
Measuring the Humoral Immune Response in Cats Exposed to Feline Leukaemia Virus

 ,
,  , and
, and 
Abstract
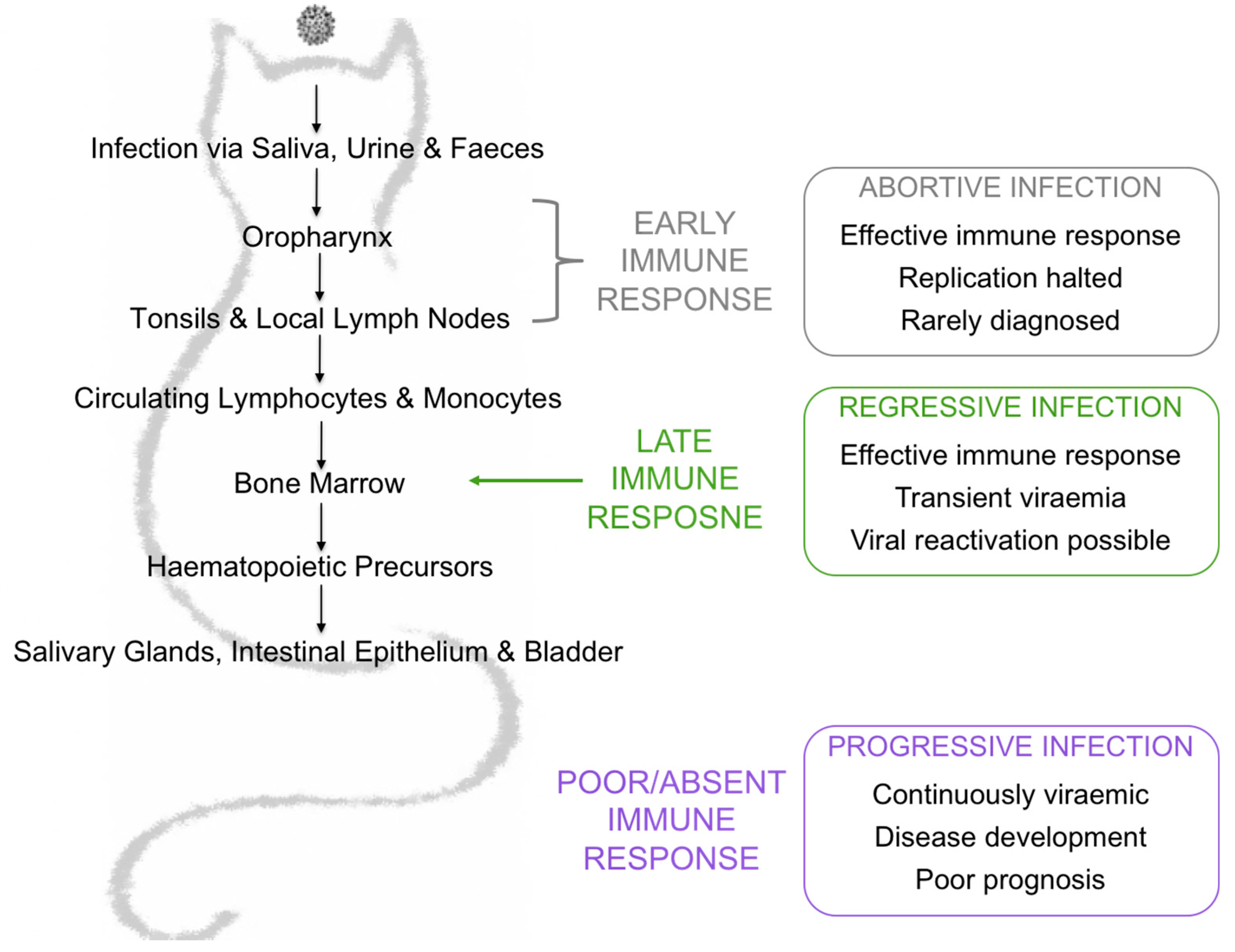
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Detection of Plasma Antigenaemia
2.3. Virus Isolation from PBMC
2.4. Detection of Reverse Transcriptase in PBMC Culture Fluids
2.5. Detection of p27 Capsid Antigen in PBMC Culture Fluids
2.6. Detection of FeLV Proviral DNA in PBMC
2.7. Live Virus Neutralisation Assay
2.8. Preparation of FeLV-A Immunoblots
2.9. FeLV-A Immunoblot Analysis
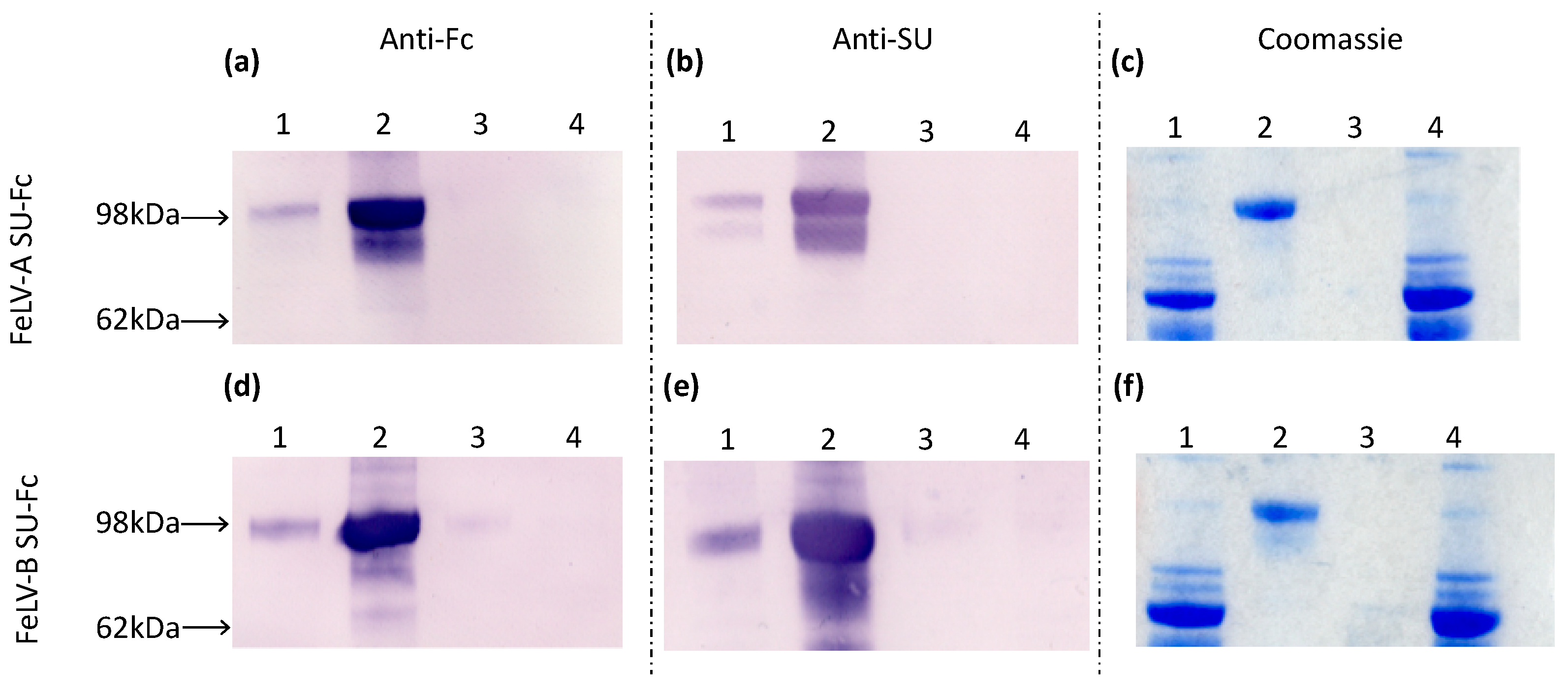
2.10. Production of SU Fusion Proteins
2.11. Testing Plasma Samples for Reactivity against FeLV-SU
2.12. Statistics
3. Results
3.1. Production of FeLV-SU Proteins
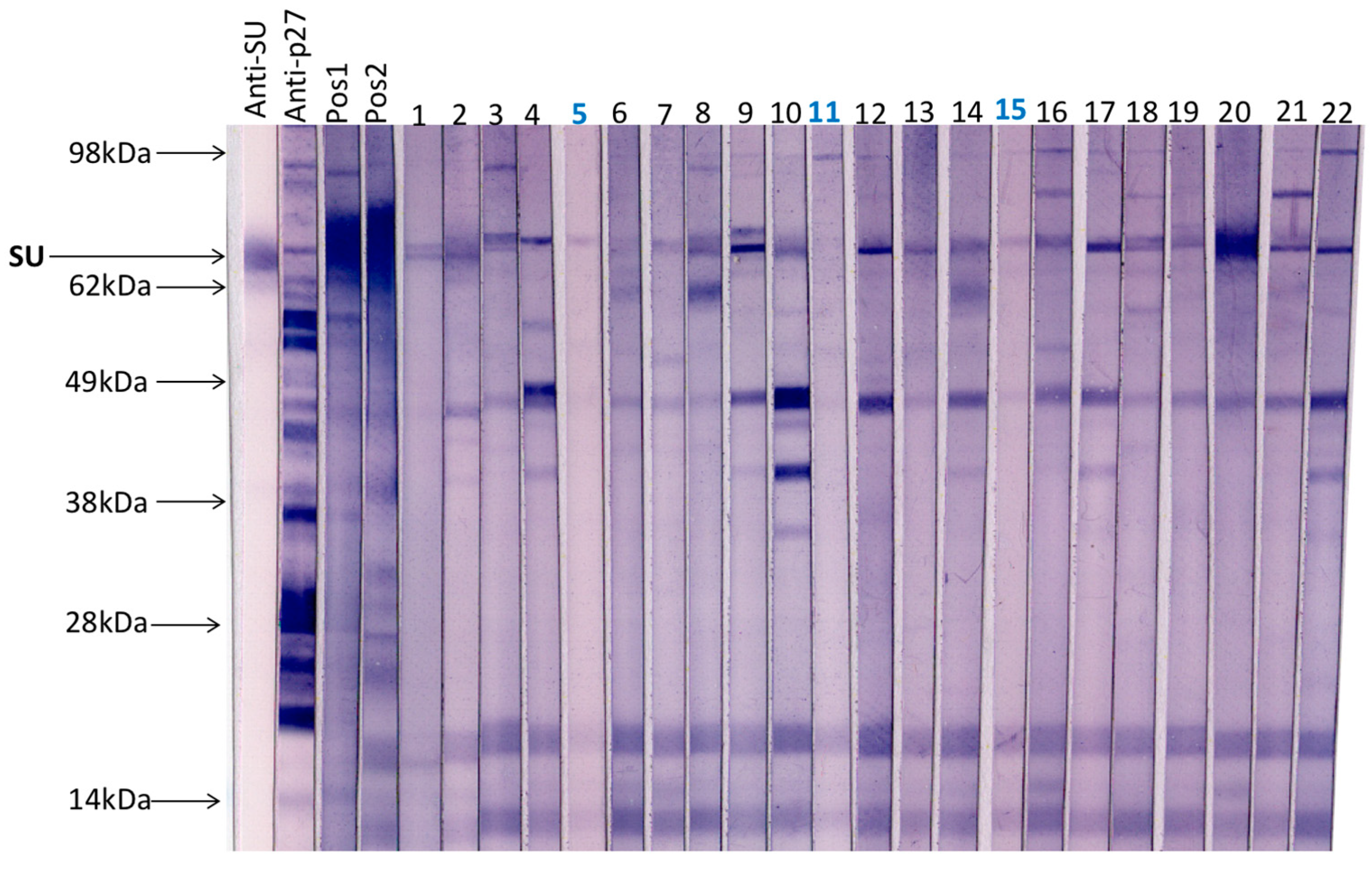
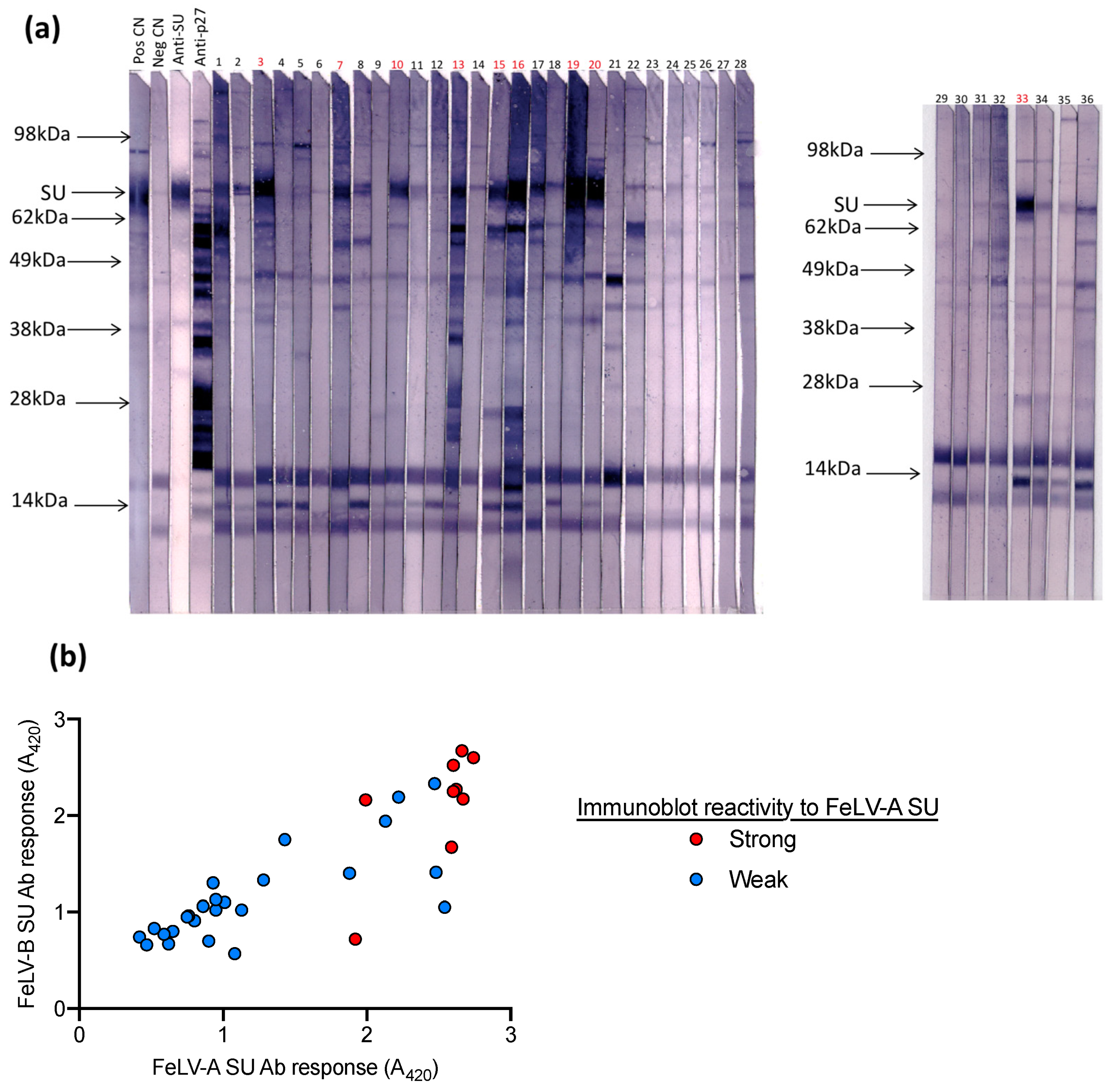
3.2. Immunoblot Analysis of Field Samples and Samples from FeLV Naïve SPF Cats
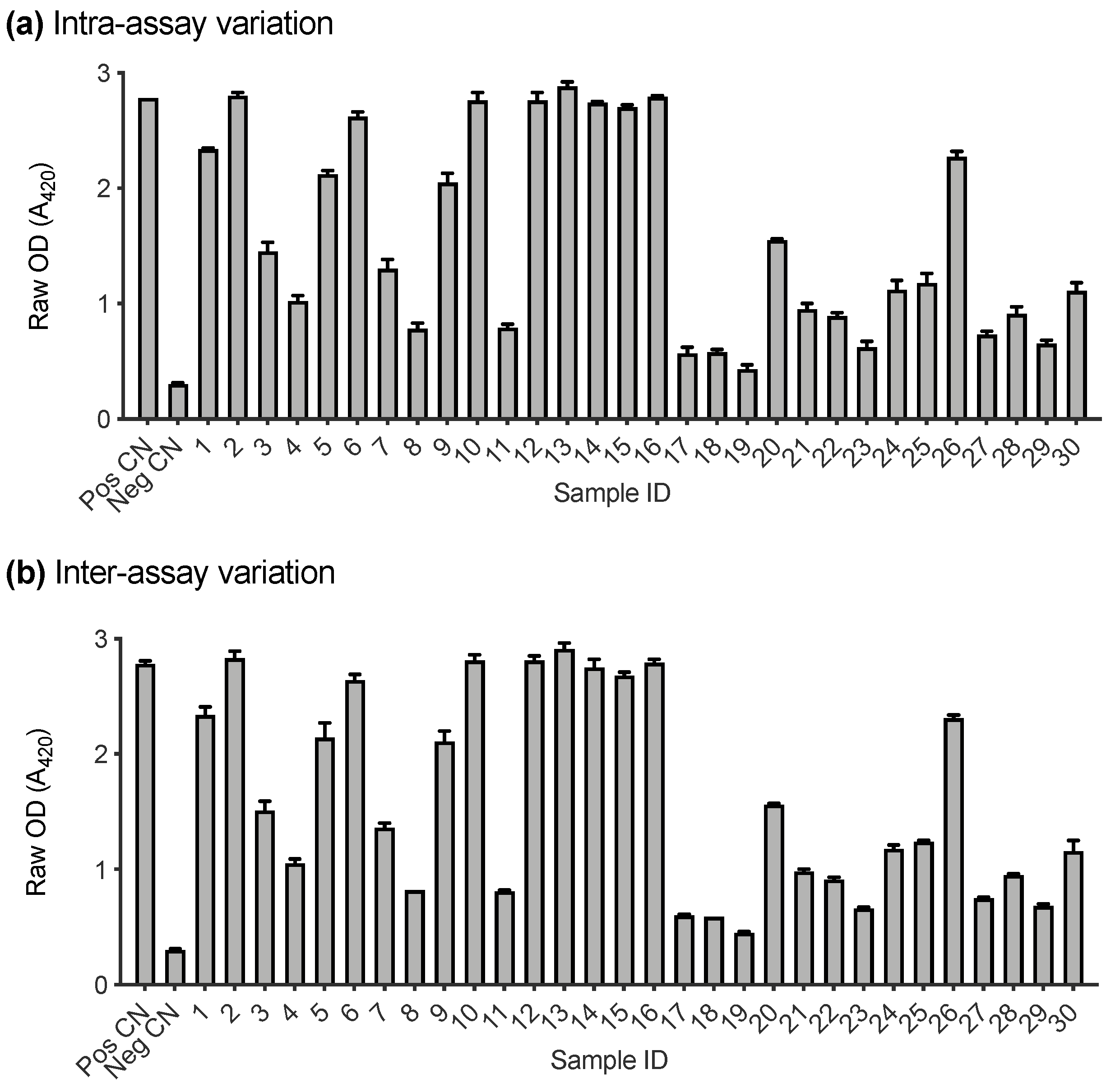
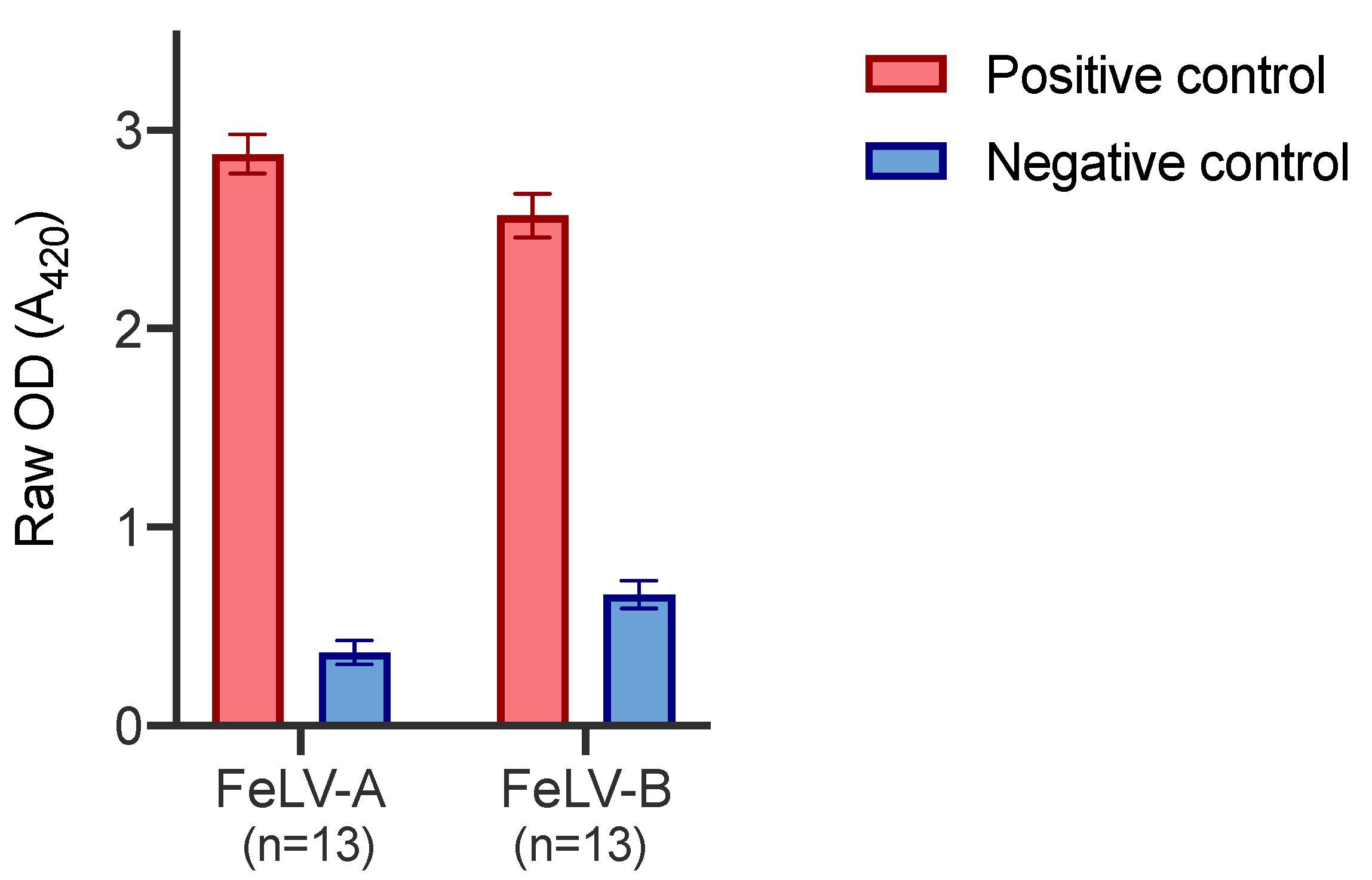
3.3. Assay Performance Analysis
3.4. Assigning Exposure Outcomes
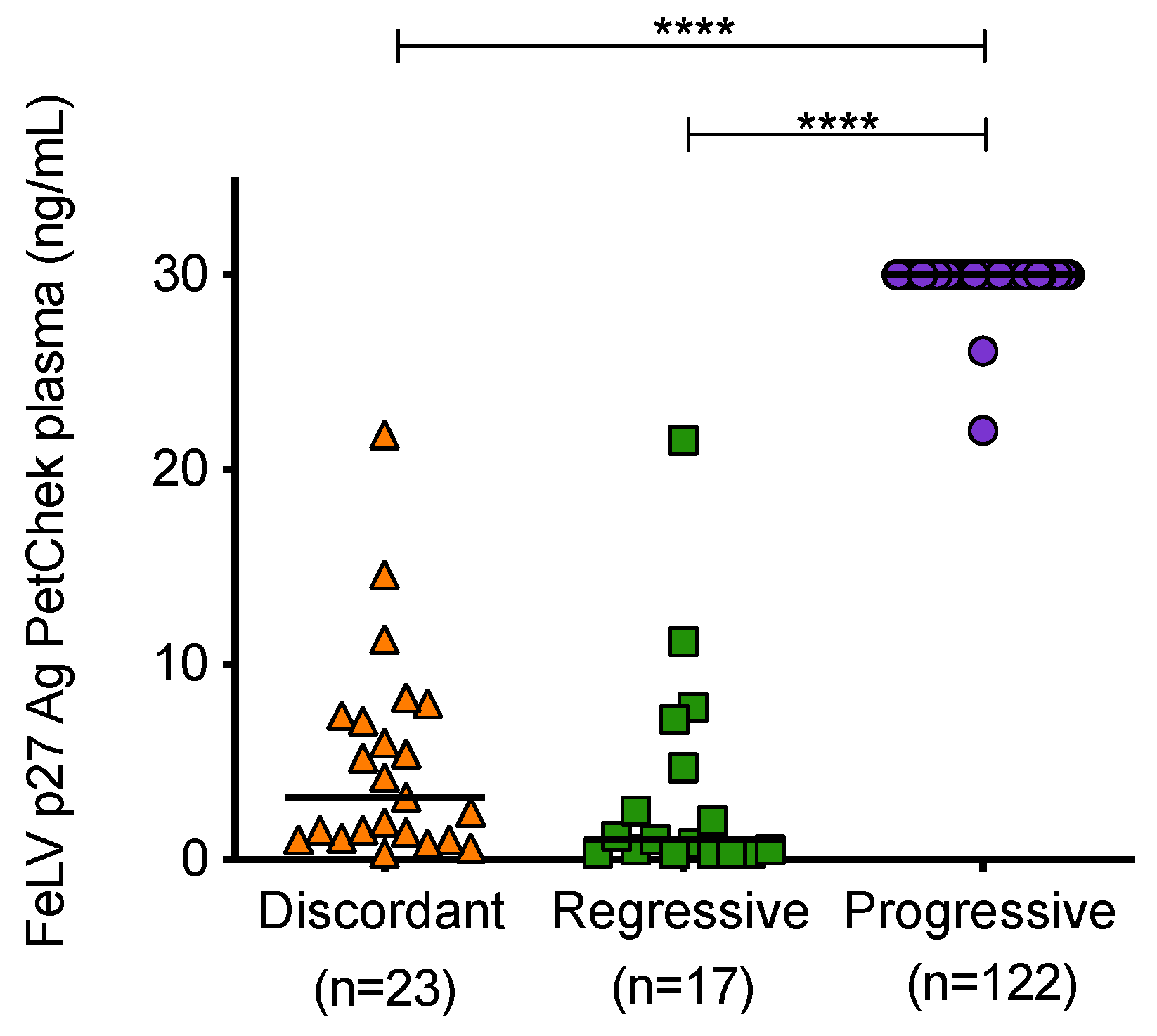
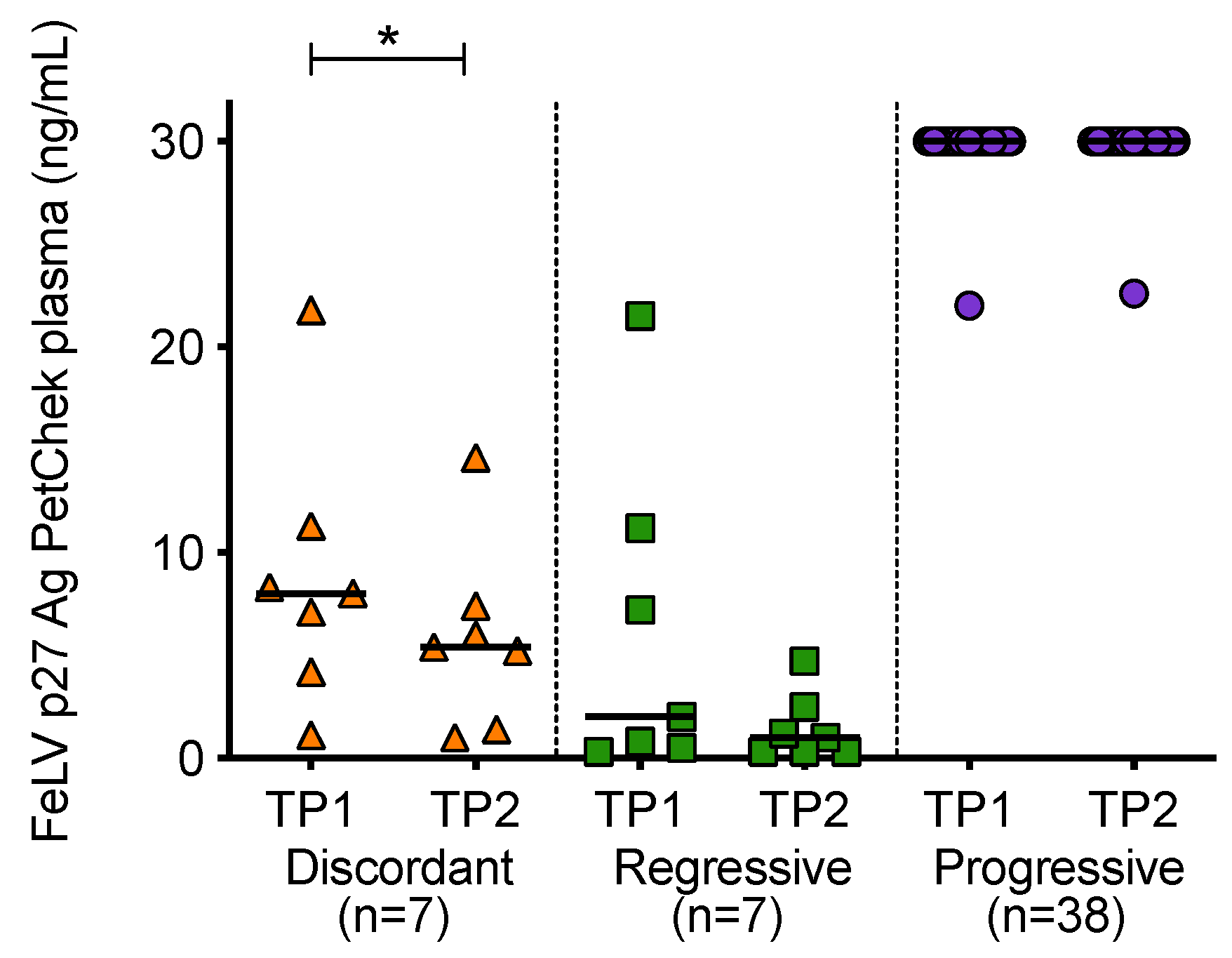
3.5. Antigenaemia in Different FeLV Exposure Outcome Groups
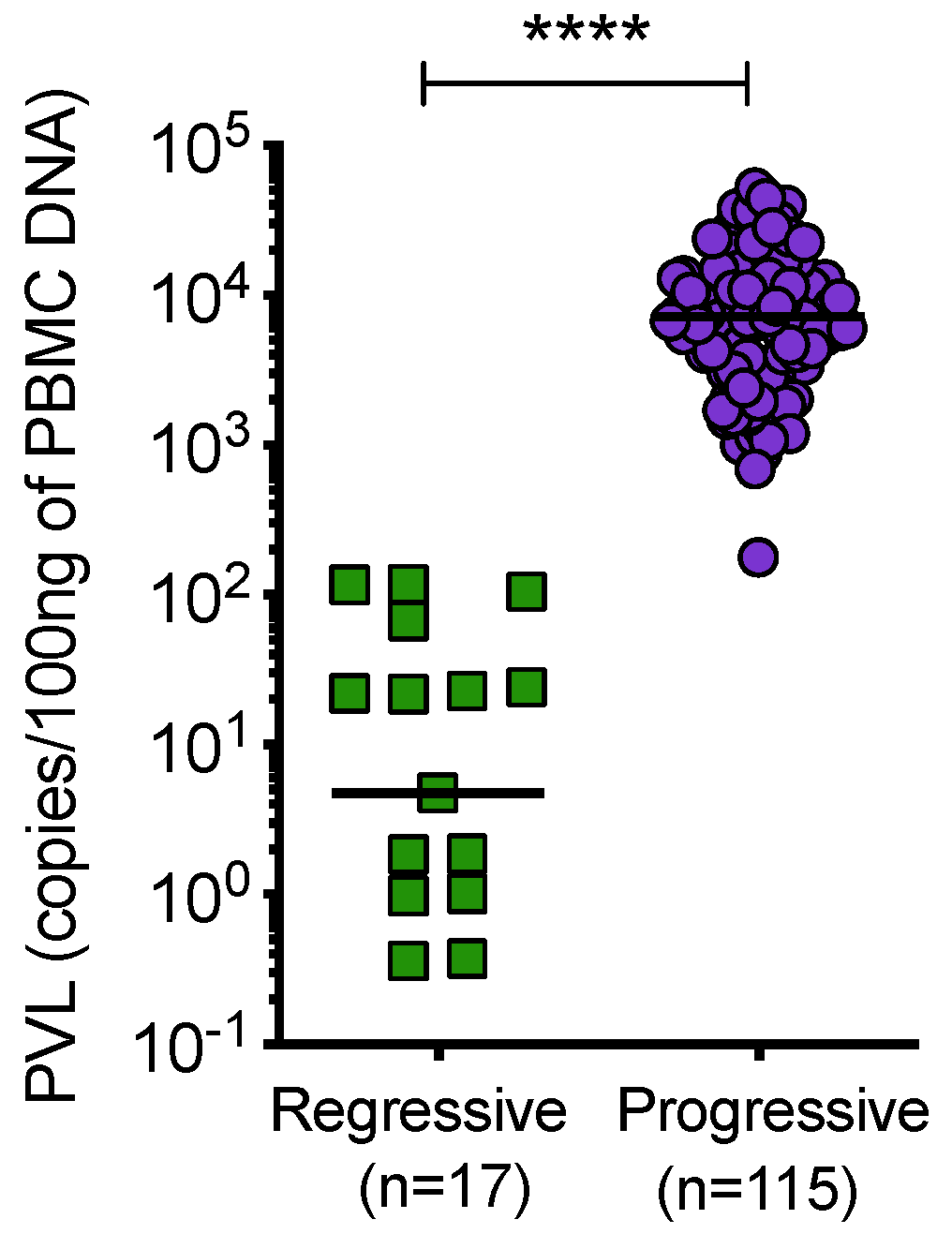
3.6. Proviral Load of Samples from Cats with Regressive and Progressive Infection
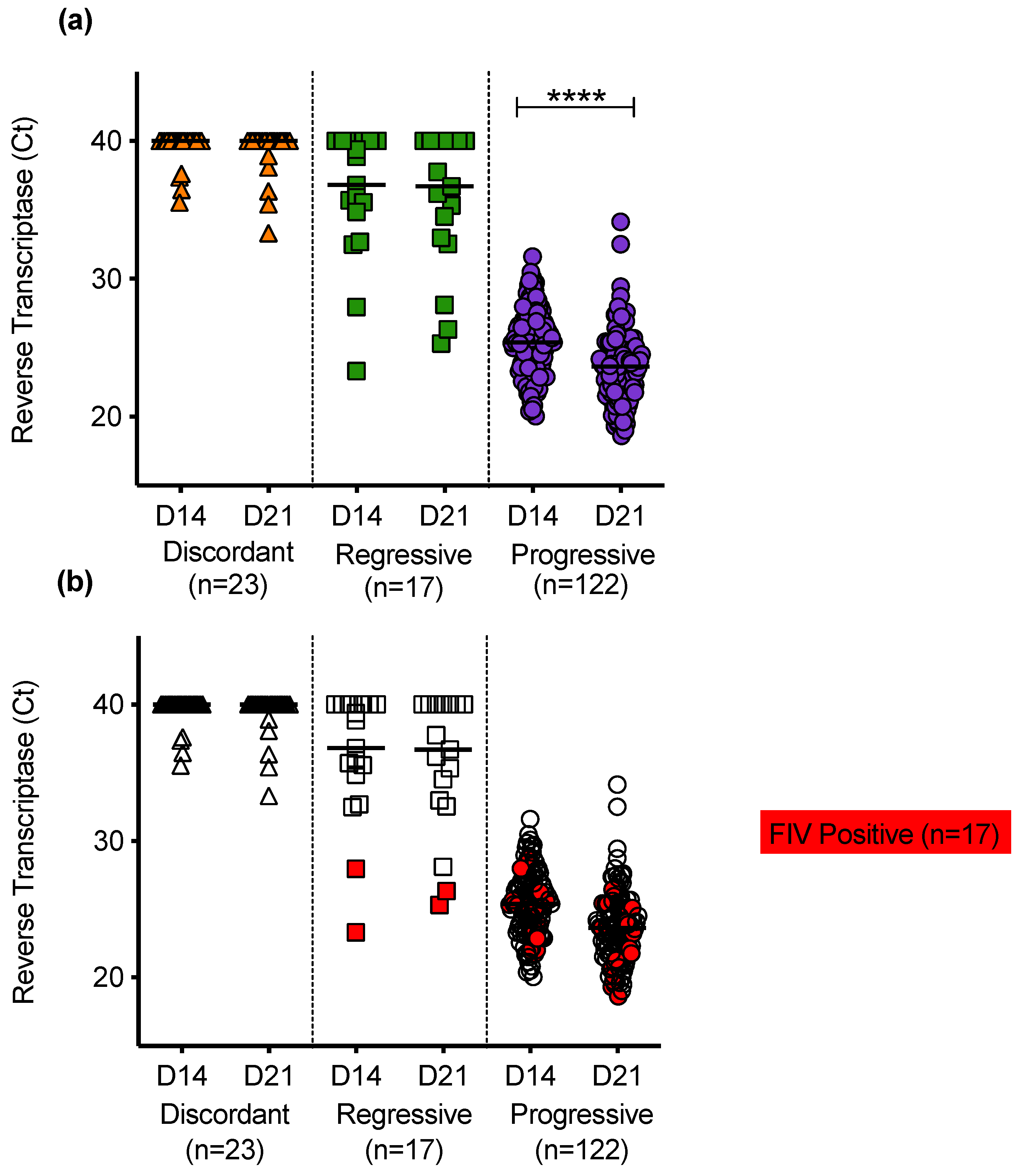
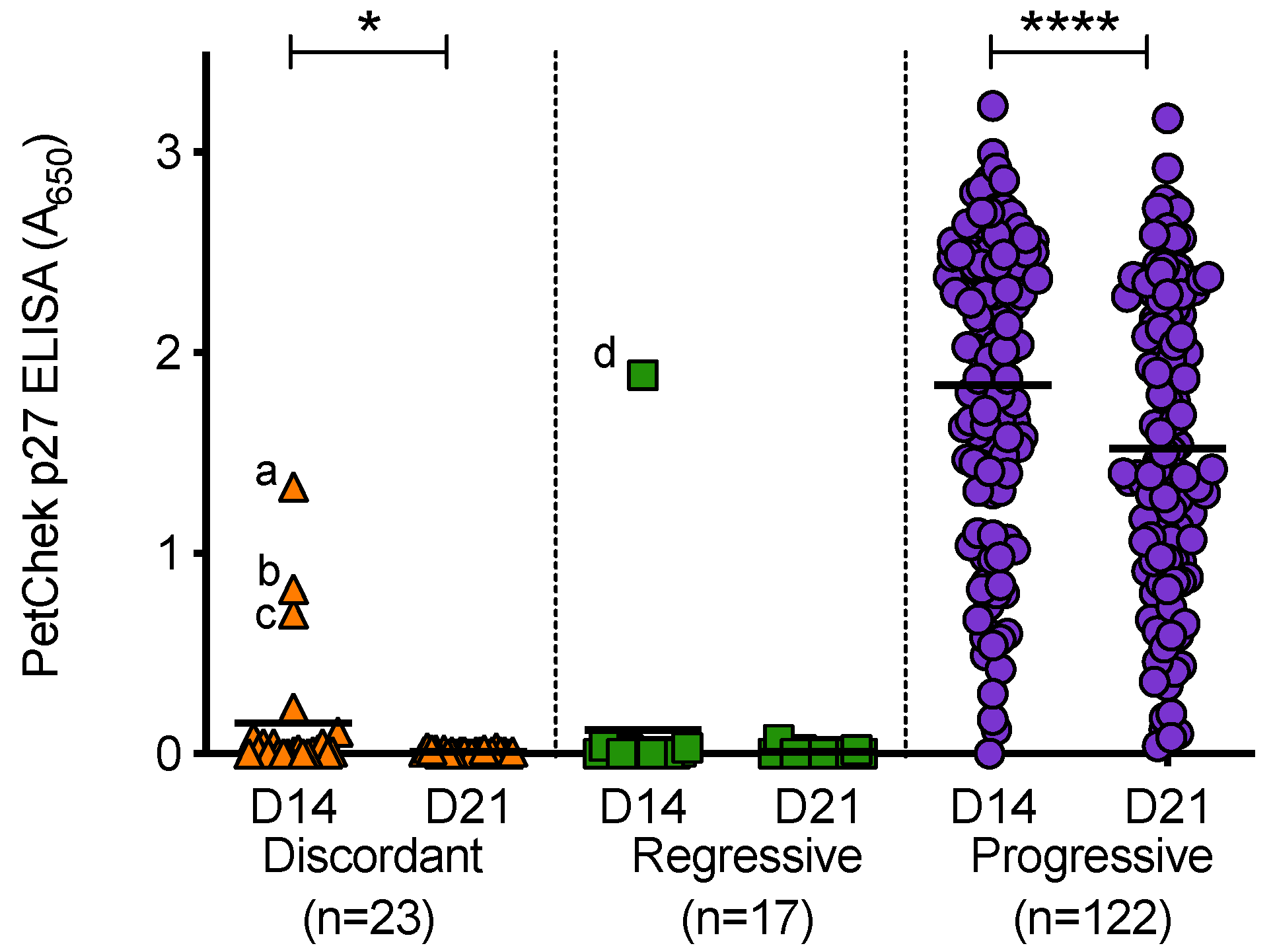
3.7. Analysis of PBMC Culture Fluids for p27 Capsid Antigen and RT
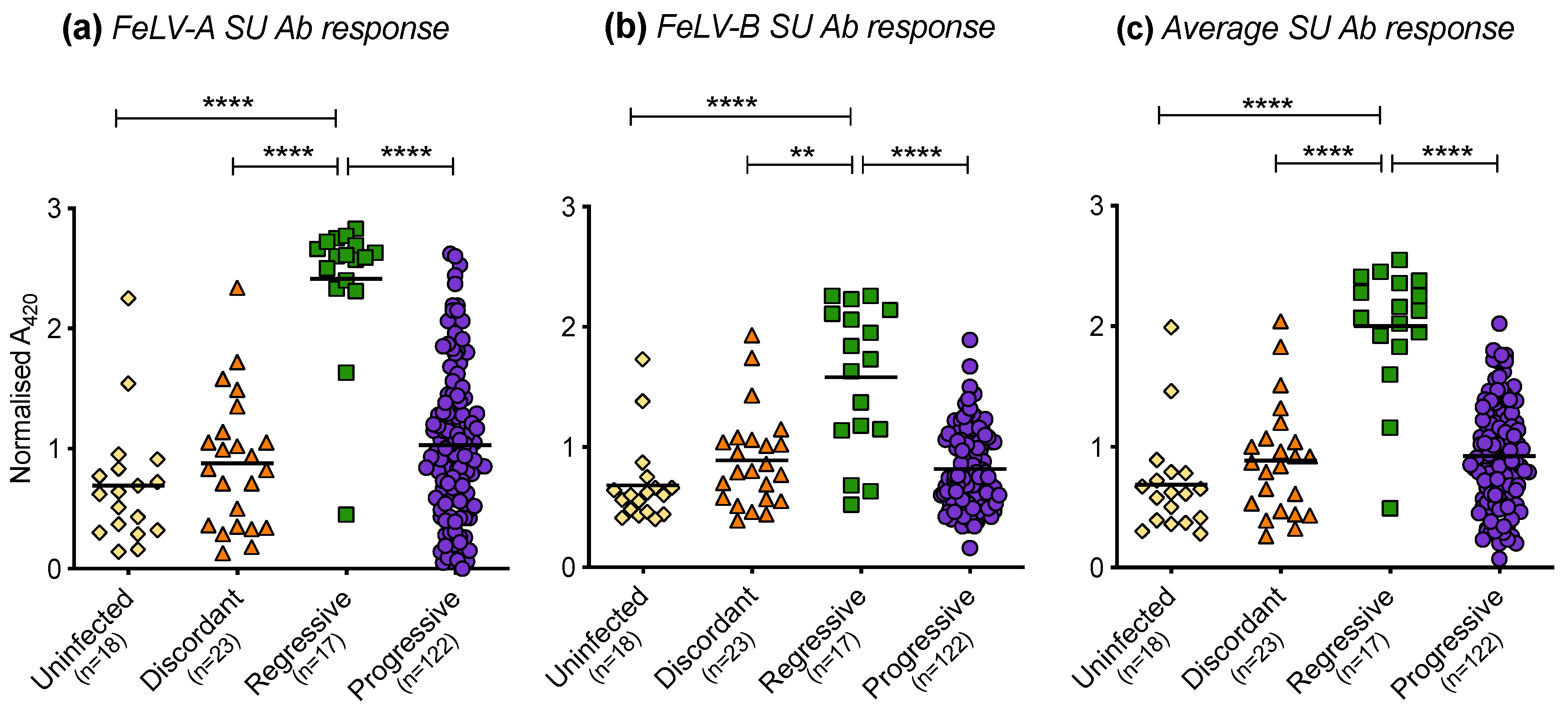
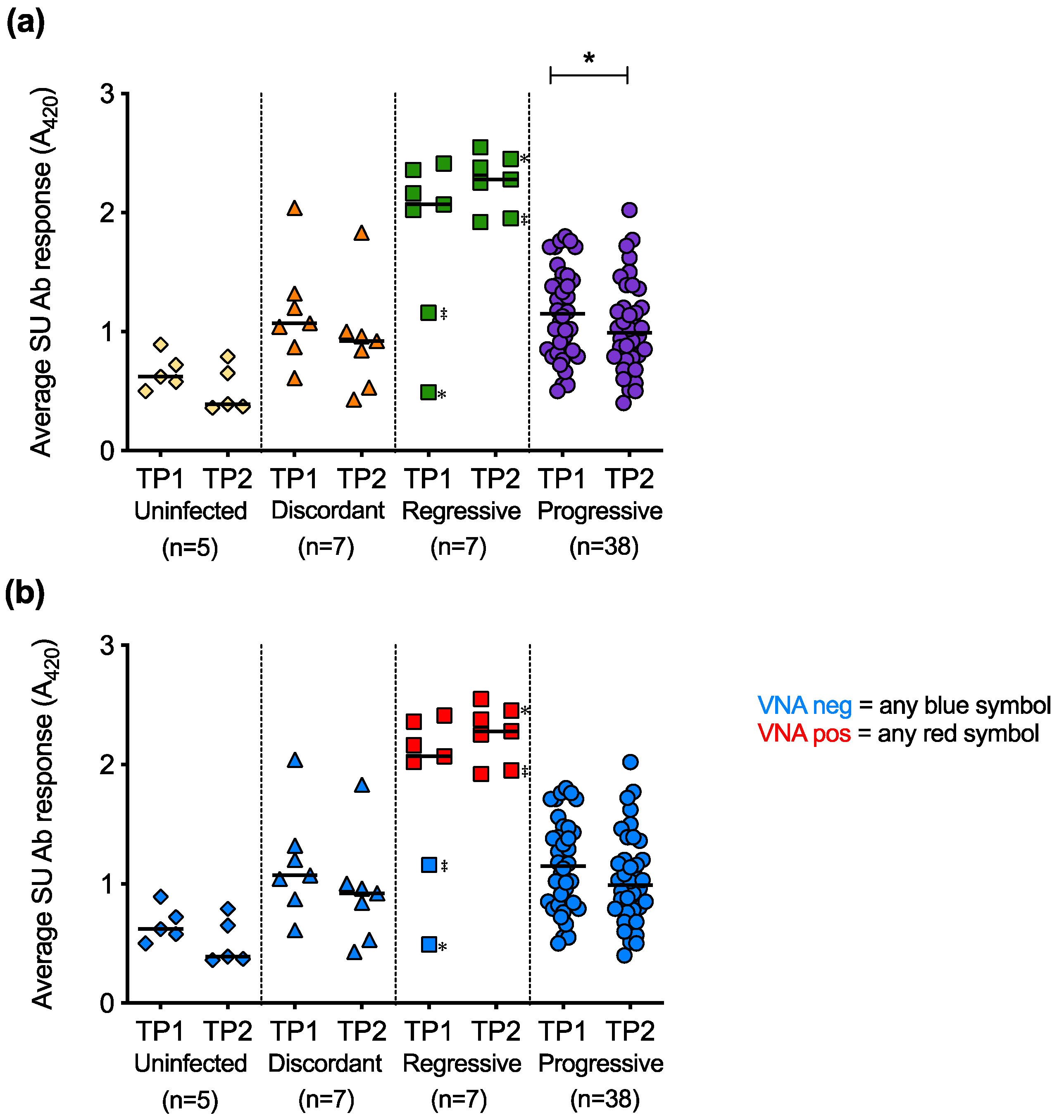
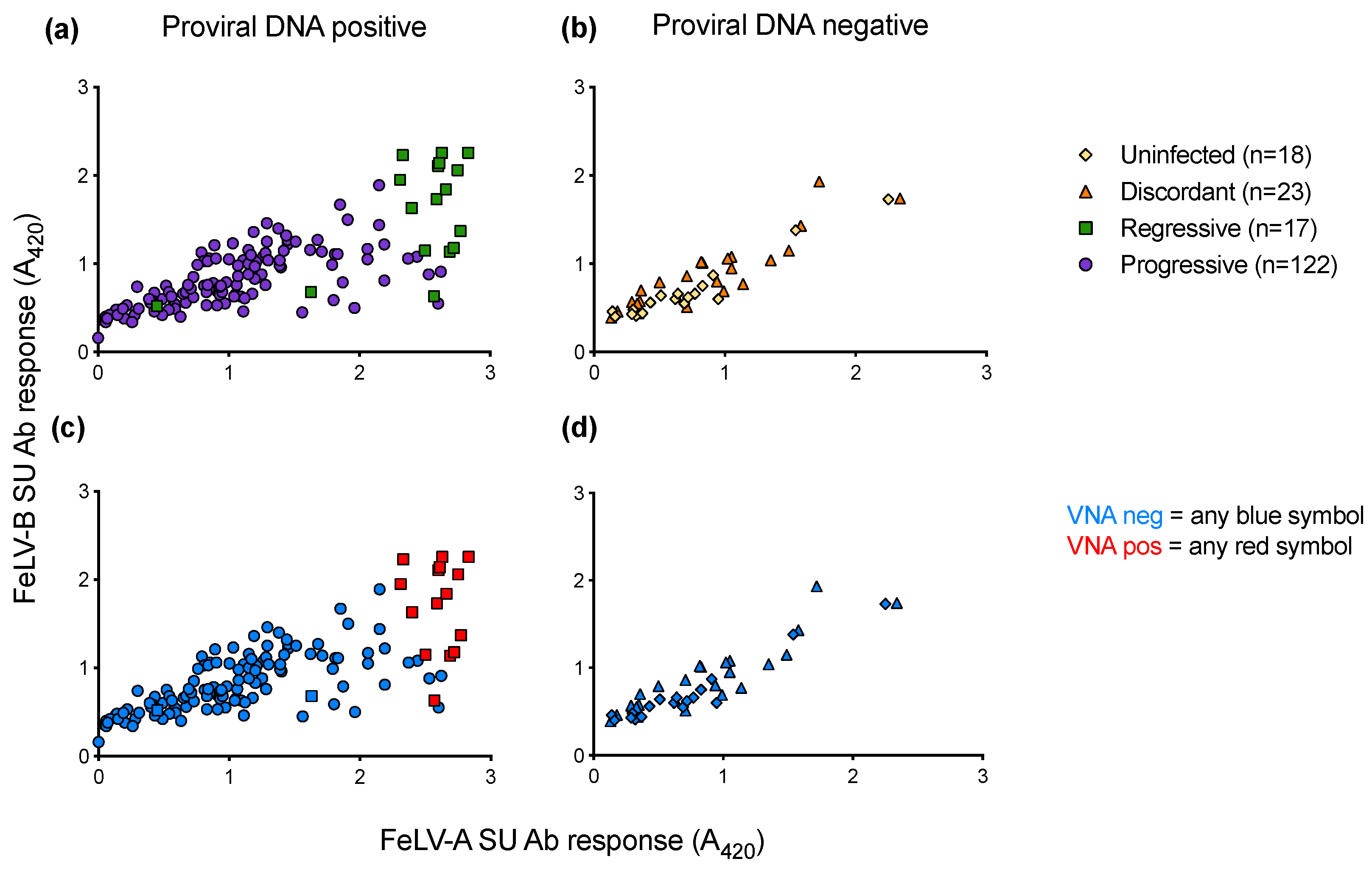
