
Phage Therapy Related Microbial Succession Associated with Successful Clinical Outcome for a Recurrent Urinary Tract Infection

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Subjects
2.2. Phage Discovery/Purification
2.3. Phage Sequencing Analysis
2.4. Bacterial Genome Assembly and Analysis
2.5. Cocktail Formulation
2.6. Killing Assays
2.7. Luminex Data Analysis and Heatmap Creation
2.8. Synography
2.9. Serum Neutralization
3. Results
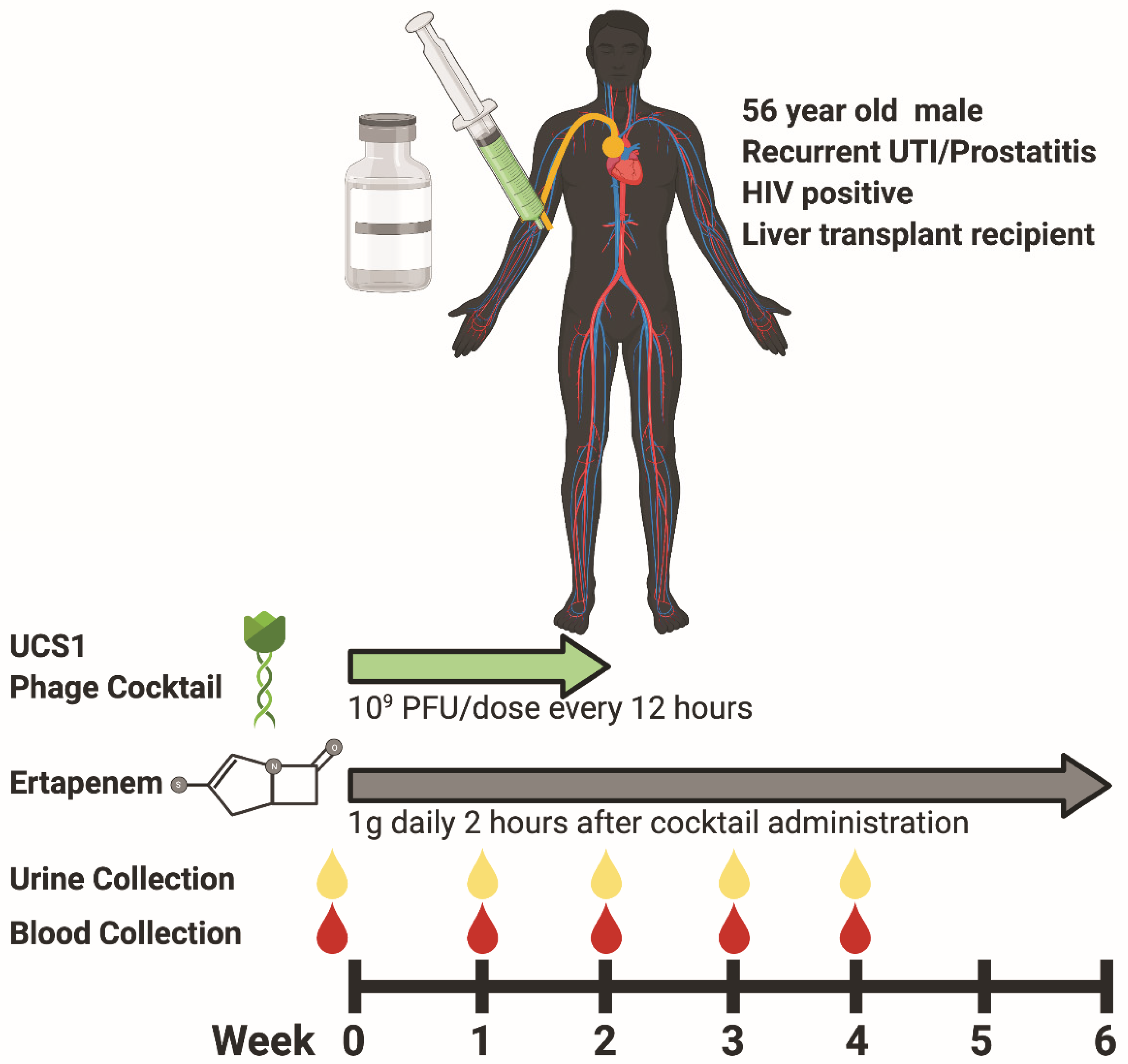
3.1. Participant Characteristics and Phage Therapy Design Modality
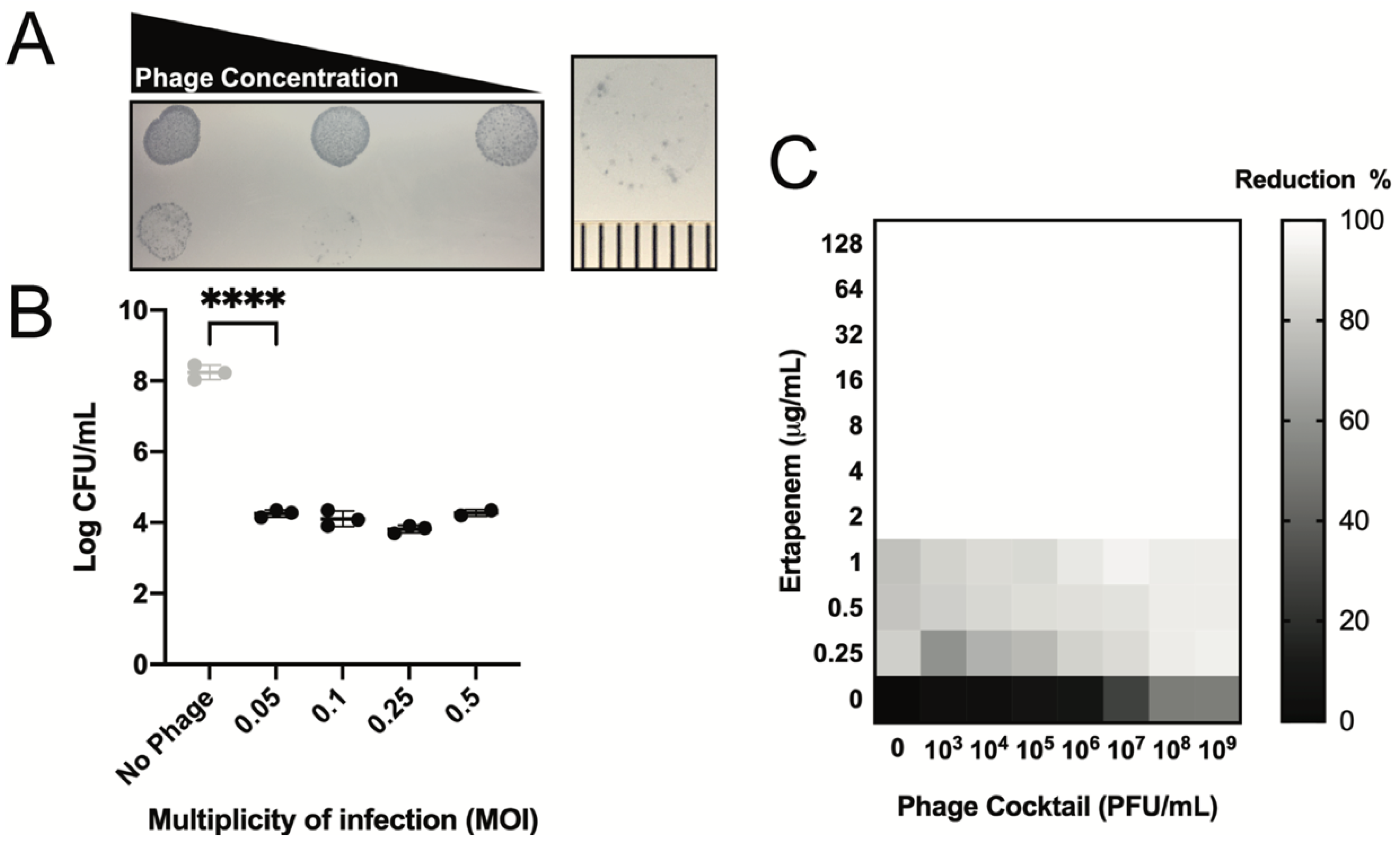
3.2. Safety and Efficacy of Phage Cocktail
