
Australian Bat Lyssavirus: Analysis of National Bat Surveillance Data from 2010 to 2016


Abstract
1. Introduction
2. Materials and Methods
2.1. Data, Data Sources and Data Collection
2.2. Data Manipulation and Analysis
3. Results
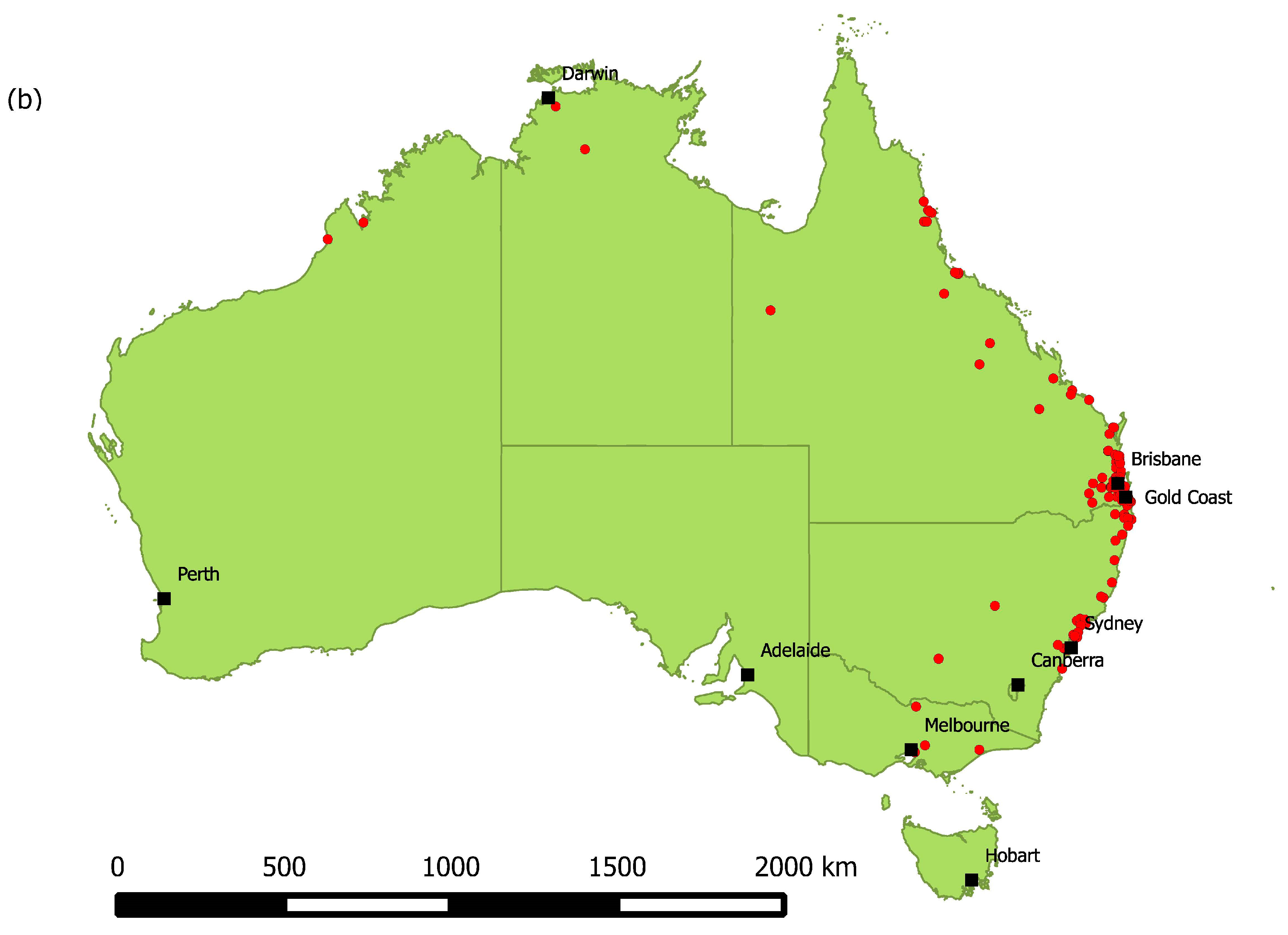
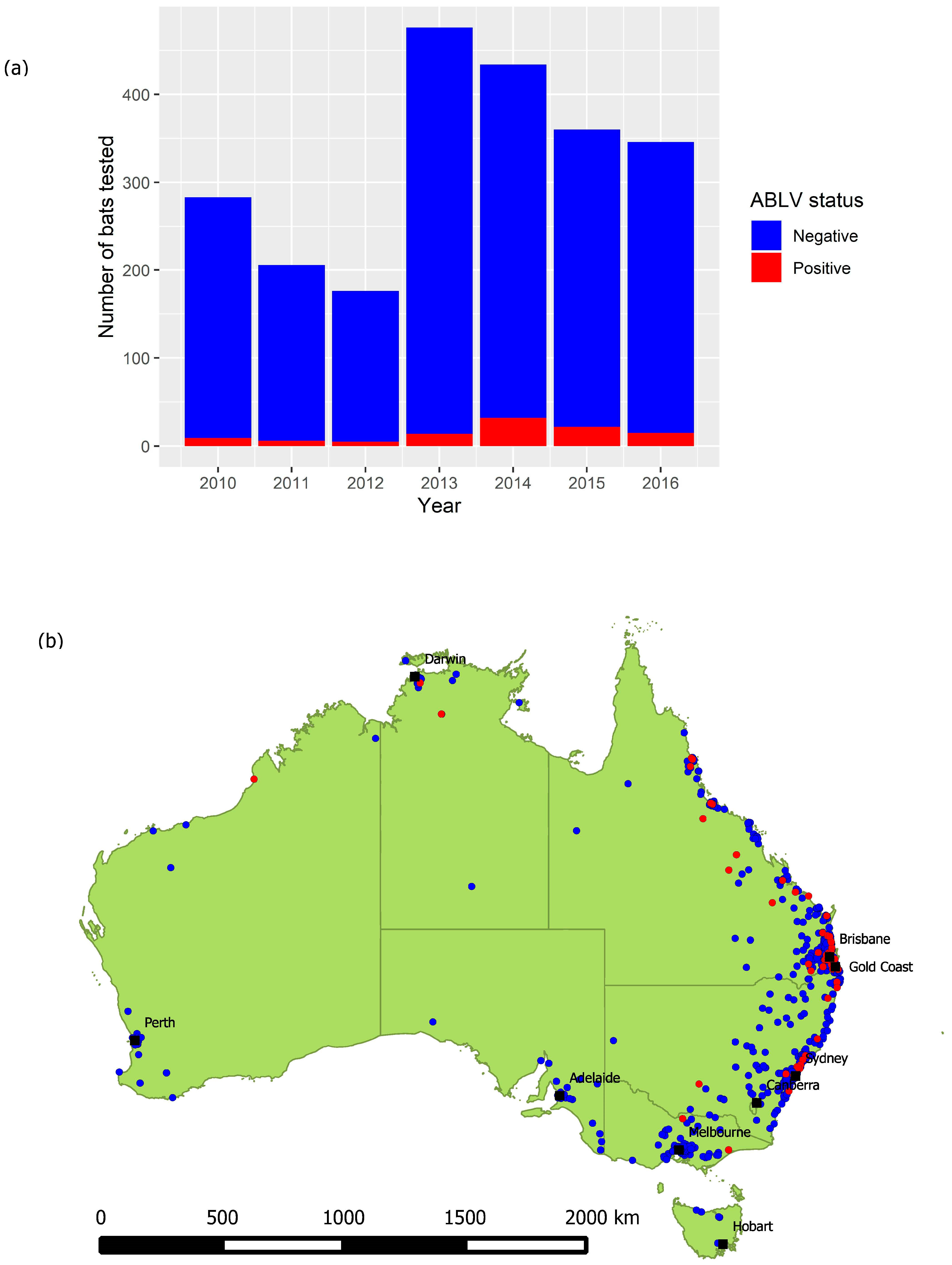
3.1. Risk Factors for ABLV Infection in Flying-Foxes
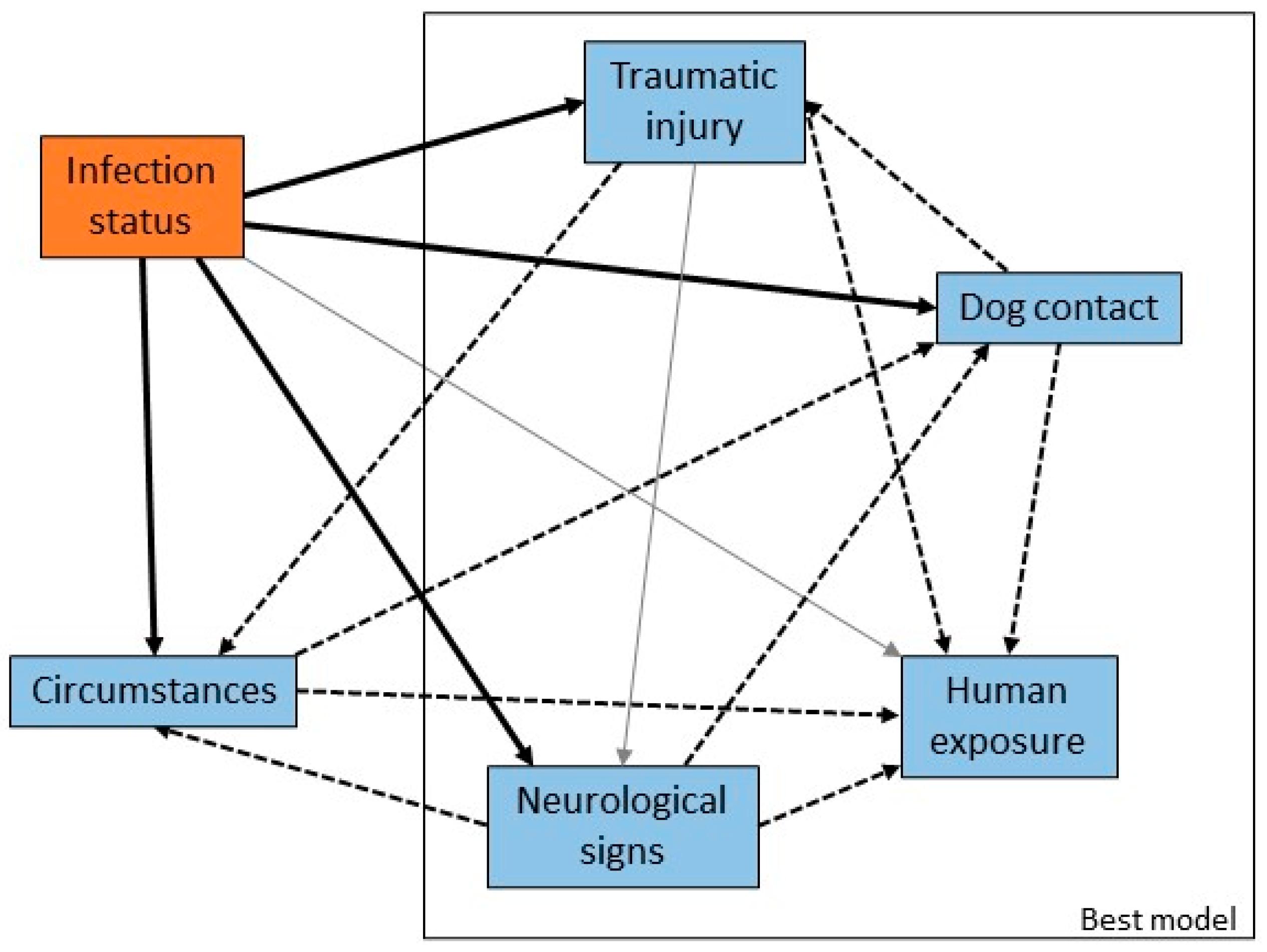
3.2. Circumstances and Clinical Signs in Pteropid Bats
3.3. Risk Factors for Human Contact with Flying-Foxes
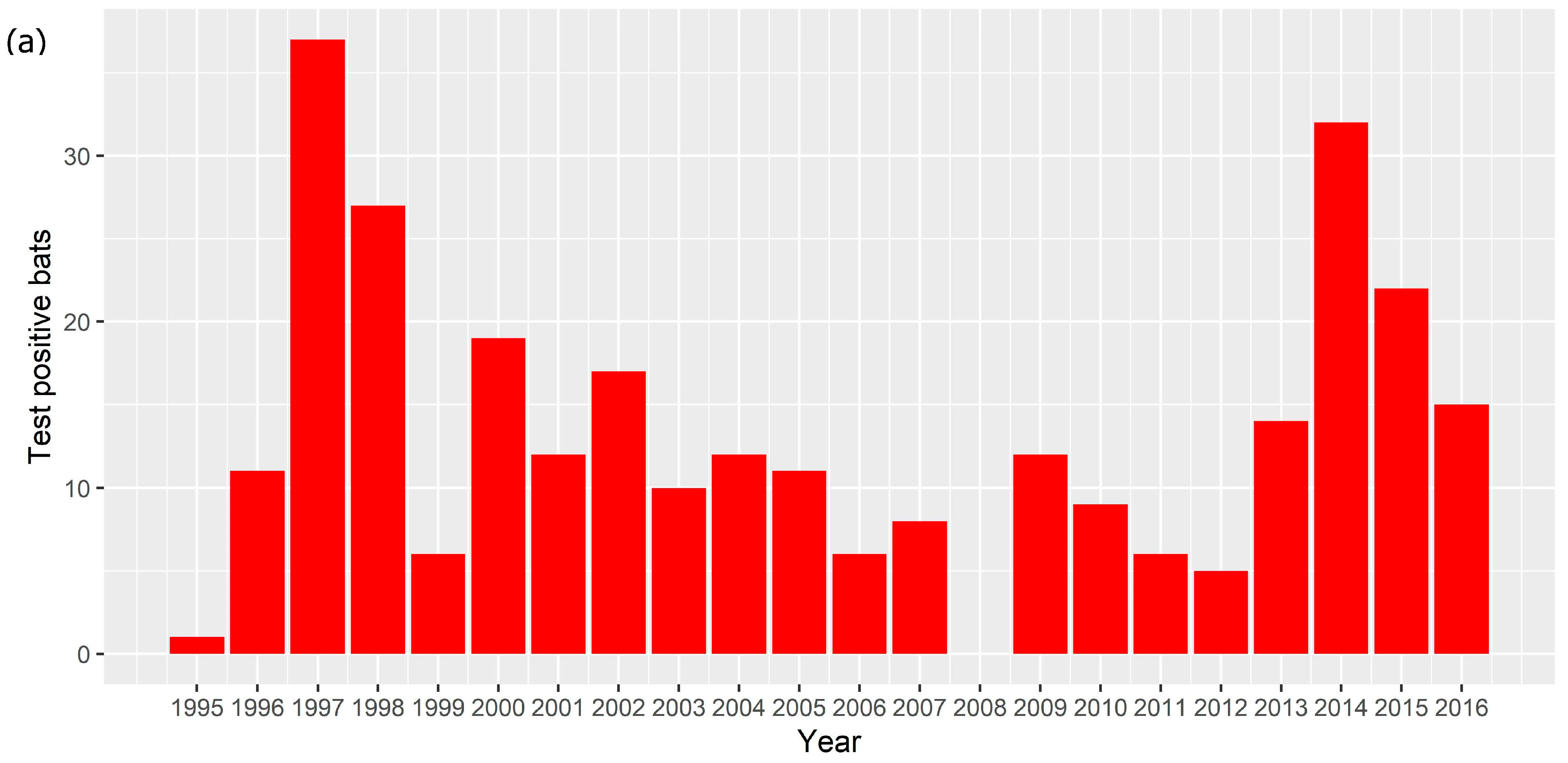
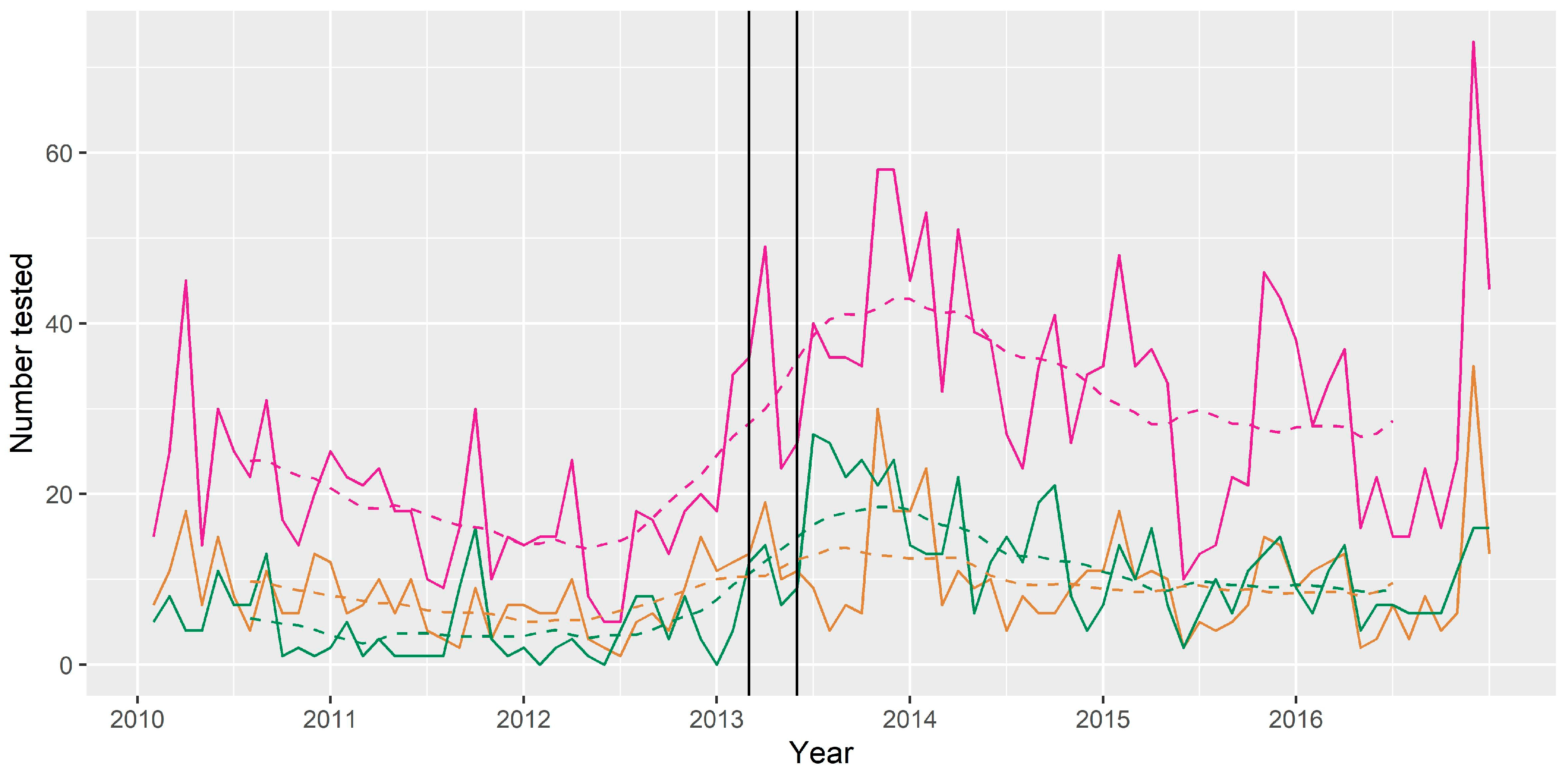
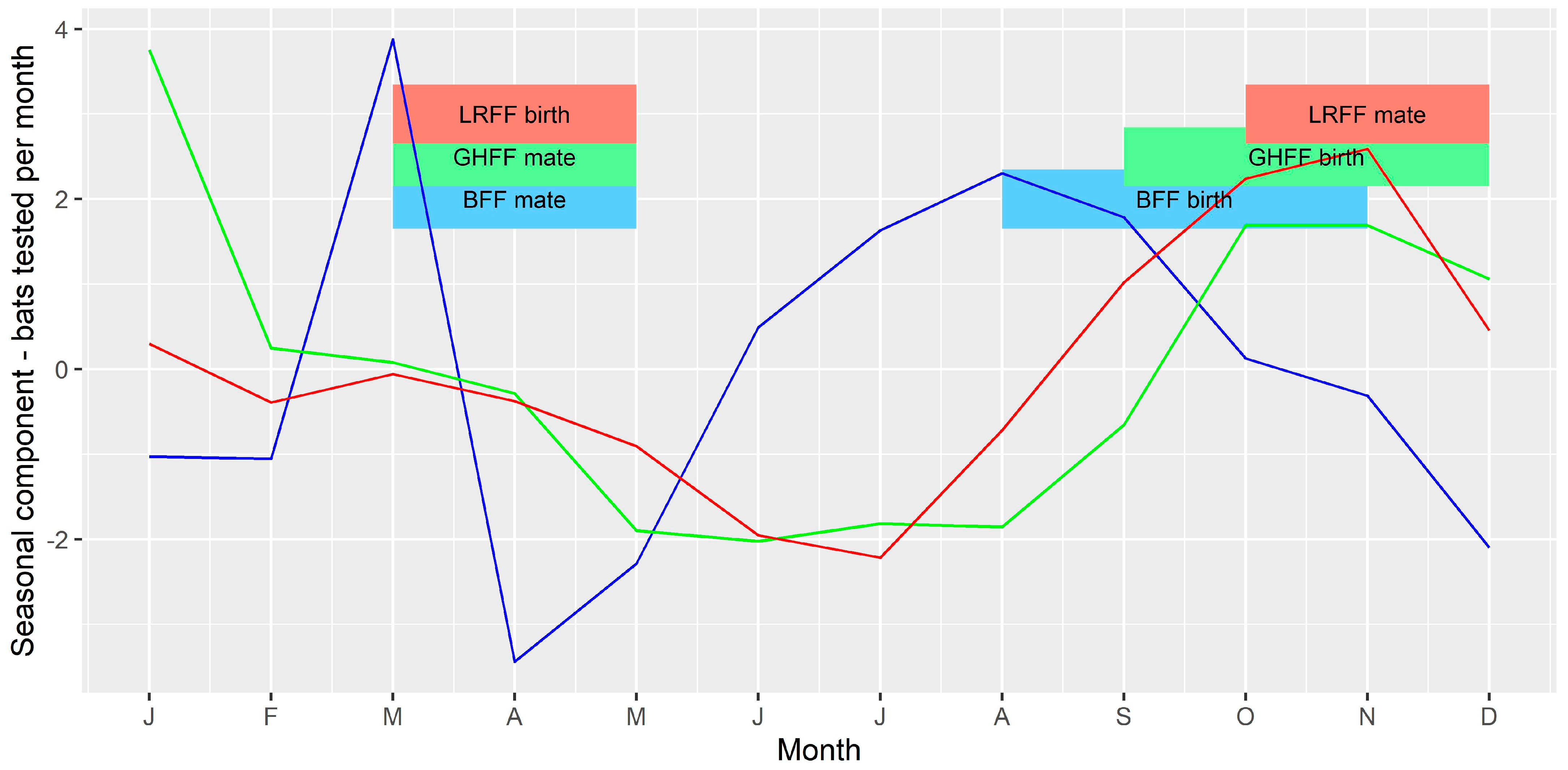
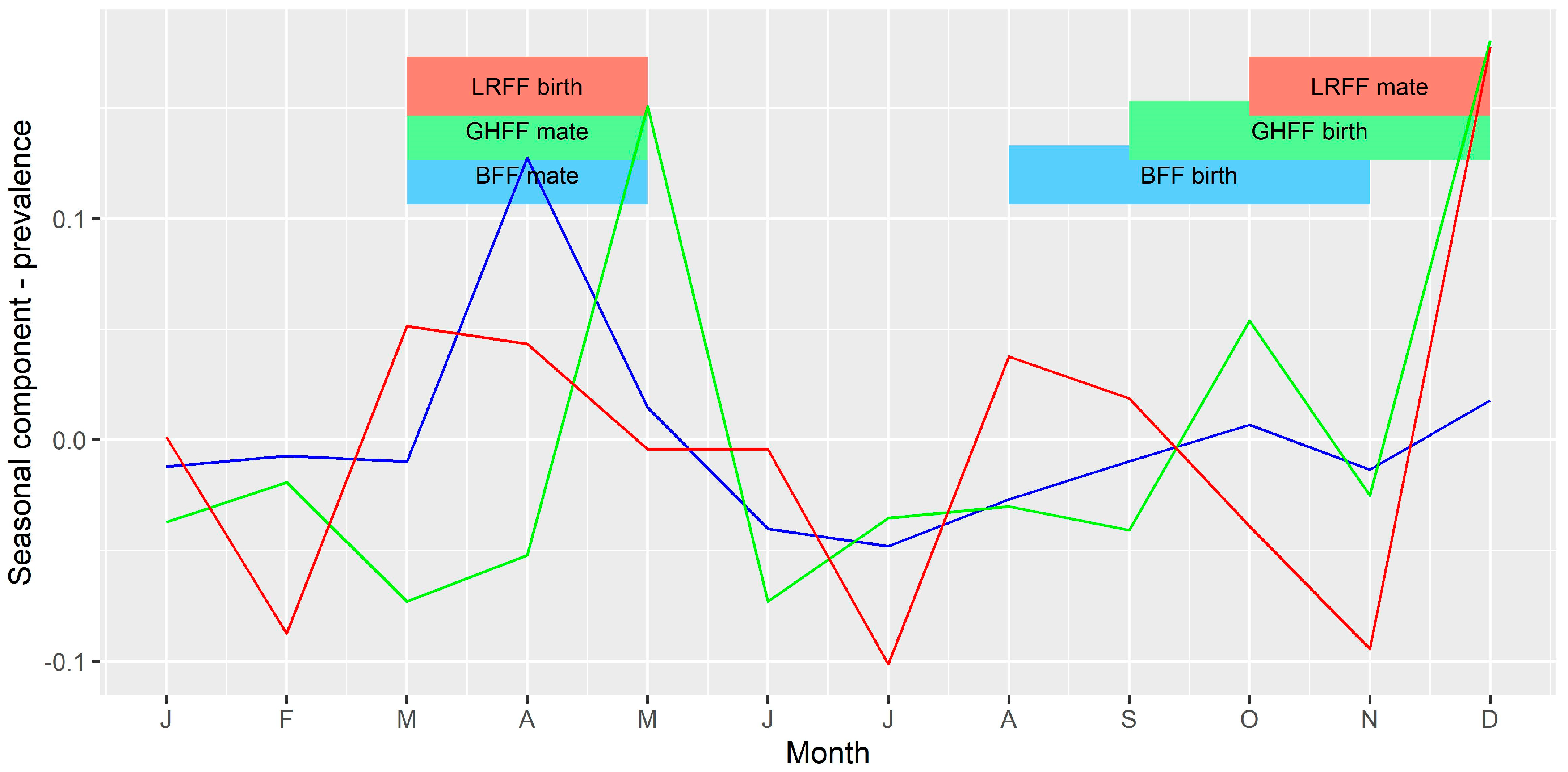
3.4. Time Series Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Afonso, C.L.; Amarasinghe, G.K.; Bányai, K.; Bào, Y.; Basler, C.F.; Bavari, S.; Bejerman, N.; Blasdell, K.R.; Briand, F.-X.; Briese, T.; et al. Taxonomy of the order Mononegavirales: Update 2016. Arch. Virol. 2016, 161, 2351–2360. [Google Scholar] [CrossRef]
- Gould, A.R.; Hyatt, A.D.; Lunt, R.; Kattenbelt, J.A.; Hengstberger, S.; Blacksell, S. Characterisation of a novel lyssavirus isolated from Pteropid bats in Australia. Virus Res. 1998, 54, 165–187. [Google Scholar] [CrossRef]
