
Growth Kinetics of Influenza C Virus Antigenic Mutants That Escaped from Anti-Hemagglutinin Esterase Monoclonal Antibodies and Viral Antigenic Changes Found in Field Isolates

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Hemagglutination Inhibition Test
2.3. Nucleotide Sequencing
2.4. Structural Analysis
2.5. Viral Growth Kinetics
3. Results
3.1. Antigenic Properties of 12 Escape Mutants
3.2. Growth Kinetics of the Escape Mutants in Cultured Cells
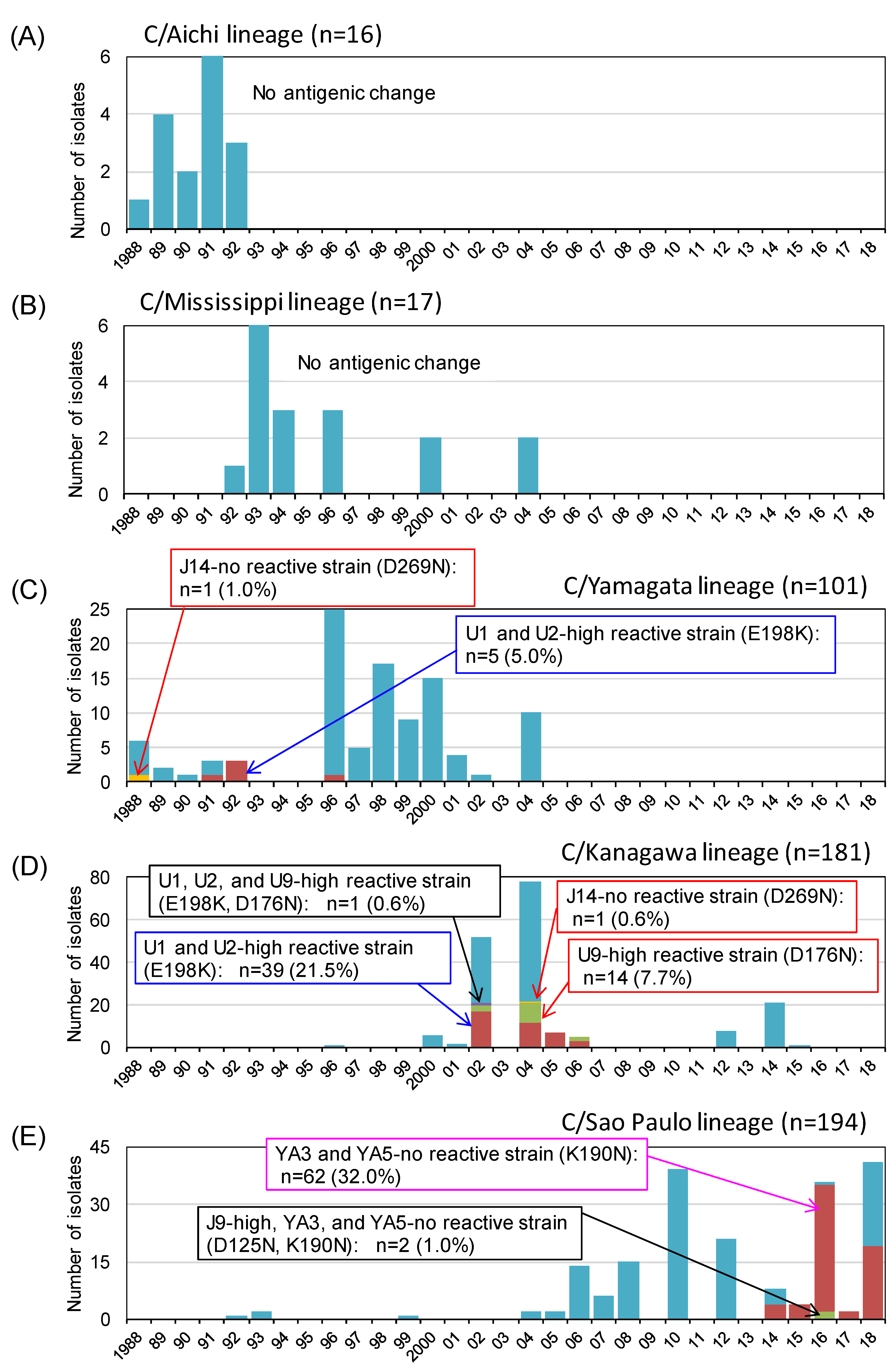
3.3. Antigenic Changes among the Natural Isolates of Influenza C Virus in Japan between 1988 and 2018
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taylor, R.M. Studies on survival of influenza virus between epidemics and antigenic variants of the virus. Am. J. Public Health Nations Health 1949, 39, 171–178. [Google Scholar] [CrossRef]
- Thielen, B.K.; Friedlander, H.; Bistodeau, S.; Shu, B.; Lynch, B.; Martin, K.; Bye, E.; Como-Sabetti, K.; Boxrud, D.; Strain, A.K.; et al. Detection of Influenza C Viruses Among Outpatients and Patients Hospitalized for Severe Acute Respiratory Infection, Minnesota, 2013–2016. Clin. Infect. Dis. 2018, 66, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Daniels, R.S.; Tse, H.; Ermetal, B.; Xiang, Z.; Jackson, D.J.; Guntoro, J.; Nicod, J.; Stewart, A.; Cross, K.J.; Hussain, S.; et al. Molecular Characterization of Influenza C Viruses from Outbreaks in Hong Kong SAR, China. J. Virol. 2020, 94, e01051-20. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, Y.; Katsushima, N.; Nagai, Y.; Shoji, M.; Itagaki, T.; Sakamoto, M.; Kitaoka, S.; Mizuta, K.; Nishimura, H. Clinical features of influenza C virus infection in children. J. Infect. Dis. 2006, 193, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Odagiri, T.; Matsuzaki, Y.; Okamoto, M.; Suzuki, A.; Saito, M.; Tamaki, R.; Lupisan, S.P.; Sombrero, L.T.; Hongo, S.; Oshitani, H. Isolation and characterization of influenza C viruses in the Philippines and Japan. J. Clin. Microbiol. 2015, 53, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Abiko, C.; Ikeda, T.; Mizuta, K.; Matsuzaki, Y. Influenza C virus and human metapneumovirus infections in hospitalized children with lower respiratory tract illness. Pediatr. Infect. Dis. J. 2015, 34, 1273–1275. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Sugawara, K.; Abiko, C.; Ikeda, T.; Aoki, Y.; Mizuta, K.; Katsushima, N.; Katsushima, F.; Katsushima, Y.; Itagaki, T.; et al. Epidemiological information regarding the periodic epidemics of influenza C virus in Japan (1996–2013) and the seroprevalence of antibodies to different antigenic groups. J. Clin. Virol. 2014, 61, 87–93. [Google Scholar] [CrossRef]
