
Inactivation of Polyomavirus SV40 as Surrogate for Human Papillomaviruses by Chemical Disinfectants

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Disinfectants, Conditions, and Parameters Used in This Study
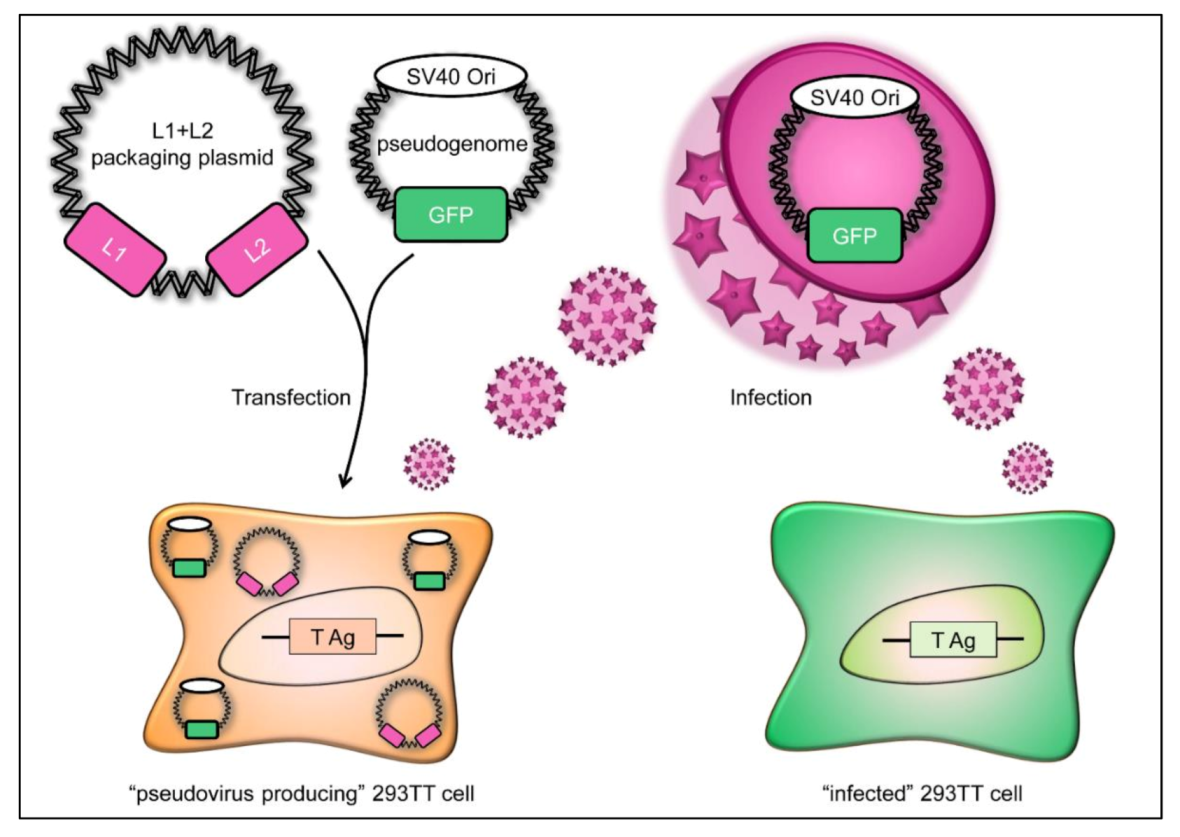
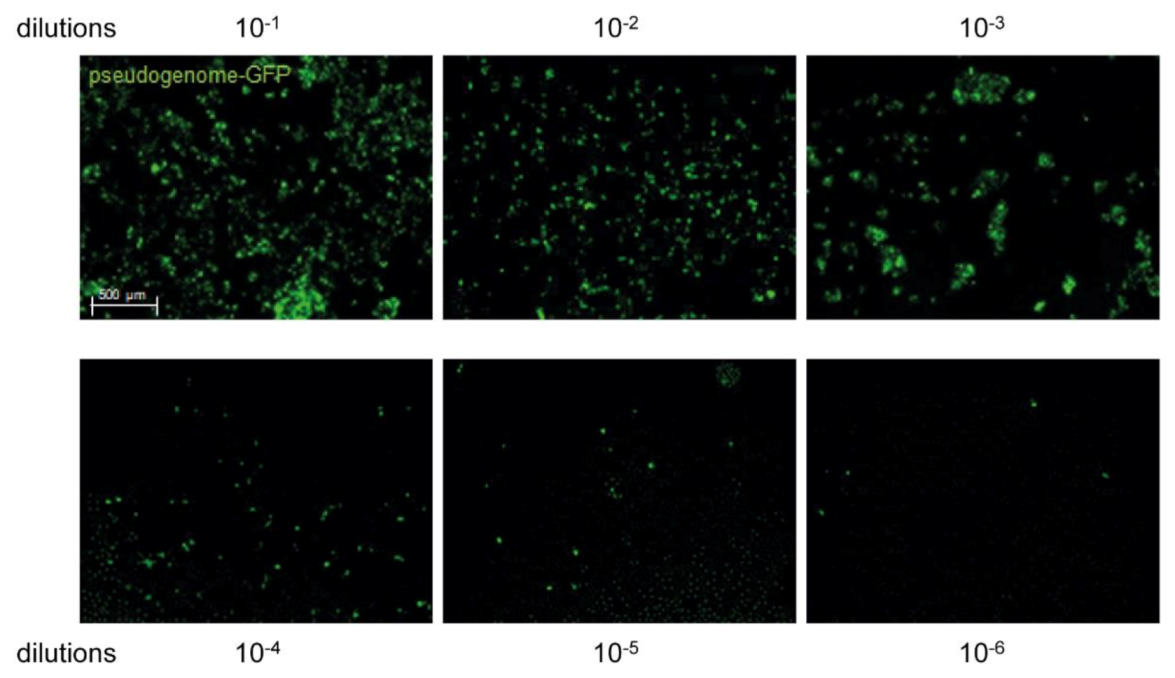
2.2. HPV16 Pseudovirus Production and Titration
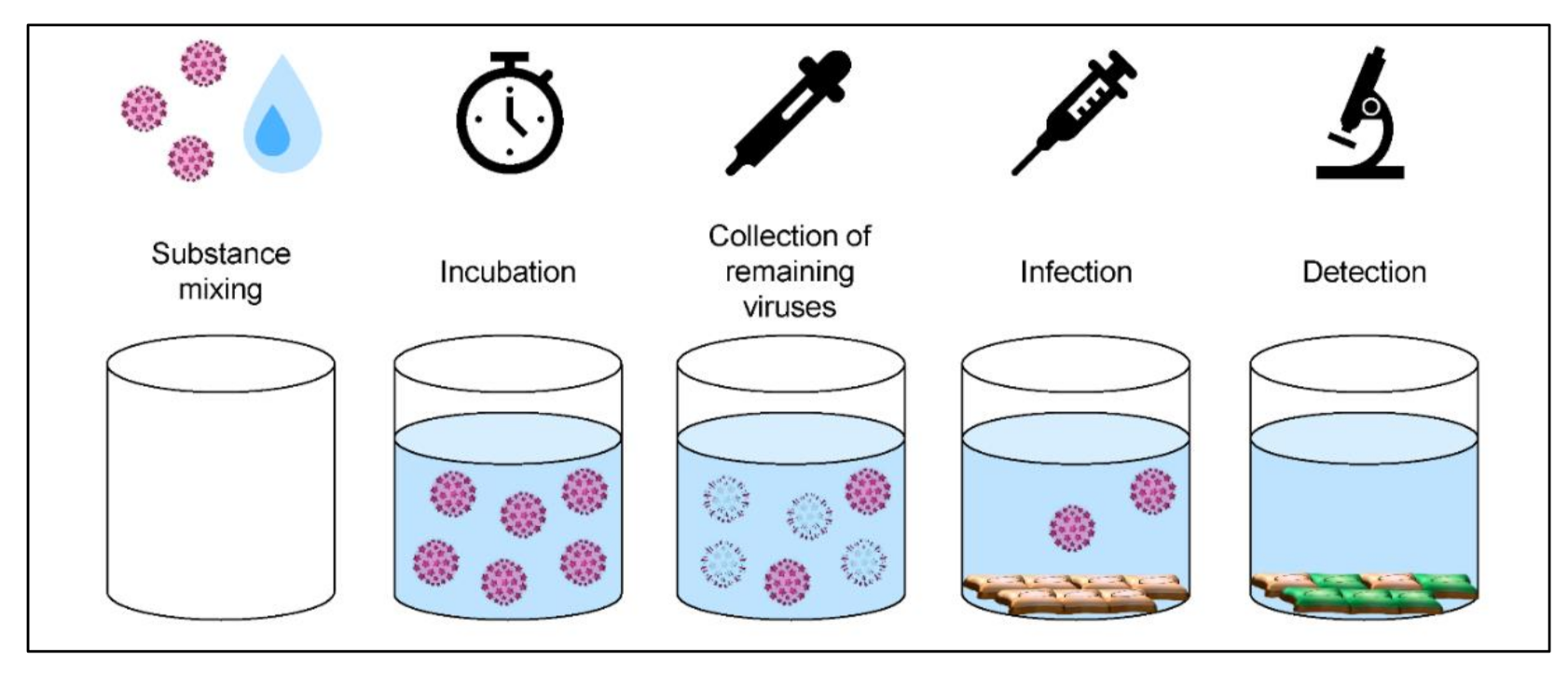
2.3. Quantitative Suspension Test Using HPV16 Pseudovirus
2.4. Production of SV40 Test Virus Suspension
2.5. Quantitative Suspension Tests with SV40
2.6. Statistical Analysis
3. Results
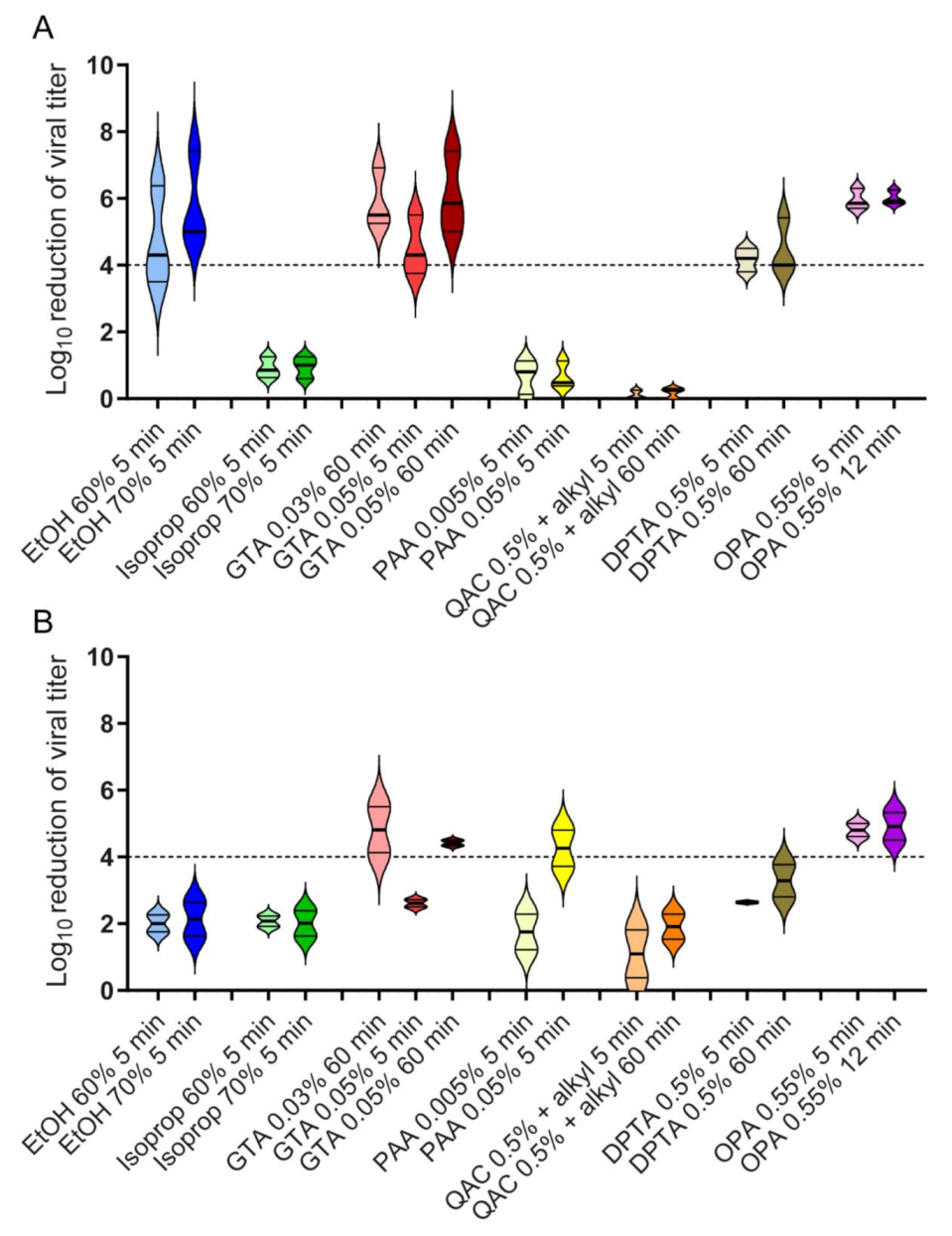
3.1. Efficacy of Chemical Disinfectants against HPV16 Pseudovirus
3.2. Efficacy of Chemical Disinfectants against Polyomavirus SV40
3.3. Comparison of the Effectiveness of Chemical Disinfectants against HPV16 Pseudovirus and SV40
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schiffman, M.; Castle, P.E.; Jeronimo, J.; Rodriguez, A.C.; Wacholder, S. Human papillomavirus and cervical cancer. Lancet 2007, 370, 890–907. [Google Scholar] [CrossRef]
- Bouvard, V.; Baan, R.; Straif, K.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A review of human carcinogens—Part B: Biological agents. Lancet Oncol. 2009, 10, 321–322. [Google Scholar] [CrossRef]