3.8. Humoral Immune Response of Cats with Different FeLV Exposure Outcomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hardy, W.D.; Hess, P.W.; MacEwen, E.G.; McClelland, A.J.; Zuckerman, E.E.; Essex, M.; Cotter, S.M.; Jarrett, O. Biology of feline leukemia virus in the natural environment. Cancer Res. 1976, 36, 582–588. [Google Scholar] [PubMed]
- Hoover, E.; Schaller, J.; Mathes, L.E.; Olsen, R. Passive immunity to feline leukemia: Evaluation of immunity from dams naturally infected and experimentally vaccinated. Infect. Immun. 1977, 16, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Cattori, V.; Tandon, R.; Riond, B.; Pepin, A.C.; Lutz, H.; Hofmann-Lehmann, R. The kinetics of feline leukaemia virus shedding in experimentally infected cats are associated with infection outcome. Vet. Microbiol 2009, 133, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Keller, M.A.; Gonczi, E.; Grenacher, B.; Tandon, R.; Hofman-Lehmann, R.; Lutz, H. Fecal shedding of infectious feline leukemia virus and its nucleic acids: A transmission potential. Vet. Microbiol. 2009, 134, 208–217. [Google Scholar] [CrossRef]
- Lutz, H.; Addie, D.; Belak, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; et al. Feline leukaemia. ABCD guidelines on prevention and management. J. Feline Med. Surg. 2009, 11, 565–574. [Google Scholar] [CrossRef]
- Burling, A.N.; Levy, J.K.; Scott, H.M.; Crandall, M.M.; Tucker, S.J.; Wood, E.G.; Foster, J.D. Seroprevalences of feline leukemia virus and feline immunodeficiency virus infection in cats in the United States and Canada and risk factors for seropositivity. J. Am. Vet. Med. Assoc. 2017, 251, 187–194. [Google Scholar] [CrossRef]