- Liu, R.; Sheng, Z.; Lin, T.; Sreenivasan, C.; Gao, R.; Thomas, M.; Druce, J.; Hause, B.M.; Kaushik, R.S.; Li, F.; et al. Genetic and antigenic characteristics of a human influenza C virus clinical isolate. J. Med. Virol. 2020, 92, 161–166. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Sugawara, K.; Furuse, Y.; Shimotai, Y.; Hongo, S.; Oshitani, H.; Mizuta, K.; Nishimura, H. Genetic Lineage and Reassortment of Influenza C Viruses Circulating between 1947 and 2014. J. Virol. 2016, 90, 8251–8265. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Muraki, Y.; Sugawara, K.; Hongo, S.; Nishimura, H.; Kitame, F.; Katsushima, N.; Numazaki, Y.; Nakamura, K. Cocirculation of two distinct groups of influenza C virus in Yamagata City, Japan. Virology 1994, 202, 796–802. [Google Scholar] [CrossRef]
- Kimura, H.; Abiko, C.; Peng, G.; Muraki, Y.; Sugawara, K.; Hongo, S.; Kitame, F.; Mizuta, K.; Numazaki, Y.; Suzuki, H.; et al. Interspecies transmission of influenza C virus between humans and pigs. Virus Res. 1997, 48, 71–79. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Mizuta, K.; Kimura, H.; Sugawara, K.; Tsuchiya, E.; Suzuki, H.; Hongo, S.; Nakamura, K. Characterization of antigenically unique influenza C virus strains isolated in Yamagata and Sendai cities, Japan, during 1992–1993. J. Gen. Virol. 2000, 81 Pt 6, 1447–1452. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Sugawara, K.; Mizuta, K.; Tsuchiya, E.; Muraki, Y.; Hongo, S.; Suzuki, H.; Nakamura, K. Antigenic and genetic characterization of influenza C viruses which caused two outbreaks in Yamagata City, Japan, in 1996 and 1998. J. Clin. Microbiol. 2002, 40, 422–429. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Mizuta, K.; Sugawara, K.; Tsuchiya, E.; Muraki, Y.; Hongo, S.; Suzuki, H.; Nishimura, H. Frequent reassortment among influenza C viruses. J. Virol. 2003, 77, 871–881. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Takao, S.; Shimada, S.; Mizuta, K.; Sugawara, K.; Takashita, E.; Muraki, Y.; Hongo, S.; Nishimura, H. Characterization of antigenically and genetically similar influenza C viruses isolated in Japan during the 1999–2000 season. Epidemiol Infect. 2004, 132, 709–720. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Sato, K.; Sugawara, K.; Takashita, E.; Muraki, Y.; Morishita, T.; Kumagai, N.; Suzuki, S.; Hongo, S. Isolation of an influenza C virus introduced into Japan by a traveler from Malaysia. J. Clin. Microbiol. 2005, 43, 993–995. [Google Scholar] [CrossRef][Green Version]
- Matsuzaki, Y.; Abiko, C.; Mizuta, K.; Sugawara, K.; Takashita, E.; Muraki, Y.; Suzuki, H.; Mikawa, M.; Shimada, S.; Sato, K.; et al. A nationwide epidemic of influenza C virus infection in Japan in 2004. J. Clin. Microbiol. 