3.3. Phage Treatment: Clinical Safety and Efficacy
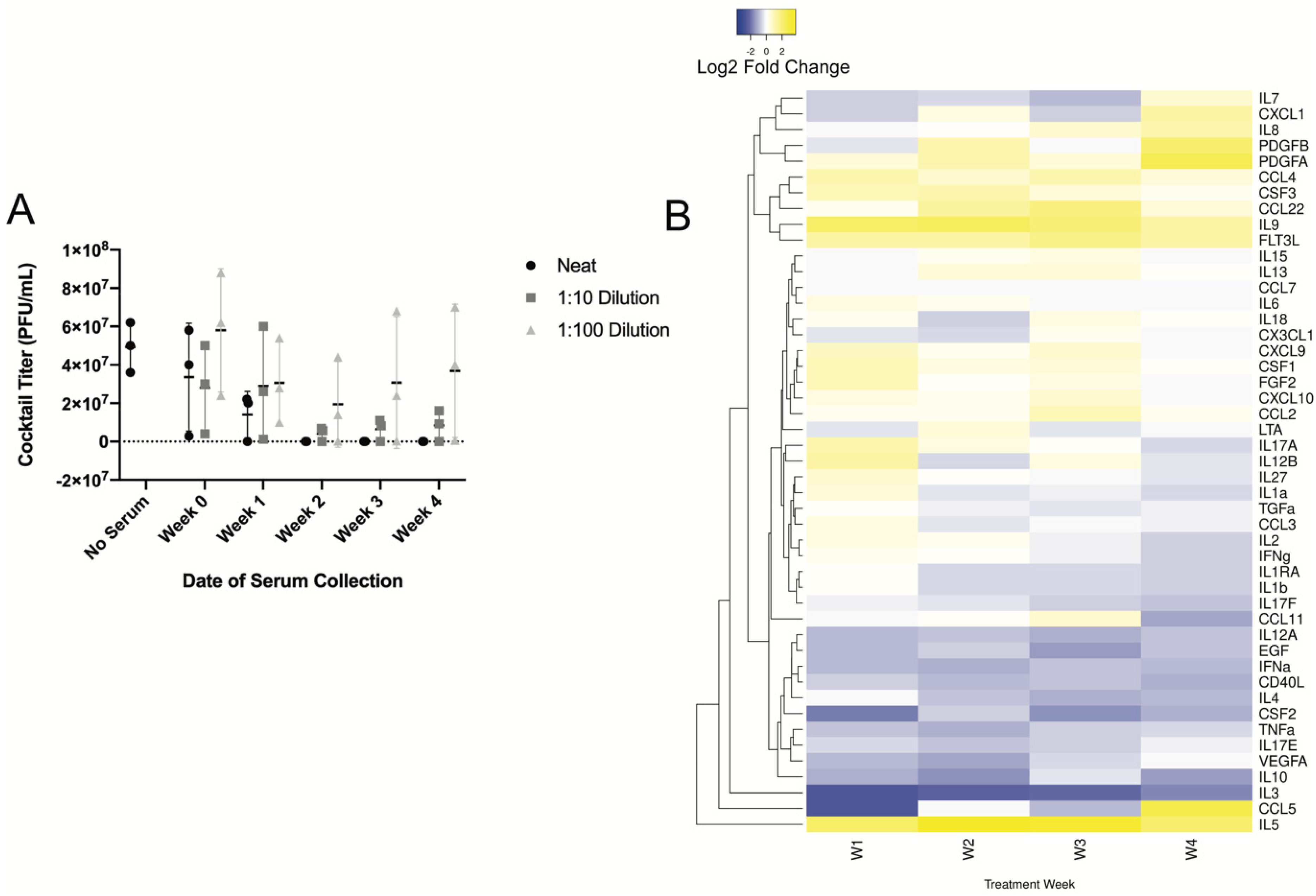
3.4. Longitudinal Assessment of Humoral Response
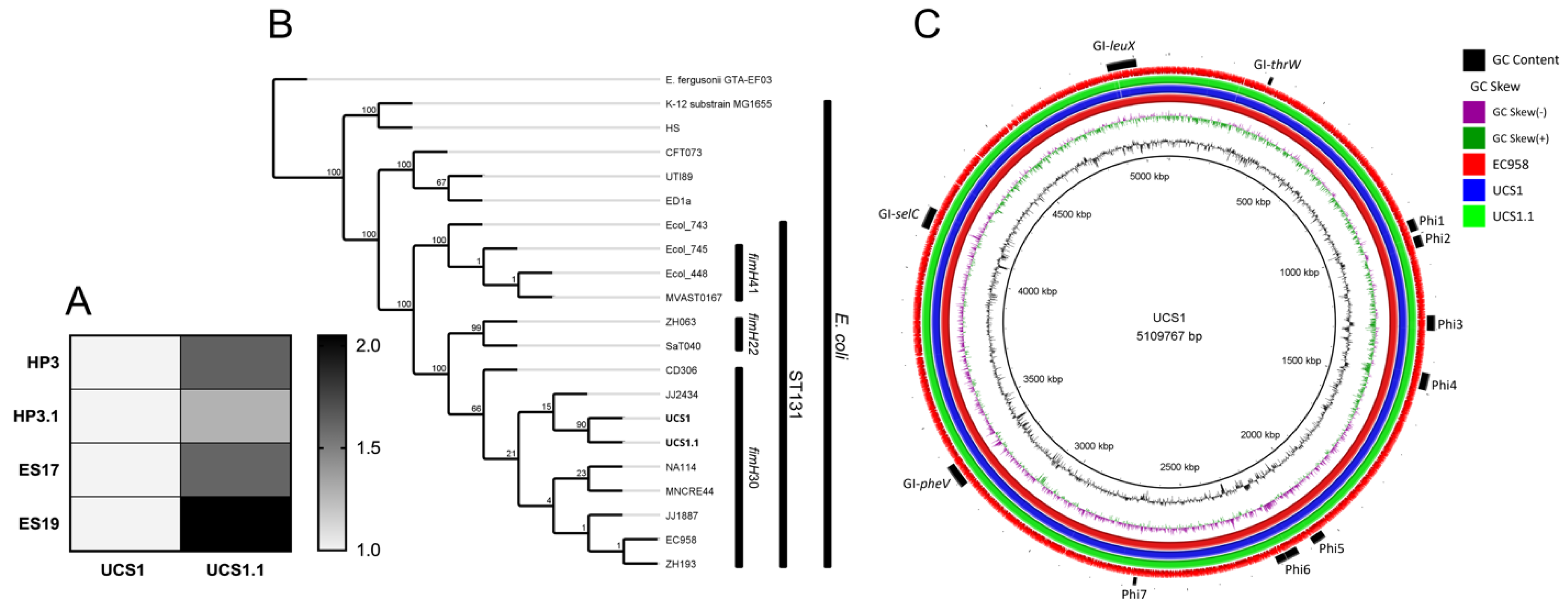
3.5. Assessment of ESBL-Producing E. coli Strain Following Phage Therapy Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hendrix, R.W.; Smith, M.C.M.; Burns, R.N.; Ford, M.E.; Hatfull, G.F. Evolutionary Relationships among Diverse Bacteriophages and Prophages: All the World’s a Phage. Proc. Natl. Acad. Sci. USA 1999, 96, 2192–2197. [Google Scholar] [CrossRef] [PubMed]