- Fraser, G.C.; Hooper, P.T.; Lunt, R.A.; Gould, A.R.; Gleeson, L.J.; Hyatt, A.D.; Russell, G.M.; Kattenbelt, J.A. Encephalitis Caused by a Lyssavirus in Fruit Bats in Australia. Emerg. Infect. Dis. 1996, 2, 327–331. [Google Scholar] [CrossRef]
- Skerratt, L.F.; Speare, R.; Berger, L.; Winsor, H. Lyssaviral Infection and Lead Poisoning in Black Flying Foxes from Queensland. J. Wildl. Dis. 1998, 34, 355–361. [Google Scholar] [CrossRef]
- Gould, A.R.; Kattenbelt, J.A.; Gumley, S.G.; Lunt, R.A. Characterisation of an Australian bat lyssavirus variant isolated from an insectivorous bat. Virus Res. 2002, 89, 1–28. [Google Scholar] [CrossRef]
- Field, H.E. Evidence of Australian bat lyssavirus infection in diverse Australian bat taxa. Zoonoses Public Health 2018, 65, 742–748. [Google Scholar] [CrossRef]
- Prada, D.; Boyd, V.; Baker, M.L.; Jackson, B.; O’Dea, M. Insights into Australian Bat Lyssavirus in Insectivorous Bats of Western Australia. Trop. Med. Infect. Dis. 2019, 4, 46. [Google Scholar] [CrossRef]
- Barrett, J. Australian Bat Lyssavirus; School of Veterinary Science, The University of Queensland: Brisbane, Australia, 2004. [Google Scholar]
