Chikungunya E2 Protein Produced in E. coli and HEK293-T Cells—Comparison of Their Performances in ELISA

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Design and Production of CHIKV-E2 Protein in E. coli
2.2. Design and Production of CHIKV-E2 Protein in HEK-293T
2.3. Sera Bank and Ethical Considerations
2.4. ELISA
3. Results
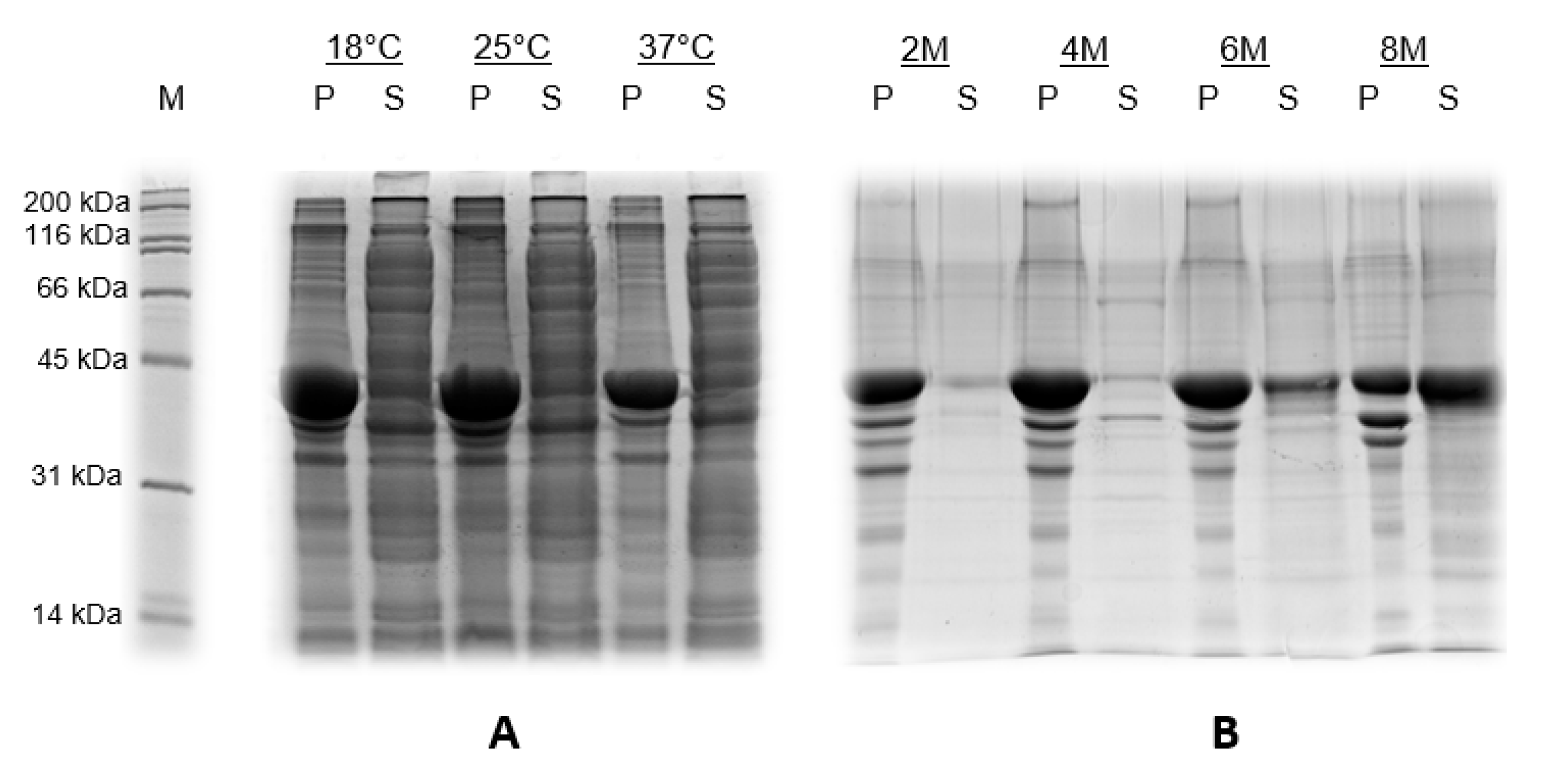
3.1. Production of CHIKV-E2 Protein in E. coli
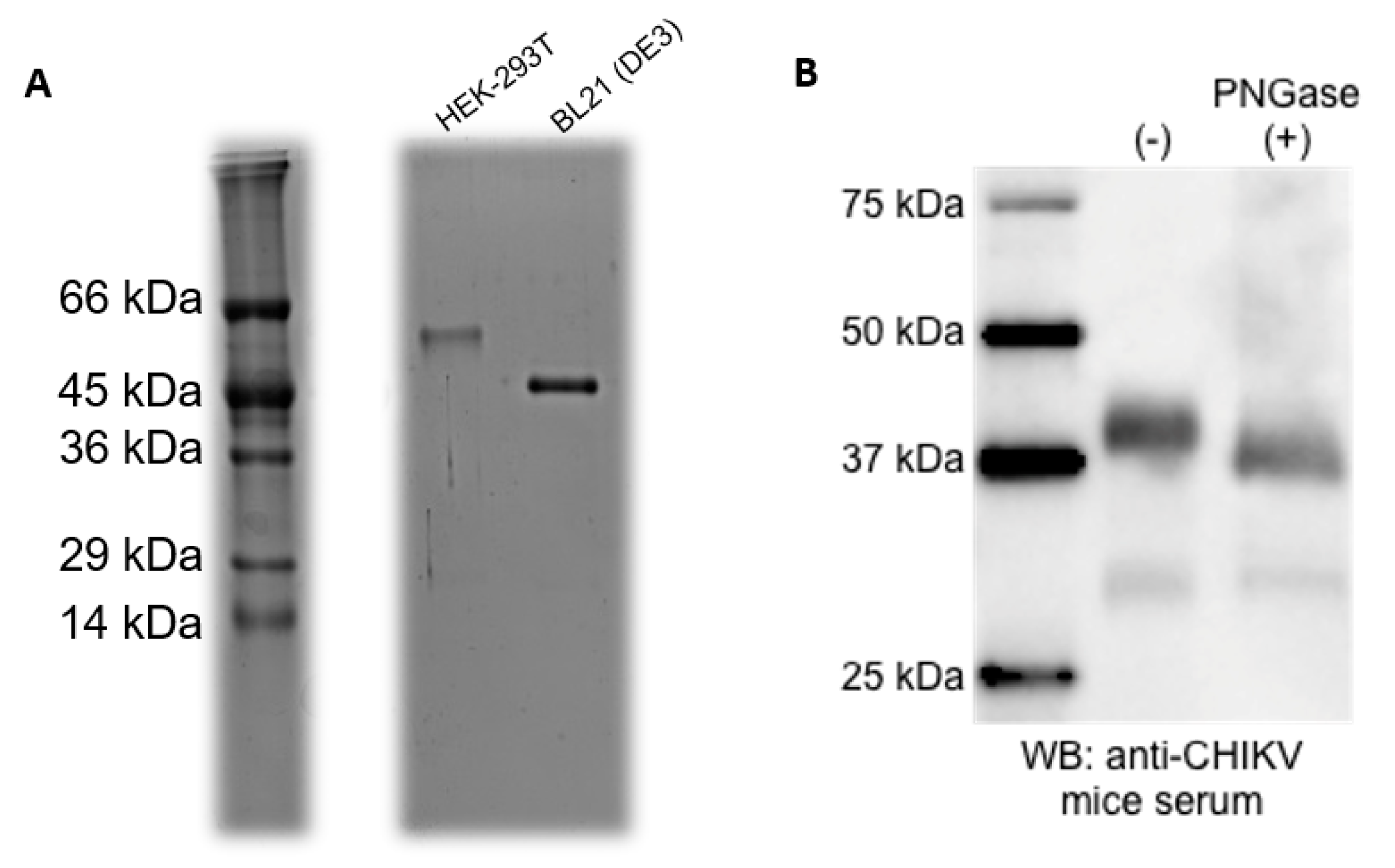
3.2. Purification of the Recombinant Proteins
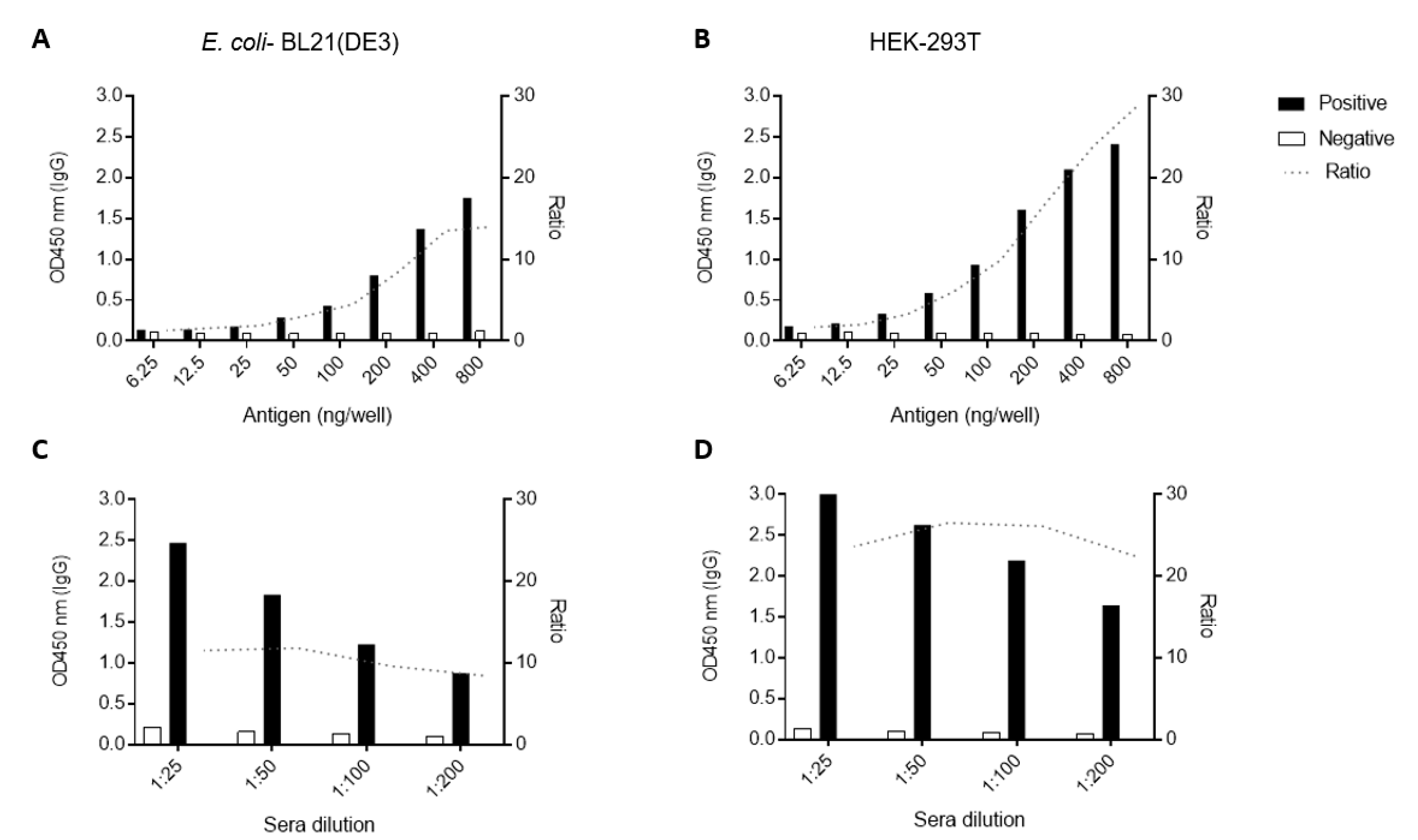