- Noble, R.T.; Fuhrman, J.A. Rapid Virus Production and Removal as Measured with Fluorescently Labeled Viruses as Tracers. Appl. Environ. Microbiol. 2000, 66, 3790–3797. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Shlaes, D. Fix the Antibiotics Pipeline. Nature 2011, 472, 32. [Google Scholar] [CrossRef] [PubMed]
- Schooley, R.T.; Strathdee, S. Treat Phage like Living Antibiotics. Nat. Microbiol. 2020, 5, 391–392. [Google Scholar] [CrossRef] [PubMed]
- Green, S.; Ma, L.; Maresso, A. Phage therapy. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The, Netherlands, 2018; p. B9780128012383662000. ISBN 978-0-12-801238-3. [Google Scholar]
- Rostkowska, O.M.; Międzybrodzki, R.; Miszewska-Szyszkowska, D.; Górski, A.; Durlik, M. Treatment of Recurrent Urinary Tract Infections in a 60-year-old Kidney Transplant Recipient. The Use of Phage Therapy. Transpl. Infect. Dis. 2020, 23, e13391. [Google Scholar] [CrossRef]
- Caldwell, J.A. Bacteriophagy in urinary infections following the administration of the bacteriophage therapeutically. Arch. Intern. Med. 1928, 41, 189. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (U.S.). Antibiotic Resistance Threats in the United States, 2019; Centers for Disease Control and Prevention (U.S.): Atalanta, GA, USA, 2019.