- de Martel, C.; Plummer, M.; Vignat, J.; Franceschi, S. Worldwide burden of cancer attributable to HPV by site, country and HPV type. Int. J. Cancer 2017, 141, 664–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, S. Infectious risk of endovaginal and transrectal ultrasonography: Systematic review and meta-analysis. J. Hosp. Infect. 2013, 83, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.T.; Yeung, A.C.; Chan, P.K.; Graham, C.A. Transvaginal ultrasound probe contamination by the human papillomavirus in the emergency department. Emerg. Med. J. 2013, 30, 472–475. [Google Scholar] [CrossRef]
- M’Zali, F.; Bounizra, C.; Leroy, S.; Mekki, Y.; Quentin-Noury, C.; Kann, M. Persistence of microbial contamination on transvaginal ultrasound probes despite low-level disinfection procedure. PLoS ONE 2014, 9, e93368. [Google Scholar] [CrossRef]
- Ryndock, E.J.; Meyers, C. A risk for non-sexual transmission of human papillomavirus? Expert Rev. Anti-Infect. Ther. 2014, 12, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Moshkanbaryans, L.; Meyers, C.; Ngu, A.; Burdach, J. The importance of infection prevention and control in medical ultrasound. Australas. J. Ultrasound Med. 2015, 18, 96–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casalegno, J.S.; Le