- Stewart, H.; Jarrett, O.; Hosie, M.J.; Willett, B.J. Are endogenous feline leukemia viruses really endogenous? Vet. Immunol. Immunopathol. 2011, 143, 325–331. [Google Scholar] [CrossRef]
- Schaller, J.P.; Mathes, L.E.; Hoover, E.A.; Koestner, A.; Olsen, R.G. Increased susceptibility to feline leukemia virus infection in cats exposed to methylnitrosourea. Cancer Res. 1978, 38, 996–998. [Google Scholar]
- Bolin, L.L.; Ahmad, S.; Levy, L.S. The surface glycoprotein of a natural feline leukemia virus subgroup A variant, FeLV-945, as a determinant of disease outcome. Vet. Immunol. Immunopathol. 2011, 143, 221–226. [Google Scholar] [CrossRef]
- Jarrett, O.; Neil, J.C. Feline Leukaemia Virus. In eLS; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012. [Google Scholar] [CrossRef]
- Sykes, J.E.; Hartmann, K. Feline Leukemia Virus Infection. In Canine and Feline Infectious Diseases; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 224–238. [Google Scholar] [CrossRef]
- Torres, A.N.; Mathiason, C.K.; Hoover, E.A. Re-examination of feline leukemia virus: Host relationships using real-time PCR. Virology 2005, 332, 272–283. [Google Scholar] [CrossRef]
- Major, A.; Cattori, V.; Boenzli, E.; Riond, B.; Ossent, P.; Meli, M.L.; Hofmann-Lehmann, R.; Lutz, H. Exposure of cats to low doses of FeLV: Seroconversion as the sole parameter of infection. Vet. Res. 2010, 41, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.; Lorentzen, L.; Shields, J.; Lewis, H. Long-term outcome of cats with natural FeLV and FIV infection. In Proceedings of the 8th International Feline Retrovirus Research Symposium, Washington, DC, USA, 8–11 October 2006; pp. 8–13. [Google Scholar]