- Takhar, S.S.; Moran, G.J. Diagnosis and Management of Urinary Tract Infection in the Emergency Department and Outpatient Settings. Infect. Dis. Clin. N. Am. 2014, 28, 33–48. [Google Scholar] [CrossRef]
- Wagenlehner, F.M.E.; Bjerklund Johansen, T.E.; Cai, T.; Koves, B.; Kranz, J.; Pilatz, A.; Tandogdu, Z. Epidemiology, Definition and Treatment of Complicated Urinary Tract Infections. Nat. Rev. Urol. 2020, 17, 586–600. [Google Scholar] [CrossRef]
- Zhao, F.; Yang, H.; Bi, D.; Khaledi, A.; Qiao, M. A Systematic Review and Meta-Analysis of Antibiotic Resistance Patterns, and the Correlation between Biofilm Formation with Virulence Factors in Uropathogenic E. coli Isolated from Urinary Tract Infections. Microb. Pathog. 2020, 144, 104196. [Google Scholar] [CrossRef]
- De Souza, R.M.; Olsburgh, J. Urinary Tract Infection in the Renal Transplant Patient. Nat. Rev. Nephrol. 2008, 4, 252–264. [Google Scholar] [CrossRef]
- Wagenlehner, F.M.E.; Naber, K.G. Fluoroquinolone Antimicrobial Agents in the Treatment of Prostatitis and Recurrent Urinary Tract Infections in Men. Curr. Urology Rep. 2004, 5, 309–316. [Google Scholar] [CrossRef]
- Green, S.I.; Liu, C.G.; Yu, X.; Gibson, S.; Salmen, W.; Rajan, A.; Carter, H.E.; Clark, J.R.; Song, X.; Ramig, R.F.; et al. Targeting of Mammalian Glycans Enhances Phage Predation in the Gastrointestinal Tract. mBio 2021, 12, e03474-20. [Google Scholar] [CrossRef]
- Salazar, K.C.; Ma, L.; Green, S.I.; Zulk, J.J.; Trautner, B.W.; Ramig, R.F.; Clark, J.R.; Terwilliger, A.L.; Maresso, A.W. Antiviral Resistance and Phage Counter Adaptation to Antibiotic-Resistant Extraintestinal Pathogenic Escherichia Coli. mBio 2021, 12, 20. [Google Scholar] [CrossRef]
- Clokie, M.R.J.; Kropinski, A.; Lavigne, R. (Eds.) Bacteriophages: Methods and Protocols, Volume IV; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 1898, ISBN 978-1-4939-8939-3. [Google Scholar]
- Green, S.I.; Kaelber, J.T.; Ma, L.; Trautner, B.W.; Ramig, R.F.; Maresso, A.W. Bacteriophages from ExPEC Reservoirs Kill Pandemic Multidrug-Resistant Strains of Clonal Group ST131 in Animal Models of Bacteremia. Sci. Rep. 2017, 7, 46151. [Google Scholar] [CrossRef]
- Lo, C.-C.; Chain, P.S.G. Rapid Evaluation and Quality Control of next Generation Sequencing Data with FaQCs. BMC Bioinform. 2014, 15, 366. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A Modular and Extensible Implementation of the RAST Algorithm for Building Custom Annotation Pipelines and Annotating Batches of Genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the All-Bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Laslett, D. ARAGORN, a Program to Detect TRNA Genes and TmRNA Genes in Nucleotide Sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A Better Web Interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and Refined Dataset for Big Data Analysis—10 Years On. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- Mao, C.; Abraham, D.; Wattam, A.R.; Wilson, M.J.C.; Shukla, M.; Yoo, H.S.; Sobral, B.W. Curation, Integration and Visualization of Bacterial Virulence Factors in PATRIC. Bioinformatics 2015, 31, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Pop, M. ARDB--Antibiotic Resistance Genes Database. Nucleic Acids Res. 2009, 37, D443–D447. [Google Scholar] [CrossRef] [PubMed]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The Comprehensive Antibiotic Resistance Database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, J.; Gibson, M.K.; Franzosa, E.A.; Segata, N.; Dantas, G.; Huttenhower, C. High-Specificity Targeted Functional Profiling in Microbial Communities with ShortBRED. PLoS Comput. Biol. 2015, 11, e1004557. [Google Scholar] [CrossRef]
- Gibson, M.K.; Forsberg, K.J.; Dantas, G. Improved Annotation of Antibiotic Resistance Determinants Reveals Microbial Resistomes Cluster by Ecology. ISME J. 2015, 9, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A Better, Faster Version of the PHAST Phage Search Tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed]
- McNair, K.; Bailey, B.A.; Edwards, R.A. PHACTS, a Computational Approach to Classifying the Lifestyle of Phages. Bioinformatics 2012, 28, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.C.E. Mauve: Multiple Alignment of Conserved Genomic Sequence With Rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Joensen, K.G.; Tetzschner, A.M.M.; Iguchi, A.; Aarestrup, F.M.; Scheutz, F. Rapid and Easy In Silico Serotyping of Escherichia Coli Isolates by Use of Whole-Genome Sequencing Data. J. Clin. Microbiol. 2015, 53, 2410–2426. [Google Scholar] [CrossRef]
- Roer, L.; Tchesnokova, V.; Allesøe, R.; Muradova, M.; Chattopadhyay, S.; Ahrenfeldt, J.; Thomsen, M.C.F.; Lund, O.; Hansen, F.; Hammerum, A.M.; et al. Development of a Web Tool for Escherichia Coli Subtyping Based on FimH Alleles. J. Clin. Microbiol. 2017, 55, 2538–2543. [Google Scholar] [CrossRef]
- Clark, J.R.; Maresso, A.M. Comparative Pathogenomics of Escherichia Coli: Polyvalent Vaccine Target Identification through Virulome Analysis. Infect Immun. 2021, 89. [Google Scholar] [CrossRef] [PubMed]
- Alanjary, M.; Steinke, K.; Ziemert, N. AutoMLST: An Automated Web Server for Generating Multi-Locus Species Trees Highlighting Natural Product Potential. Nucleic Acids Res. 2019, 47, W276–W282. [Google Scholar] [CrossRef] [PubMed]
- Seemann, Torsten Rapid Haploid Variant Calling and Core Genome Alignment. Available online: https://github.com/tseemann/snippy (accessed on 12 September 2021).