Bail Carval, K.; Eibach, D.; Valdeyron, M.L.; Lamblin, G.; Jacquemoud, H.; Mellier, G.; Lina, B.; Gaucherand, P.; Mathevet, P.; et al. High risk HPV contamination of endocavity vaginal ultrasound probes: An underestimated route of nosocomial infection? PLoS ONE 2012, 7, e48137. [Google Scholar] [CrossRef] [Green Version]
- Gallay, C.; Miranda, E.; Schaefer, S.; Catarino, R.; Jacot-Guillarmod, M.; Menoud, P.A.; Guerry, F.; Achtari, C.; Sahli, R.; Vassilakos, P.; et al. Human papillomavirus (HPV) contamination of gynaecological equipment. Sex. Transm. Infect. 2016, 92, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Buck, C.B.; Pastrana, D.V.; Lowy, D.R.; Schiller, J.T. Efficient intracellular assembly of papillomaviral vectors. J. Virol. 2004, 78, 751–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, C.B.; Pastrana, D.V.; Lowy, D.R.; Schiller, J.T. Generation of HPV pseudovirions using transfection and their use in neutralization assays. Methods Mol. Med. 2005, 119, 445–462. [Google Scholar] [PubMed]
- Buck, C.B.; Thompson, C.D. Production of papillomavirus-based gene transfer vectors. Curr. Protoc. Cell Biol. 2007. [CrossRef]
- Biryukov, J.; Meyers, C. Papillomavirus Infectious Pathways: A Comparison of Systems. Viruses 2015, 7, 4303–4325. [Google Scholar] [CrossRef] [Green Version]