2007, 45, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Aoki, Y.; Matoba, Y.; Yahagi, K.; Mizuta, K.; Itagaki, T.; Katsushima, F.; Katsushima, Y.; Matsuzaki, Y. The dominant antigenic group of influenzaCinfections changed from C/Sao Paulo/378/82-lineage to C/Kanagawa/1/76-lineage in Yamagata, Japan, in 2014. Jpn J. Infect. Dis. 2015, 68, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, Y.; Shimotai, Y.; Kadowaki, Y.; Sugawara, K.; Hongo, S.; Mizuta, K.; Nishimura, H. Antigenic changes among the predominantly circulating C/Sao Paulo lineage strains of influenza C virus in Yamagata, Japan, between 2015 and 2018. Infect. Genet Evol. 2020, 81, 104269. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, M.; Sugawara, K.; Adachi, K.; Hongo, S.; Nishimura, H.; Kitame, F.; Nakamura, K. Location of neutralizing epitopes on the hemagglutinin-esterase protein of influenza C virus. Virology 1992, 189, 79–87. [Google Scholar] [CrossRef]
- Pfeifer, J.B.; Compans, R.W. Structure of the influenza C glycoprotein gene as determined from cloned DNA. Virus Res. 1984, 1, 281–296. [Google Scholar] [CrossRef]
- Vlasak, R.; Krystal, M.; Nacht, M.; Palese, P. The influenza C virus glycoprotein (HE) exhibits receptor-binding (hemagglutinin) and receptor-destroying (esterase) activities. Virology 1987, 160, 419–425. [Google Scholar] [CrossRef]
- Herrler, G.; Dürkop, I.; Becht, H.; Klenk, H.D. The glycoprotein of influenza C virus is the haemagglutinin, esterase and fusion factor. J. Gen. Virol. 1988, 69 Pt 4, 839–846. [Google Scholar] [CrossRef]
- Formanowski, F.; Meier-Ewert, H. Isolation of the influenza C virus glycoprotein in a soluble form by bromelain digestion. Virus Res. 1988, 10, 177–191. [Google Scholar] [CrossRef]
- Rosenthal, P.B.; Zhang, X.; Formanowski, F.; Fitz, W.; Wong, C.H.; Meier-Ewert, H.; Skehel, J.J.; Wiley, D.C. Structure of the haemagglutinin-esterase-fusion glycoprotein of influenza C virus. Nature 1998, 396, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, Y.; Sugawara, K.; Furuse, Y.; Shimotai, Y.; Hongo, S.; Mizuta, K.; Nishimura, H. Neutralizing Epitopes and Residues Mediating the Potential Antigenic Drift of the Hemagglutinin-Esterase Protein of Influenza C Virus. Viruses 2018, 10, 417. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, K.; Nishimura, H.; Hongo, S.; Muraki, Y.; Kitame, F.; Nakamura, K. Construction of an antigenic map of the haemagglutinin-esterase protein of influenza C virus. J. Gen. Virol. 1993, 74 Pt 8, 1661–1666. [Google Scholar] [CrossRef]
- Kawamura, H.; Tashiro, M.; Kitame, F.; Homma, M.; Nakamura, K. Genetic variation among human strains of influenza C virus isolated in Japan. Virus Res. 1986, 4, 275–288. [Google Scholar] [CrossRef]