- Hartmann, K. Clinical aspects of feline retroviruses: A review. Viruses 2012, 4, 2684–2710. [Google Scholar] [CrossRef]
- Lutz, H.; Jarrett, O. Detection of feline leukemia virus infection in saliva. J. Clin. Microbiol. 1987, 25, 827–831. [Google Scholar] [CrossRef]
- Little, S.; Levy, J.; Hartmann, K.; Hofmann-Lehmann, R.; Hosie, M.; Olah, G.; Denis, K.S. 2020 AAFP feline retrovirus testing and management guidelines. J. Feline Med. Surg. 2020, 22, 5–30. [Google Scholar] [CrossRef] [PubMed]
- Hofmann-Lehmann, R.; Hartmann, K. Feline leukaemia virus infection: A practical approach to diagnosis. J. Feline Med. Surg. 2020, 22, 831–846. [Google Scholar] [CrossRef] [PubMed]
- Lutz, H.; Pedersen, N.; Higgins, J.; Hübscher, U.; Troy, F.A.; Theilen, G.H. Humoral immune reactivity to feline leukemia virus and associated antigens in cats naturally infected with feline leukemia virus. Cancer Res. 1980, 40, 3642–3651. [Google Scholar]
- Jarrett, J. Serology in feline leukaemia virus infections (correspondence). Vet. Rec. 1981, 108, 465. [Google Scholar] [CrossRef]
- Jarrett, O.; Russell, P.; Stewart, M. Protection of kittens from feline leukaemia virus infection by maternally-derived antibody. Vet. Rec. 1977, 101, 304–305. [Google Scholar] [CrossRef]