- Allworth, A.; Murray, K.; Morgan, J. A human case of encephalitis due to a lyssavirus recently identified in fruit bats. Commun. Dis. Intell. 1996, 20, 504. [Google Scholar]
- Francis, J.R.; Nourse, C.; Vaska, V.L.; Calvert, S.; Northill, J.A.; McCall, B.; Mattke, A.C. Australian Bat Lyssavirus in a Child: The First Reported Case. Pediatrics 2014, 133, e1063–e1067. [Google Scholar] [CrossRef]
- Hanna, J.N.; Carney, I.K.; Deverill, J.E.; Botha, J.A.; Smith, G.A.; Serafin, I.L.; Harrower, B.J.; Tannenberg, A.E.G.; Fitzpatrick, P.F.; Searle, J.W. Australian bat lyssavirus infection: A second human case, with a long incubation period. Med. J. Aust. 2000, 172, 597–599. [Google Scholar] [CrossRef]
- Samaratunga, H.; Searle, J.W.; Hudson, N. Non-rabies Lyssavirus human encephalitis from fruit bats: Australian bat Lyssavirus (pteropid Lyssavirus) infection. Neuropathol. Appl. Neurobiol. 1998, 24, 331–335. [Google Scholar] [CrossRef]
- Annand, E.J.; Reid, P.A. Clinical review of two fatal equine cases of infection with the insectivorous bat strain of Australian bat lyssavirus. Aust. Veter J. 2014, 92, 324–332. [Google Scholar] [CrossRef]