- Goto, T.; Shimotai, Y.; Matsuzaki, Y.; Muraki, Y.; Sho, R.; Sugawara, K.; Hongo, S. Effect of Phosphorylation of CM2 Protein on Influenza C Virus Replication. J. Virol. 2017, 91, e00773-17. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, K.; Muraki, Y.; Takashita, E.; Matsuzaki, Y.; Hongo, S. Conformational maturation of the nucleoprotein synthesized in influenza C virus-infected cells. Virus Res. 2006, 122, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Furuse, Y.; Matsuzaki, Y.; Nishimura, H.; Oshitani, H. Analyses of Evolutionary Characteristics of the Hemagglutinin-Esterase Gene of Influenza C Virus during a Period of 68 Years Reveals Evolutionary Patterns Different from Influenza A and B Viruses. Viruses 2016, 8, 321. [Google Scholar] [CrossRef] [PubMed]
- Mellquist, J.L.; Kasturi, L.; Spitalnik, S.L.; Shakin-Eshleman, S.H. The amino acid following an asn-X-Ser/Thr sequon is an important determinant of N-linked core glycosylation efficiency. Biochemistry 1998, 37, 6833–6837. [Google Scholar] [CrossRef]
- Sugahara, K.; Hongo, S.; Sugawara, K.; Li, Z.N.; Tsuchiya, E.; Muraki, Y.; Matsuzaki, Y.; Nakamura, K. Role of individual oligosaccharide chains in antigenic properties, intracellular transport, and biological activities of influenza C virus hemagglutinin-esterase protein. Virology 2001, 285, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Potdar, V.A.; Hinge, D.D.; Dakhave, M.R.; Manchanda, A.; Jadhav, N.; Kulkarni, P.B.; Chadha, M.S. Molecular detection and characterization of Influenza ‘C’ viruses from western India. Infect. Genet. Evol. 2017, 54, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Jelley, L.; Levy, A.; Deng, Y.M.; Spirason, N.; Lang, J.; Buettner, I.; Druce, J.; Blyth, C.; Effler, P.; Smith, D.; et al. Influenza C infections in Western Australia and Victoria from 2008 to 2014. Influenza Other Respir Viruses 2016, 10, 455–461. [Google Scholar] [CrossRef]
- Fritsch, A.; Schweiger, B.; Biere, B. Influenza C virus in pre-school children with respiratory infections: Retrospective analysis of data from the national influenza surveillance system in Germany, 2012 to 2014. Eurosurveillance 2019, 24, 1800174. [Google Scholar] [CrossRef] [PubMed]
- Skehel, J.J.; Stevens, D.J.; Daniels, R.S.; Douglas, A.R.; Knossow, M.; Wilson, I.A.; Wiley, D.C. A carbohydrate side chain on hemagglutinins of Hong Kong influenza viruses inhibits recognition by a monoclonal antibody. Proc. Natl. Acad. Sci. USA 1984, 81, 1779–1783. [Google Scholar] [CrossRef]
- Abe, Y.; Takashita, E.; Sugawara, K.; Matsuzaki, Y.; Muraki, Y.; Hongo, S. Effect of the addition of oligosaccharides on the biological activities and antigenicity of influenza A/H3N2 virus hemagglutinin. J. Virol. 2004, 78, 9605–9611. [Google Scholar] [CrossRef] [PubMed]
- Liao, F.; Nishimura, H.; Ito, H.; Zhang, Y.; Matsuzaki, Y. Longitudinal course of influenza C virus antibody titers of healthy adults in Sendai, Japan. J. Clin. Virol. 2020, 133, 104662. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viruses | Amino Acid Substitution | HI Titer of MAbs 1 | HI Titer of Chicken Antiserum against | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A-1 Site | A-2 Site | A-3 Site | Y-1 Site | C/Ann Arbor/1/50 2 | C/Yamagata/ 10/89 3 | |||||||