- Gu Liu, C.; Green, S.I.; Min, L.; Clark, J.R.; Salazar, K.C.; Terwilliger, A.L.; Kaplan, H.B.; Trautner, B.W.; Ramig, R.F.; Maresso, A.W. Phage-Antibiotic Synergy Is Driven by a Unique Combination of Antibacterial Mechanism of Action and Stoichiometry. mBio 2020, 11, e01462-20. [Google Scholar] [CrossRef] [PubMed]
- Gibson, S.B.; Green, S.I.; Liu, C.G.; Salazar, K.C.; Clark, J.R.; Terwilliger, A.L.; Kaplan, H.B.; Maresso, A.W.; Trautner, B.W.; Ramig, R.F. Constructing and Characterizing Bacteriophage Libraries for Phage Therapy of Human Infections. Front. Microbiol. 2019, 10, 2537. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.R.; Schuksz, M.; Esko, J.D. Heparan Sulphate Proteoglycans Fine-Tune Mammalian Physiology. Nature 2007, 446, 1030–1037. [Google Scholar] [CrossRef]
- Pirnay, J.; Blasdel, B.G.; Bretaudeau, L.; Buckling, A.; Chanishvili, N.; Clark, J.R.; Corte-real, S.; Debarbieux, L.; Dublanchet, A.; De Vos, D.; et al. Quality and Safety Requirements for Sustainable Phage Therapy Products. Pharm. Res. 2015, 32, 2173–2179. [Google Scholar] [CrossRef]
- Office of Regulatory Affairs Bacterial Endotoxins/Pyrogens. Available online: https://www.fda.gov/inspections-compliance-enforcement-and-criminal-investigations/inspection-technical-guides/bacterial-endotoxinspyrogens (accessed on 27 December 2020).
- Johnson, J.R.; Johnston, B.; Clabots, C.; Kuskowski, M.A.; Castanheira, M. Escherichia Coli Sequence Type ST131 as the Major Cause of Serious Multidrug-Resistant E. Coli Infections in the United States. Clin. Infect. Dis. 2010, 51, 286–294. [Google Scholar] [CrossRef]
- Johnson, J.R.; Tchesnokova, V.; Johnston, B.; Clabots, C.; Roberts, P.L.; Billig, M.; Riddell, K.; Rogers, P.; Qin, X.; Butler-Wu, S.; et al. Abrupt Emergence of a Single Dominant Multidrug-Resistant Strain of Escherichia Coli. J. Infect. Dis. 2013, 207, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.M.; Ben Zakour, N.L.; Stanton-Cook, M.; Phan, M.-D.; Totsika, M.; Peters, K.M.; Chan, K.G.; Schembri, M.A.; Upton, M.; Beatson, S.A. The Complete Genome Sequence of Escherichia Coli EC958: A High Quality Reference Sequence for the Globally Disseminated Multidrug Resistant E. Coli O25b:H4-ST131 Clone. PLoS ONE 2014, 9, e104400. [Google Scholar] [CrossRef] [PubMed]
- Speck, P.; Smithyman, A. Safety and Efficacy of Phage Therapy via the Intravenous Route. FEMS Microbiol. Lett. 2016, 363, fnv242. [Google Scholar] [CrossRef]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage Treatment of Human Infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef] [PubMed]
- Aslam, S.; Lampley, E.; Wooten, D.; Karris, M.; Benson, C.; Strathdee, S.; Schooley, R.T. Lessons Learned From the First 10 Consecutive Cases of Intravenous Bacteriophage Therapy to Treat Multidrug-Resistant Bacterial Infections at a Single Center in the United States. Open Forum Infect. Dis. 2020, 7, ofaa389. [Google Scholar] [CrossRef]