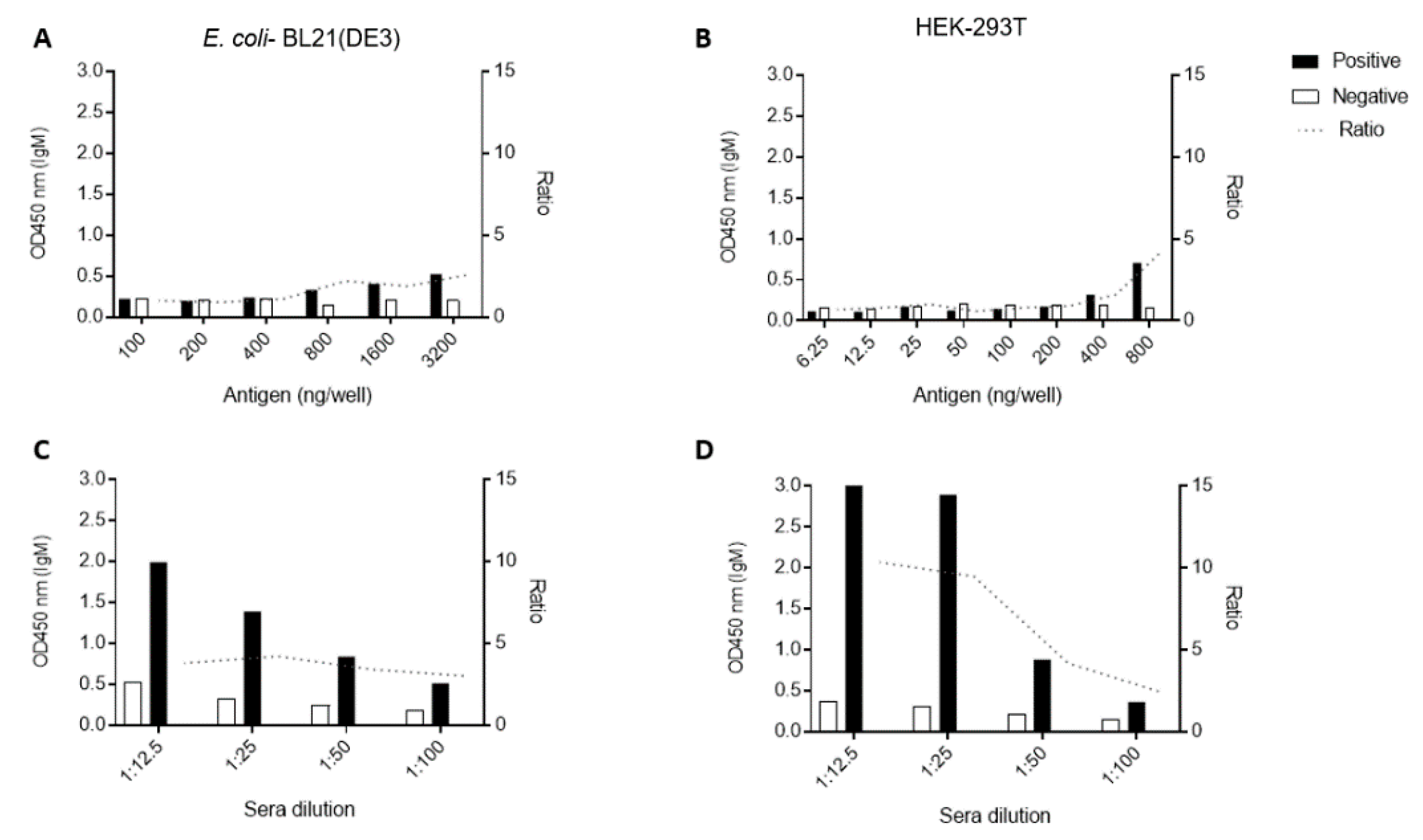
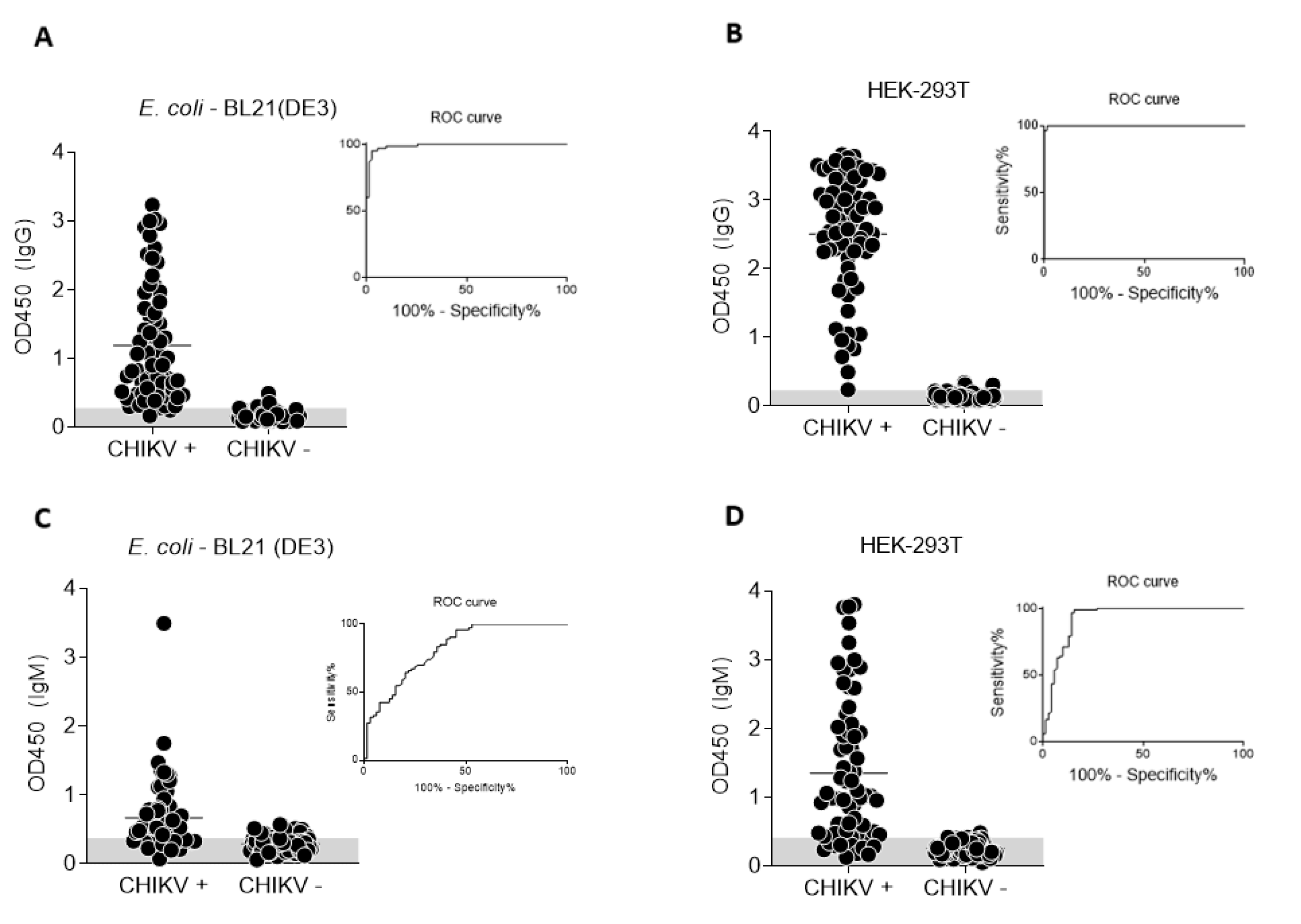
3.3. Standardization of ELISA Using Recombinant CHIKV-E2 from Prokaryotic and Eukaryotic Systems
3.4. Performance of CHIKV-E2 from Prokaryotic and Eukaryotic Systems as Antigens in ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pialoux, G.; Gaüzère, B.A.; Jauréguiberry, S.; Strobel, M. Chikungunya, an epidemic arbovirosis. Lancet Infect. Dis. 2007, 7, 319–327. [Google Scholar] [CrossRef]
- Mendoza, E.J.; Robinson, A.; Dimitrova, K.; Mueller, N.; Holloway, K.; Makowski, K.; Wood, H. Combining