimental Inoculation of Young Calves with SARS-CoV-2. Viruses 2021, 13, 441. https://doi.org/10.3390/v13030441
Falkenberg S, Buckley A, Laverack M, Martins M, Palmer MV, Lager K, Diel DG. Experimental Inoculation of Young Calves with SARS-CoV-2. Viruses. 2021; 13(3):441. https://doi.org/10.3390/v13030441
Chicago/Turabian StyleFalkenberg, Shollie, Alexandra Buckley, Melissa Laverack, Mathias Martins, Mitchell V. Palmer, Kelly Lager, and Diego G. Diel. 2021. "Experimental Inoculation of Young Calves with SARS-CoV-2" Viruses 13, no. 3: 441. https://doi.org/10.3390/v13030441
APA StyleFalkenberg, S., Buckley, A., Laverack, M., Martins, M., Palmer, M. V., Lager, K., & Diel, D. G. (2021). Experimental Inoculation of Young Calves with SARS-CoV-2. Viruses, 13(3), 441. https://doi.org/10.3390/v13030441