- Flynn, J.N.; Dunham, S.P.; Watson, V.; Jarrett, O. Longitudinal analysis of feline leukemia virus-specific cytotoxic T lymphocytes: Correlation with recovery from infection. J. Virol. 2002, 76, 2306–2315. [Google Scholar] [CrossRef] [PubMed]
- Dunham, S.P.; Graham, E. Retroviral infections of small animals. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 879–901. [Google Scholar] [CrossRef]
- Buch, J.S.; Clark, G.H.; Cahill, R.; Thatcher, B.; Smith, P.; Chandrashekar, R.; Leutenegger, C.M.; O’Connor, T.P.; Beall, M.J. Analytical validation of a reference laboratory ELISA for the detection of feline leukemia virus p27 antigen. J. Vet. Diagn. Investig. 2017, 29, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Beall, M.J.; Buch, J.; Cahill, R.J.; Clark, G.; Hanscom, J.; Estrada, M.; Leutenegger, C.M.; Chandrashekar, R. Evaluation of a quantitative enzyme-linked immunosorbent assay for feline leukemia virus p27 antigen and comparison to proviral DNA loads by real-time polymerase chain reaction. Comp. Immunol. Microbiol. Infect. Dis. 2019, 67, 101348. [Google Scholar] [CrossRef]
- Chang, A.; Ostrove, J.M.; Bird, R.E. Development of an improved product enhanced reverse transcriptase assay. J. Virol. Methods 1997, 65, 45–54. [Google Scholar] [CrossRef]
- Arnold, B.A.; Hepler, R.W.; Keller, P.M. One-step fluorescent probe product-enhanced reverse transcriptase assay. Biotechniques 1998, 25, 98–106. [Google Scholar] [CrossRef]
- Lovatt, A. Applications of quantitative PCR in the biosafety and genetic stability assessment of biotechnology products. Rev. Mol. Biotechnol. 2002, 82, 279–300. [Google Scholar] [CrossRef]