- DiGiuseppe, S.; Luszczek, W.; Keiffer, T.R.; Bienkowska-Haba, M.; Guion, L.G.; Sapp, M.J. Incoming human papillomavirus type 16 genome resides in a vesicular compartment throughout mitosis. Proc. Natl. Acad. Sci. USA 2016, 113, 6289–6294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moens, U. Human Polyomaviruses and Papillomaviruses. Int. J. Mol. Sci. 2018, 19, 2360. [Google Scholar] [CrossRef] [Green Version]
- Robert, K.-I.; Deutschen Gesellschaft zur Bekämpfung der Viruskrankheiten; Disinfektionsmittelkommission der Deutschen Gesellschaft für Hygiene und Mikrobiologie. [Evaluation and declaration of effectiveness of disinfectants against viruses. Position of the Virucide Study Group of the Robert Koch Institute (RKI) and the “Virus Disinfection” Professional Committee of the German Society for Control of “Virus Infections” and the Disinfectant Committee of the German Society of Public Health and Microbiology]. Bundesgesundheitsblatt Gesundh. Gesundh. 2004, 47, 62–66. [Google Scholar]
- Buck, C.B.; Thompson, C.D.; Roberts, J.N.; Muller, M.; Lowy, D.R.; Schiller, J.T. Carrageenan is a potent inhibitor of papillomavirus infection. PLoS Pathog. 2006, 2, e69. [Google Scholar] [CrossRef] [Green Version]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Arch. Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Spearman, C. The Method of “Right and Wrong Cases” (Constant Stimuli) without Gauss’s Formula. Br. J. Psychol. 1908, 2, 227–242. [Google Scholar]
- Rabenau, H.F.; Schwebke, I.; Blumel, J.; Eggers, M.; Glebe, D.; Rapp, I.; Sauerbrei, A.; Steinmann, E.; Steinmann, J.; Willkommen, H.; et al. [Guideline of the German Association for the Control of Viral Diseases (DVV) eV and the Robert Koch Institute (RKI) for testing chemical disinfectants for effectiveness against viruses in human medicine. Version of 1 December, 2014]. Bundesgesundheitsblatt Gesundh. Gesundh. 2015, 58, 493–504. [Google Scholar] [CrossRef] [Green Version]
- EN 14476:2013/A1:2015. Chemical Disinfectants and Antiseptics. Virucidal Quantitative Suspension Test for Chemical Disinfectants and Antiseptics Used in Human Medicine. Test Method and Requirements (Phase 2, Step 1). 2015. Available online: https://shop.bsigroup.com/products/chemical-disinfectants-and-antiseptics-quantitative-suspension-test-for-the-evaluation-of-virucidal-activity-in-the-medical-area-test-method-and-requirements-phase-2-step-1?pid=000000000030401479 (accessed on 5 June 2021).