- Shinwari, M.W.; Annand, E.J.; Driver, L.; Warrilow, D.; Harrower, B.; Allcock, R.J.; Pukallus, D.; Harper, J.; Bingham, J.; Kung, N.; et al. Australian bat lyssavirus infection in two horses. Veter Microbiol. 2014, 173, 224–231. [Google Scholar] [CrossRef]
- Wright, T. A Dog Tests Antibody Positive for Lyssavirus. In NSW Animal Health Surveillance 2013; NSW Department of Primary Industries: New South Wales, Australia, 2013; pp. 3–4. [Google Scholar]
- Woods, R.; Reiss, A.; Cox-Witton, K.; Grillo, V.L.; Peters, A. The Importance of Wildlife Disease Monitoring as Part of Global Surveillance for Zoonotic Diseases: The Role of Australia. Trop. Med. Infect. Dis. 2019, 4, 29. [Google Scholar] [CrossRef]
- Kardamanidis, K.; Cashman, P.; Durrheim, D. Travel and non-travel associated rabies post exposure treatment in New South Wales residents, Australia, 2007–2011: A cross-sectional analysis. Travel Med. Infect. Dis. 2013, 11, 421–426. [Google Scholar] [CrossRef]
- Paterson, B.J.; Butler, M.; Eastwood, K.; Cashman, P.; Jones, A.; Durrheim, D.N. Cross sectional survey of human-bat interaction in Australia: Public health implications. BMC Public Health 2014, 14, 58. [Google Scholar] [CrossRef]