- Cattori, V.; Tandon, R.; Pepin, A.; Lutz, H.; Hofmann-Lehmann, R. Rapid detection of feline leukemia virus provirus integration into feline genomic DNA. Mol. Cell. Probes 2006, 20, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, J.; Jarrett, O.; Neil, J.; Desmettre, P.; Paoletti, E. Protection of cats against feline leukemia virus by vaccination with a canarypox virus recombinant, ALVAC-FL. J. Virol. 1993, 67, 2370–2375. [Google Scholar] [CrossRef] [PubMed]
- Noronha, F.; Dougherty, E.; Poco, A.; Gries, C.; Post, J.; Rickard, C. Cytological and serological studies of a feline endogenous C-type virus. Arch. für die Gesamte Virusforsch. 1974, 45, 235–248. [Google Scholar] [CrossRef]
- Spiller, O.B.; Mark, L.; Blue, C.E.; Proctor, D.G.; Aitken, J.A.; Blom, A.M.; Blackbourn, D.J. Dissecting the regions of virion-associated Kaposi’s sarcoma-associated herpesvirus complement control protein required for complement regulation and cell binding. J. Virol. 2006, 80, 4068–4078. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Englert, T.; Lutz, H.; Sauter-Louis, C.; Hartmann, K. Survey of the feline leukemia virus infection status of cats in Southern Germany. J. Feline Med. Surg. 2012, 14, 392–398. [Google Scholar] [CrossRef]
- Boenzli, E.; Hadorn, M.; Hartnack, S.; Huder, J.; Hofmann-Lehmann, R.; Lutz, H. Detection of antibodies to the feline leukemia virus (FeLV) transmembrane protein p15E: An alternative approach for serological FeLV detection based on antibodies to p15E. J. Clin. Microbiol. 2014, 52, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Hofmann-Lehmann, R.; Huder, J.B.; Gruber, S.; Boretti, F.; Sigrist, B.; Lutz, H. Feline leukaemia provirus load during the course of experimental infection and in naturally infected cats. J. Gen. Virol. 2001, 82, 1589–1596. [Google Scholar] [CrossRef]