- Łusiak-Szelachowska, M.; Żaczek, M.; Weber-Dąbrowska, B.; Międzybrodzki, R.; Kłak, M.; Fortuna, W.; Letkiewicz, S.; Rogóż, P.; Szufnarowski, K.; Jończyk-Matysiak, E.; et al. Phage Neutralization by Sera of Patients Receiving Phage Therapy. Viral Immunol. 2014, 27, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Moon, B.; Takaki, S.; Miyake, K.; Takatsu, K. The Role of IL-5 for Mature B-1 Cells in Homeostatic Proliferation, Cell Survival, and Ig Production. J. Immunol. 2004, 172, 6020–6029. [Google Scholar] [CrossRef]
- Kerr, M.A. The Structure and Function of Human IgA. Biochem. J. 1990, 271, 285–296. [Google Scholar] [CrossRef]
- Easton, D.M.; Allsopp, L.P.; Phan, M.-D.; Moriel, D.G.; Goh, G.K.; Beatson, S.A.; Mahony, T.J.; Cobbold, R.N.; Schembri, M.A. The Intimin-Like Protein FdeC Is Regulated by H-NS and Temperature in Enterohemorrhagic Escherichia Coli. Appl. Environ. Microbiol. 2014, 80, 7337–7347. [Google Scholar] [CrossRef]
- Nesta, B.; Spraggon, G.; Alteri, C.; Moriel, D.G.; Rosini, R.; Veggi, D.; Smith, S.; Bertoldi, I.; Pastorello, I.; Ferlenghi, I.; et al. FdeC, a Novel Broadly Conserved Escherichia Coli Adhesin Eliciting Protection against Urinary Tract Infections. mBio 2012, 3, 9. [Google Scholar] [CrossRef]
- Olschläger, T.; Braun, V. Sequence, Expression, and Localization of the Immunity Protein for Colicin M. J. Bacteriol. 1987, 169, 4765–4769. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mysorekar, I.U.; Hultgren, S.J. Mechanisms of Uropathogenic Escherichia Coli Persistence and Eradication from the Urinary Tract. Proc. Natl. Acad. Sci. USA 2006, 103, 14170–14175. [Google Scholar] [CrossRef]
- Rosen, D.A.; Hooton, T.M.; Stamm, W.E.; Humphrey, P.A.; Hultgren, S.J. Detection of Intracellular Bacterial Communities in Human Urinary Tract Infection. PLoS Med. 2007, 4, e329. [Google Scholar] [CrossRef] [PubMed]
- Chibeu, A.; Lingohr, E.J.; Masson, L.; Manges, A.; Harel, J.; Ackermann, H.-W.; Kropinski, A.M.; Boerlin, P. Bacteriophages with the Ability to Degrade Uropathogenic Escherichia Coli Biofilms. Viruses 2012, 4, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Żaczek, M.; Weber-Dąbrowska, B.; Międzybrodzki, R.; Górski, A. Phage Prevalence in the Human Urinary Tract—Current Knowledge and Therapeutic Implications. Microorganisms 2020, 8, 1802. [Google Scholar] [CrossRef] [PubMed]
- Garretto, A.; Miller-Ensminger, T.; Wolfe, A.J.; Putonti, C. Bacteriophages of the Lower Urinary Tract. Nat. Rev. Urol. 2019, 16, 422–432. [Google Scholar] [CrossRef]
- Adaptive Phage Therapeutics, Inc. A Phase I/II Study of Bacteriophage Therapy to Evaluate Safety, Tolerability, and Efficacy of Targeted “Personalized” Bacteriophage Treatments in Patients With Bacterial Infection of the Urinary Tract; Adaptive Phage Therapeutics, Inc.: Gaithersburg, MD, USA, 2020. Available online: clinicaltrials.gov (accessed on 28 December 2020).