- Quinn, E.; Massey, P.D.; Cox-Witton, K.; Paterson, B.J.; Eastwood, K.; Durrheim, D. Understanding human—bat interactions in NSW, Australia: Improving risk communication for prevention of Australian bat lyssavirus. BMC Veter Res. 2014, 10, 144. [Google Scholar] [CrossRef]
- Sánchez, C.A.; Baker, M.L. Disease Risk Perception and Safety Practices: A Survey of Australian Flying Fox Rehabilitators. PLoS Negl. Trop. Dis. 2016, 10, e0004411. [Google Scholar] [CrossRef]
- Young, M.K.; McCall, B.J. Potential exposure to Australian bat lyssavirus in south east Queensland: What has changed in 12 years? Commun. Dis. Intell. Q. Rep. 2010, 34, 334. [Google Scholar]
- Holz, P.H.; Lumsden, L.F.; Druce, J.; Legione, A.R.; Vaz, P.; Devlin, J.M.; Hufschmid, J. Virus survey in populations of two subspecies of bent-winged bats (Miniopterus orianae bassanii and oceanensis) in south-eastern Australia reveals a high prevalence of diverse herpesviruses. PLoS ONE 2018, 13, e0197625. [Google Scholar] [CrossRef]
- Bunn, C.; Garner, G. Update on surveillance for Australian bat Lyssavirus. Australas. Epidemiol. 1997, 4, 27. [Google Scholar]
- Si, D.; Marquess, J.; Donnan, E.; Harrower, B.; McCall, B.; Bennett, S.; Lambert, S. Potential Exposures to Australian Bat Lyssavirus Notified in Queensland, Australia, 2009−2014. PLoS Negl. Trop. Dis. 2016, 10, e0005227. [Google Scholar] [CrossRef]
- Warrilow, D.; Harrower, B.; Smith, I.L.; Field, H.; Taylor, R.; Walker, G.C.; Smith, G.A. Public health surveillance for Australian bat lyssavirus in Queensland, Australia, 2000–2001. Emerg. Infect. Dis. 2003, 9, 262. [Google Scholar] [CrossRef]
- Animal Health Australia. Disease strategy: Australian bat lyssavirus (Version 3.0). In Australian Veterinary Emergency Plan (Ausvetplan), 3rd ed.; Primary Industries Ministerial Council: Canberra, Australia, 2009. [Google Scholar]
- Wildlife Health Australia. Australian Bat Lyssavirus—Fact Sheet; Wildlife Health Australia: Mosman, Australia, 2019. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Zeileis, A.; Hothorn, T. Diagnostic Checking in Regression Relationships. 2002. Available online: https://cran.r-project.org/web/packages/lmtest/vignettes/lmtest-intro.pdf (accessed on 25 September 2019).
- Lele, S.; Keim, J.; Solymos, P. Resource selection (probability) functions for use-availability data. Package ‘ResourceSelection’, Version 0.3-2. 2017. Available online: https://rdrr.io/cran/ResourceSelection/man/rsf.html (accessed on 25 September 2019).