- Beall, M.J.; Buch, J.; Clark, G.; Estrada, M.; Rakitin, A.; Hamman, N.T.; Frenden, M.K.; Jefferson, E.P.; Amirian, E.S.; Levy, J.K. Feline Leukemia Virus p27 Antigen Concentration and Proviral DNA Load Are Associated with Survival in Naturally Infected Cats. Viruses 2021, 13, 302. [Google Scholar] [CrossRef] [PubMed]
- Westman, M.; Norris, J.; Malik, R.; Hofmann-Lehmann, R.; Parr, Y.A.; Armstrong, E.; McDonald, M.; Hall, E.; Sheehy, P.; Hosie, M.J. Anti-SU Antibody Responses in Client-Owned Cats Following Vaccination against Feline Leukaemia Virus with Two Inactivated Whole-Virus Vaccines (Fel-O-Vax® Lv-K and Fel-O-Vax® 5). Viruses 2021, 13, 240. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Turnaround Time | Hours | Hours | 1 Week | 5 Days |
|---|---|---|---|---|
| Outcome of Exposure | p27 Capsid Antigen | Proviral DNA | Plasma Virus Isolation | Virus Neutralising Antibody |
| Abortive infection | Negative | Negative | Negative | Positive |
| Regressive infection (shedding) | Positive | Positive | Positive | Negative |
| Regressive infection (recovered) | Negative | Positive | Negative | Positive |
| Progressive infection | Positive | Positive | Positive | Negative |
| Focal infection | Positive | Positive (*) | Negative | Positive |
| Months between Timepoints | Number of Cats |
|---|---|
| 1 | 1 |
| 2 | 0 |
| 3 | 5 |
| 4 | 5 |