- Eggers, H.J. Experiments on antiviral activity of hand disinfectants. Some theoretical and practical considerations. Zentralbl. Bakteriol. 1990, 273, 36–51. [Google Scholar] [CrossRef]
- Sauerbrei, A.; Wutzler, P. Virucidal efficacy of povidone-iodine-containing disinfectants. Lett. Appl. Microbiol. 2010, 51, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.; Henningsen, L.; Paulmann, D.; Bischoff, B.; Todt, D.; Steinmann, E.; Steinmann, J.; Brill, F.H.H.; Steinmann, J. Evaluation of the virucidal efficacy of disinfectant wipes with a test method simulating practical conditions. Antimicrob. Resist. Infect. Control. 2019, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Meyers, J.; Ryndock, E.; Conway, M.J.; Meyers, C.; Robison, R. Susceptibility of high-risk human papillomavirus type 16 to clinical disinfectants. J. Antimicrob. Chemother. 2014, 69, 1546–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egawa, N.; Shiraz, A.; Crawford, R.; Saunders-Wood, T.; Yarwood, J.; Rogers, M.; Sharma, A.; Eichenbaum, G.; Doorbar, J. Dynamics of papillomavirus in vivo disease formation & susceptibility to high-level disinfection-Implications for transmission in clinical settings. EBioMedicine 2021, 63, 103177. [Google Scholar] [PubMed]
- Ryndock, E.; Robison, R.; Meyers, C. Susceptibility of HPV16 and 18 to high level disinfectants indicated for semi-critical ultrasound probes. J. Med. Virol. 2016, 88, 1076–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabenau, H.F.; Steinmann, J.; Rapp, I.; Schwebke, I.; Eggers, M. Evaluation of a virucidal quantitative carrier test for surface disinfectants. PLoS ONE 2014, 9, e86128. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Biocidal Substance | Company |
|---|---|
| Ethanol (EtOH), ≥99.5% | Roth (Karlsruhe, Germany) |
| Isopropanol (Isoprop.), ≥99.8% | Roth (Karlsruhe, Germany) |
| Glutaraldehyde (GTA) | Dow (Midland, MI, USA) |
| Peracetic Acid (PAA) | Stockmeier (Bielefeld, Germany) |
| Quaternary Ammonium Compounds (QAC) + alkylamines | B.Braun (Melsungen, Germany) |
| Dodecyldipropylentriamin (DPTA) | Lonza (Basel, Switzerland) |
| Orthophthalic Acid (OPA) | Advanced Sterilization Products (Irvine, CA, USA) |
| Active Substance | Concentration | Contact Time | RF + SD HPV Pseudovirus | RF +SD SV40 |
|---|---|---|---|---|
| EtOH | 60% | 5 min | 4.73 ± 1.49 | 2.00 ± 0.36 |
| 70% | 5 min | 5.81 ± 1.40 | 2.14 ± 0.71 | |
| Isoprop. | 60% | 5 min | 0.91 ± 0.31 | 2.08 ± 0.22 |
| 70% | 5 min | 0.95 ± 0.33 | 2.01 ± 0.54 | |
| GTA | 0.03% | 60 min | 5.89 ± 0.90 | 4.82 ± 0.97 |
| 0.05% | 5 min | 4.52 ± 0.90 | 2.61 ± 0.16 | |
| 0.05% | 60 min | 6.10 ± 1.23 | 4.42 ± 0.11 | |
| PAA | 0.005% | 5 min | 0.69 ± 0.51 | 1.76 ± 0.76 |
| 0.05% | 5 min | 0.66 ± 0.41 | 4.26 ± 0.76 | |
| QAC + alkylamines | 0.5% | 5 min | 0.08 ± 0.14 | 1.10 ± 1.01 |
| 0.5% | 60 min | 0.18 ± 0.16 | 1.92 ± 0.53 | |
| DPTA | 0.5% | 5 min | 4.17 ± 0.35 | 2.64 ± 0.04 |
| 0.5% | 60 min | 4.47 ± 0.82 | 3.29 ± 0.69 | |
| OPA | 0.55% | 5 min | 5.95 ± 0.31 | 4.81 ± 0.28 |
| 0.55% | 12 min | 6.00 ± 0.21 | 4.92 ± 0.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hufbauer, M.; Wieland, U.; Gebel, J.; Steinmann, J.; Akgül, B.; Eggers, M. Inactivation of Polyomavirus SV40 as Surrogate for Human Papillomaviruses by Chemical Disinfectants. Viruses 2021, 13, 2207. https://doi.org/10.3390/v13112207
Hufbauer M, Wieland U, Gebel J, Steinmann J, Akgül B, Eggers M. Inactivation of Polyomavirus SV40 as Surrogate for Human Papillomaviruses by Chemical Disinfectants. Viruses. 2021; 13(11):2207. https://doi.org/10.3390/v13112207
Chicago/Turabian StyleHufbauer, Martin, Ulrike Wieland, Jürgen Gebel, Jochen Steinmann, Baki Akgül, and Maren Eggers. 2021. "Inactivation of Polyomavirus SV40 as Surrogate for Human Papillomaviruses by Chemical Disinfectants" Viruses 13, no. 11: 2207. https://doi.org/10.3390/v13112207
APA StyleHufbauer, M., Wieland, U., Gebel, J., Steinmann, J., Akgül, B., & Eggers, M. (2021). Inactivation of Polyomavirus SV40 as Surrogate for Human Papillomaviruses by Chemical Disinfectants. Viruses, 13(11), 2207. https://doi.org/10.3390/v13112207