- Hall, L.S.; Richards, G. Flying Foxes: Fruit and Blossom Bats of Australia; UNSW Press: Randwick, Australia, 2000. [Google Scholar]
- Heard, D. Chiroptera (bats). In Zoo and Wild Animal Medicine, 5th ed.; Fowler, M.E., Miller, R.E., Eds.; WB Saunders Co.: Philadelphia, PA, USA, 2003; pp. 315–333. [Google Scholar]
- Field, H. The Ecology of Hendra Virus and Australian Bat Lyssavirus; School of Veterinary Science, The University of Queensland: Brisbane, Australia, 2005. [Google Scholar]
- George, D.B.; Webb, C.T.; Farnsworth, M.L.; O’Shea, T.J.; Bowen, R.A.; Smith, D.L.; Stanley, T.R.; Ellison, L.E.; Rupprecht, C.E. Host and viral ecology determine bat rabies seasonality and maintenance. Proc. Natl. Acad. Sci. USA 2011, 108, 10208–10213. [Google Scholar] [CrossRef]
- Bonwitt, J.; Oltean, H.; Lang, M.; Kelly, R.M.; Goldoft, M. Bat rabies in Washington State: Temporal-spatial trends and risk factors for zoonotic transmission (2000–2017). PLoS ONE 2018, 13, e0205069. [Google Scholar] [CrossRef]
- Picard-Meyer, E.; Robardet, E.; Arthur, L.; Larcher, G.; Harbusch, C.; Servat, A.; Cliquet, F. Bat Rabies in France: A 24-Year Retrospective Epidemiological Study. PLoS ONE 2014, 9, e98622. [Google Scholar] [CrossRef]
- Qld Department of Environment and Science. Netting Fruit Trees. 2020. Available online: https://environment.des.qld.gov.au/wildlife/animals/living-with/bats/flying-foxes/netting-fruit-trees (accessed on 8 June 2020).
- WFF. Wildlife Friendly Fencing Project. Available online: https://www.wildlifefriendlyfencing.com/WFF/Home.html (accessed on 8 June 2020).
- Barrett, J.; Höger, A.; Agnihotri, K.; Oakey, J.; Skerratt, L.F.; Field, H.E.; Meers, J.; Smith, C. An unprecedented cluster of Australian bat lyssavirus in Pteropus conspicillatus indicates pre-flight flying fox pups are at risk of mass infection. Zoonoses Public Health 2020, 67, 435–442. [Google Scholar] [CrossRef]
- NSW Department of Primary Industries. CVO Bulletin to Wildlife Carers: Australian Bat Lyssavirus Infection in Juvenile Bats. 2016. Available online: https://www.vpb.nsw.gov.au/sites/default/files/images/NEWS_20151218_DPI_CVO%20Bulletin%20bat%20lyssavirus%20infection%20in%20juvenile%20bats.pdf (accessed on 8 June 2020).
- Hayman, D.T.S.; Luis, A.D.; Restif, O.; Baker, K.S.; Fooks, A.R.; Leach, C.B.; Horton, D.L.; Suu-Ire, R.; Cunningham, A.A.; Wood, J.L.N.; et al. Maternal antibody and the maintenance of a lyssavirus in populations of seasonally breeding African bats. PLoS ONE 2018, 13, e0198563. [Google Scholar] [CrossRef]
- Wise, E.L.; Marston, D.A.; Banyard, A.C.; Goharriz, H.; Selden, D.; MacLaren, N.; Goddard, T.; Johnson, N.; McElhinney, L.M.; Brouwer, A.; et al. Passive surveillance of United Kingdom bats for lyssaviruses (2005–2015). Epidemiol. Infect. 2017, 145, 2445–2457. [Google Scholar] [CrossRef]
- May, F.; Mann, K.D.; Francis, D.; Young, M.K. Identification of focus areas for Australian Bat Lyssavirus potential exposure prevention in the Metro North Hospital and Health Service region. Zoonoses Public Health 2020. [Google Scholar] [CrossRef]
- O’Dea, M.A.; Tu, S.-L.; Pang, S.; De Ridder, T.; Jackson, B.; Upton, C. Genomic characterization of a novel poxvirus from a flying fox: Evidence for a new genus? J. Gen. Virol. 2016, 97, 2363–2375. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Name | Encodes | Source |
|---|---|---|
| Sex | Sex of the tested bat (Male/Female/Unspecified) | Extracted from free text field if recorded. |
| Age | Age class of the tested bat (Adult/Juvenile/Unspecified) | Extracted from free text field if recorded. |
| Reproductive status | Reproductive status of the tested bat (Pregnant/Lactating/Unspecified) | Extracted from free text field if recorded. |
| Season | Austral seasons, aligned to temperate parts of Australia Summer (December to February) Autumn (March to May) Winter (June to August) Spring (September to November) | Determined from event start date recorded in database |
| State | State or territory where the bat was found | Location information recorded in database |
| Year | Year in which the bat was found | Extracted from event start date recorded in database |
| Month | Calendar month in which the bat was found | Extracted from event start date recorded in database (only used for time series analysis). |
| Species | Bat species | Taxonomic information recorded in database. |
| Neurological signs | Bat was recorded as having neurological signs, or neurological signs were described in the case history supplied and interpreted as such by the authors (Yes/Unspecified) | Extracted from free text field if recorded. |
| Traumatic injury | Bat was recorded as having traumatic injury, or injuries were described in the case history supplied (Yes/Unspecified) | Extracted from free text field if recorded. |
| Reason for testing | Reason why the bat was submitted for ABLV testing. Hierarchy of potential reasons, with up to three of these recorded for an event: (1) human contact (2) pet contact (3) neurological signs (4) other signs Human contact is assigned as the first submission reason even if the contact was only determined later. | Inferred from free text field |
| Circumstances | The circumstances in which the bat came to human attention (see Table 2). Designed to capture what is reported by the non-technical observer that finds or rescues the bat, so there is no interpretation of observations into clinical signs. | Extracted from free text field if recorded. |