| 5 | 3 |
| 6 | 43 |
| Outcome of Exposure | p27 Capsid Antigen | PBMC Proviral DNA | PBMC Virus Isolation | Total No. of Cats | No. of Cats with Single Sample | No. of Cats with Two Samples | Total No. of Samples |
|---|---|---|---|---|---|---|---|
| Uninfected | Negative | Negative | Negative | 13 | 8 | 5 | 18 |
| Discordant | Positive | Negative | Negative | 16 | 9 | 7 | 23 |
| Regressive | Positive/Negative | Positive (*) | Negative | 10 | 3 | 7 | 17 |
| Progressive | Positive | Positive | Positive | 84 | 46 | 38 | 122 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parr, Y.A.; Beall, M.J.; Levy, J.K.; McDonald, M.; Hamman, N.T.; Willett, B.J.; Hosie, M.J. Measuring the Humoral Immune Response in Cats Exposed to Feline Leukaemia Virus. Viruses 2021, 13, 428. https://doi.org/10.3390/v13030428
Parr YA, Beall MJ, Levy JK, McDonald M, Hamman NT, Willett BJ, Hosie MJ. Measuring the Humoral Immune Response in Cats Exposed to Feline Leukaemia Virus. Viruses. 2021; 13(3):428. https://doi.org/10.3390/v13030428
Chicago/Turabian StyleParr, Yasmin A., Melissa J. Beall, Julie K. Levy, Michael McDonald, Natascha T. Hamman, Brian J. Willett, and Margaret J. Hosie. 2021. "Measuring the Humoral Immune Response in Cats Exposed to Feline Leukaemia Virus" Viruses 13, no. 3: 428. https://doi.org/10.3390/v13030428
APA StyleParr, Y. A., Beall, M. J., Levy, J. K., McDonald, M., Hamman, N. T., Willett, B. J., & Hosie, M. J. (2021). Measuring the Humoral Immune Response in Cats Exposed to Feline Leukaemia Virus. Viruses, 13(3), 428. https://doi.org/10.3390/v13030428