- Pirnay, J.-P.; De Vos, D.; Verbeken, G.; Merabishvili, M.; Chanishvili, N.; Vaneechoutte, M.; Zizi, M.; Laire, G.; Lavigne, R.; Huys, I.; et al. The Phage Therapy Paradigm: Prêt-à-Porter or Sur-Mesure? Pharm. Res. 2011, 28, 934–937. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteriophages | HP3 | HP3.1 | ES17 | ES19 |
|---|---|---|---|---|
 100 nm |  |  |  |  |
| Characteristics | ||||
| Morphology | Myovirus | Myovirus | Podovirus | Myovirus |
| Source | Goose/Duck Feces | Directed Evolution | Human Sewage | Human Sewage |
| Genome Size (bp) | 168,188 | 168,195 | 75,134 | 167,088 |
| Coding Sequences (CDS) | 264 | 266 | 124 | 263 |
| Hypothetical Proteins | 26 | 26 | 79 | 24 |
| Proteins with Functional Assignment | 238 | 240 | 45 | 239 |
| Closest Sequenced Relative | Escherichia virus T40 | Escherichia virus T40 | Escherichia phage Eco32 | Escherichia phage HY01 |
| Antibiotic Resistance CDS | NO | NO | NO | NO |
| Bacterial Virulence CDS | NO | NO | NO | NO |
| Attachment Sites | NO | NO | 2 | NO |
| Integrases | NO | NO | NO | NO |
| Lifestyle Prediction | Lytic | Lytic | Likely Lytic | Lytic |
| Titer in Cocktail (PFU/mL) | 7.5 × 109 | 1.35 × 1010 | 1.37 × 1010 | 3.35 × 109 |
| Cocktail Properties | ||||
| Date of Manufacture | 10/23/19 | |||
| Titer (PFU/mL) | 3 × 1010 | |||
| Endotoxin Content (EU/mL) | 685 | |||
| Endotoxin Units per Dose (EU) | 22.83 | |||
| Storage | 4 °C in 2.5 mL Glass Vial | |||
| USP 71 Sterility Testing | No Growth Detected | |||
| Acid Fast Bacteria Testing | No Growth Detected | |||
| Coding Sequence | Locus Tag | Length (bp) | Type | Amino Acid Change | Protein Effect |
|---|---|---|---|---|---|
| Probable lipoprotein YPO0703 | EC958_0641 | 3 | Deletion | Extension | |
| Prophage-associated invasin-like protein | EC958_1349 | 0 | Insertion | Truncation | |
| Prophage-associated invasin-like protein | EC958_1349 | 2 | Deletion | Frame Shift | |
| Prophage-associated invasin-like protein | EC958_1349 | 4 | Substitution | Extension | |
| Large exoproteins involved in heme utilization or adhesion, upaH | EC958_1689 | 9 | Deletion | NTGT → N | Deletion |
| Glutamate decarboxylase, gadB | EC958_1756 | 5 | Deletion | Frame Shift | |
| Putative capsular polysaccaride transport protein, YegH | EC958_2403 | 1 | SNP (transition) | I → M | Substitution |
| Glutamate decarboxylase, gadA | EC958_3921 | 1 | Insertion | Extension | |
| Glutamate decarboxylase, gadA | EC958_3921 | 0 | Insertion | Frame Shift | |
| SisB (ShiA homolog) | EC958_4074 | 1 | SNP (transversion) | Q → L | Substitution |
| Uncharacterized protein YeeP | EC958_4120 | 2 | Deletion | Frame Shift | |
| Antigen 43 | EC958_4121 | 2 | Insertion | N → NAATNVT | Insertion |
| Anti-adapter protein IraM | EC958_4960 | 7 | Deletion | Extension | |
| Antigen 43 | EC958_5138 | 0 | Insertion | Frame Shift | |
| Microcin-E7 immunity protein | EC958_5152 | 2 | Deletion | Frame Shift | |
| Small inner membrane protein, YmgF family | NA | 3 | Substitution | V → R | Substitution |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terwilliger, A.; Clark, J.; Karris, M.; Hernandez-Santos, H.; Green, S.; Aslam, S.; Maresso, A. Phage Therapy Related Microbial Succession Associated with Successful Clinical Outcome for a Recurrent Urinary Tract Infection. Viruses 2021, 13, 2049. https://doi.org/10.3390/v13102049
Terwilliger A, Clark J, Karris M, Hernandez-Santos H, Green S, Aslam S, Maresso A. Phage Therapy Related Microbial Succession Associated with Successful Clinical Outcome for a Recurrent Urinary Tract Infection. Viruses. 2021; 13(10):2049. https://doi.org/10.3390/v13102049
Chicago/Turabian StyleTerwilliger, Austen, Justin Clark, Maile Karris, Haroldo Hernandez-Santos, Sabrina Green, Saima Aslam, and Anthony Maresso. 2021. "Phage Therapy Related Microbial Succession Associated with Successful Clinical Outcome for a Recurrent Urinary Tract Infection" Viruses 13, no. 10: 2049. https://doi.org/10.3390/v13102049
APA StyleTerwilliger, A., Clark, J., Karris, M., Hernandez-Santos, H., Green, S., Aslam, S., & Maresso, A. (2021). Phage Therapy Related Microbial Succession Associated with Successful Clinical Outcome for a Recurrent Urinary Tract Infection. Viruses, 13(10), 2049. https://doi.org/10.3390/v13102049