| Dog contact | Bat was recorded as having contact or possible contact with a dog (Yes/Unspecified) | Extracted from free text field if recorded. |
| Human contact | Bat had reported contact with a human (Yes/No) * | Extracted from free text field if recorded. |
| Infection status | Outcome of testing for ABLV | Testing information recorded in database. |
| Category | Definition |
|---|---|
| Abnormal behaviour | History included screaming/vocalising, self-trauma, attacking other animals or feeding in daylight hours. |
| Entanglement | Most commonly tangled in fruit netting or fences, occasionally other entanglements such as fishing line or tinsel. |
| Grounded | Found in an abnormal location unable to evade capture. History often includes the phrase ‘found on the ground’ or ‘unable to fly’ or both. |
| Found dead single | Single bat found dead without any known pet contact |
| Mass mortality/morbidity event | Multiple animals found dead or unwell in proximity at the same time or over a limited time period, including heat stress events |
| Orphaned | Described as orphaned or found with a deceased mother; young bats in care, assumed to be orphaned; or foetus tested as well as mother |
| Other | Various forms of misadventure such as electrocution or hit by car; other miscellaneous presentations such as part of a research project, or translocation. |
| Pet contact | Any actual or possible contact between a bat and pet animal—usually dog or cat—regardless whether initiated by pet or bat and regardless of whether bat was alive or dead at the time of contact |
| Unspecified | No information on circumstances of find recorded. |
| Variables | Categories | Multivariable Model | Univariable Model | ||
|---|---|---|---|---|---|
| OR (95%CI) | p | OR (95%CI) | p | ||
| Intercept | 0.02 (0.01–0.05) | <0.01 | |||
| Season | Autumn | 2.30 (1.12–5.09) | 0.03 | 2.64 (1.30–5.80) | 0.01 |
| Spring | 1.57 (0.75–3.50) | 0.25 | 1.93 (0.94–4.27) | 0.08 | |
| Summer | 2.29 (1.13–5.05) | 0.03 | 2.71 (1.35–5.91) | 0.01 | |
| Winter | 1 | 1 | |||
| Species | Black flying-fox | 1 | 1 | ||
| Grey-headed flying-fox | 1.23 (0.72–2.09) | 0.45 | 1.30 (0.76–2.19) | 0.34 | |
| Little red flying-fox | 3.01 (1.72–5.23) | <0.01 | 3.28 (1.89–5.64) | <0.01 | |
| Spectacled flying-fox | 4.82 (1.06–16.00) | 0.02 | 5.36 (1.19–17.62) | 0.01 | |
| Unidentified flying- fox | 0.78 (0.26–1.88) | 0.62 | 0.83 (0.28–1.98) | 0.70 | |
| Variables | Multivariable Model | Univariable Models | ||
|---|---|---|---|---|
| OR (95%CI) | p | OR (95%CI) | p | |
| Intercept | 0.03 (0.02–0.05) | <0.01 | ||
| Neurological signs | 13.30 (7.94–22.78) | <0.01 | 16.67 (10.83–26.04) | <0.01 |
| Traumatic injury | 0.24 (0.10–0.51) | <0.01 | 0.29 (0.12–0.60) | <0.01 |
| Human contact | 2.03 (1.22–3.40) | 0.01 | 1.49 (0.99–2.23) | 0.06 |
| Dog contact | 0.33 (0.14–0.74) | 0.01 | 0.12 (0.05–0.23) | <0.01 |
| Variables | Categories | OR (95%CI) | p |
|---|---|---|---|
| Intercept | 0.08 (0.05–0.11) | <0.01 | |
| Circumstances | Abnormal behaviour | 5.44 (2.21–12.57) | <0.01 |
| Entanglement | 0.23 (0.07–0.59) | <0.01 | |
| Grounded | 3.53 (2.14–5.89) | <0.01 | |
| Found dead single | 0.41 (0.07–1.40) | 0.23 | |
| Mass mortality/morbidity event | 0.33 (0.02–1.58) | 0.28 | |
| Orphaned | 1.76 (0.58–4.45) | 0.27 | |
| Other | 0.98 (0.28–2.58) | 0.96 | |
| Pet contact | 0.12 (0.04–0.26) | <0.01 | |
| Unspecified | 1 |
| Variables | Categories | Multivariable Model | Univariable Models | ||
|---|---|---|---|---|---|
| OR (95%CI) | p | OR (95%CI) | p | ||
| Intercept | 0.04 (0.02–0.06) | <0.01 | |||
| Circumstances | Abnormal behaviour | 17.02 (7.28–39.07) | <0.01 | 16.33 (7.02–37.33) | <0.01 |
| Entanglement | 45.10 (28.23–74.56) | <0.01 | 43.86 (27.65–72.00) | <0.01 | |
| Grounded | 7.05 (4.14–12.17) | <0.01 | 6.60 (3.90–11.34) | <0.01 | |
| Found dead single | 1.60 (0.46–4.28) | 0.40 | 1.63 (0.47–4.37) | 0.38 | |
| Mass mortality/morbidity event | 1.31 (0.21–4.64) | 0.72 | 1.29 (0.20–4.54) | 0.74 | |
| Orphaned | 12.64 (5.87–26.82) | <0.01 | 14.13 (6.62–29.76) | <0.01 | |
| Other | 15.48 (7.88–30.42) | <0.01 | 14.70 (7.51–28.76) | <0.01 | |
| Unspecified | 68.44 (44.37–109.70) | <0.01 | 64.92 (42.36–103.41) | <0.01 | |
| Pet contact | 1 | 1 | |||
| Year | Winter | 1 | 1 | ||
| Autumn | 0.81 (0.54–1.23) | 0.32 | 1.53 (1.12–2.10) | 0.01 | |
| Spring | 1.41 (0.95–2.10) | 0.09 | 1.91 (1.42–2.57) | <0.01 | |
| Summer | 0.94 (0.63–1.41) | 0.76 | 1.82 (1.34–2.47) | <0.01 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iglesias, R.; Cox-Witton, K.; Field, H.; Skerratt, L.F.; Barrett, J. Australian Bat Lyssavirus: Analysis of National Bat Surveillance Data from 2010 to 2016. Viruses 2021, 13, 189. https://doi.org/10.3390/v13020189
Iglesias R, Cox-Witton K, Field H, Skerratt LF, Barrett J. Australian Bat Lyssavirus: Analysis of National Bat Surveillance Data from 2010 to 2016. Viruses. 2021; 13(2):189. https://doi.org/10.3390/v13020189
Chicago/Turabian StyleIglesias, Rachel, Keren Cox-Witton, Hume Field, Lee F. Skerratt, and Janine Barrett. 2021. "Australian Bat Lyssavirus: Analysis of National Bat Surveillance Data from 2010 to 2016" Viruses 13, no. 2: 189. https://doi.org/10.3390/v13020189
APA StyleIglesias, R., Cox-Witton, K., Field, H., Skerratt, L. F., & Barrett, J. (2021). Australian Bat Lyssavirus: Analysis of National Bat Surveillance Data from 2010 to 2016. Viruses, 13(2), 189. https://doi.org/10.3390/v13020189