| J9 | U9 | Q5 | J14 | K16 | U1 | U2 | YA3 | YA5 | ||||
| Wild-type | 128,000 | 128,000 | 64,000 | 256,000 | 80 | 16,000 | 6400 | <20 | <20 | 1280 | 640 | |
| Escape mutant | N173I | 320 | — | — | — | — | — | — | — | — | 2560 | 1280 |
| N175S | <20 | — | — | <20 | — | — | — | — | — | 1280 | 320 | |
| S192L | — | 160 | — | — | — | — | — | 1280 | 1280 | 2560 | 1280 | |
| E193K | — | <20 | <20 | — | <20 | — | — | — | — | 320 | 160 | |
| K235R | — | <20 | <20 | — | — | — | — | — | — | 2560 | 640 | |
| D269N | — | — | — | <20 | <20 | — | — | — | — | 640 | 160 | |
| R68W | — | — | — | — | <20 | — | — | — | — | 2560 | 1280 | |
| A351V | — | — | — | — | <20 | — | — | — | — | 1280 | 640 | |
| L164P | — | — | — | — | — | 160 | <20 | 320 | 80 | 2560 | 1280 | |
| K198E | — | — | — | — | — | 320 | 20 | 40 | 20 | 1280 | 640 | |
| Viruses | Amino Acid Substitution | HI Titer of MAbs 1 | HI Titer of Chicken Antiserum against: | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A-1 Site | A-2 Site | A-3 Site | Y-1 Site | C/Ann Arbor/1/50 2 | C/Yamagata/10/89 3 | |||||||
| J9 | U9 | Q5 | J14 | K16 | U1 | U2 | YA3 | YA5 | ||||
| Wild-type | <20 | 1280 | 12,800 | 32,000 | 320 | 12,800 | 1280 | 12,800 | 12,800 | 320 | 5120 | |
| Escape mutant | ∆192–195 | — | <20 | — | — | <20 | — | — | <20 | <20 | 320 | 2560 |
| ∆198 | — | <20 | 80 | — | <20 | — | — | <20 | <20 | 80 | 640 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuzaki, Y.; Sugawara, K.; Shimotai, Y.; Kadowaki, Y.; Hongo, S.; Mizuta, K.; Nishimura, H. Growth Kinetics of Influenza C Virus Antigenic Mutants That Escaped from Anti-Hemagglutinin Esterase Monoclonal Antibodies and Viral Antigenic Changes Found in Field Isolates. Viruses 2021, 13, 401. https://doi.org/10.3390/v13030401
Matsuzaki Y, Sugawara K, Shimotai Y, Kadowaki Y, Hongo S, Mizuta K, Nishimura H. Growth Kinetics of Influenza C Virus Antigenic Mutants That Escaped from Anti-Hemagglutinin Esterase Monoclonal Antibodies and Viral Antigenic Changes Found in Field Isolates. Viruses. 2021; 13(3):401. https://doi.org/10.3390/v13030401
Chicago/Turabian StyleMatsuzaki, Yoko, Kanetsu Sugawara, Yoshitaka Shimotai, Yoko Kadowaki, Seiji Hongo, Katsumi Mizuta, and Hidekazu Nishimura. 2021. "Growth Kinetics of Influenza C Virus Antigenic Mutants That Escaped from Anti-Hemagglutinin Esterase Monoclonal Antibodies and Viral Antigenic Changes Found in Field Isolates" Viruses 13, no. 3: 401. https://doi.org/10.3390/v13030401
APA StyleMatsuzaki, Y., Sugawara, K., Shimotai, Y., Kadowaki, Y., Hongo, S., Mizuta, K., & Nishimura, H. (2021). Growth Kinetics of Influenza C Virus Antigenic Mutants That Escaped from Anti-Hemagglutinin Esterase Monoclonal Antibodies and Viral Antigenic Changes Found in Field Isolates. Viruses, 13(3), 401. https://doi.org/10.3390/v13030401