anti-IgM and IgG immunoassays for comprehensive chikungunya virus diagnostic testing. Zoonoses Public Health 2019, 66, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.W.; Goodman, C.H.; Holloway, K.; de Salazar, P.M.; Valadere, A.M.; Drebot, M.A. Evaluation of commercially available chikungunya virus immunoglobulin M detection assays. Am. J. Trop. Med. Hyg. 2016, 95, 182–192. [Google Scholar] [CrossRef] [PubMed]
- United States, National Center for Emerging and Zoonotic Infectious Diseases (CDC); Pan American Health Organization. Preparedness and Response for Chikungunya Virus Introduction in the Americas; Pan American Health Organization: Washington, DC, USA, 2011. Available online: https://iris.paho.org/bitstream/handle/10665.2/4009/Chikungunya%20Virus.pdf?sequence=1&isAllowed=y (accessed on 11 August 2020).
- Goodman, C.H.; Russell, B.J.; Velez, J.O.; Laven, J.J.; Nicholson, W.L.; Bagarozzi, D.A., Jr.; Moon, J.L.; Bedi, K.; Johnson, B.W. Development of an algorithm for production of inactivated arbovirus antigens in cell culture. J. Virol. Methods 2014, 208, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Erasmus, J.H.; Needham, J.; Raychaudhuri, S.; Diamond, M.S.; Beasley, D.W.C.; Morkowski, S.; Salje, H.; Salas, I.F.; Kim, D.Y.; Frolov, I.; et al. Utilization of an eilat virus-based chimera for serological detection of chikungunya infection. PLoS Negl. Trop. Dis. 2015, 9, e0004119. [Google Scholar] [CrossRef]
- Spencer, K.A.; Osorio, F.A.; Hiscox, J.A. Recombinant viral proteins for use in diagnostic ELISAs to detect virus infection. Vaccine 2007, 25, 5653–5659. [Google Scholar] [CrossRef]
- Cho, B.; Jeon, B.Y.; Kim, J.; Noh, J.; Kim, J.; Park, M.; Park, S. Expression and evaluation of Chikungunya virus E1 and E2 envelope proteins for serodiagnosis of Chikungunya virus infection. Yonsei Med. J. 2008, 49, 828–835. [Google Scholar] [CrossRef]
- Jin, J.; Liss, N.M.; Chen, D.H.; Liao, M.; Fox, J.M.; Shimak, R.M.; Simmons, G. Neutralizing monoclonal antibodies block chikungunya virus entry and release by targeting an epitope critical to viral pathogenesis. Cell Rep. 2015, 13, 2553–2564. [Google Scholar] [CrossRef]
- Weber, C.; Berberich, E.; von Rhein, C.; Henß, L.; Hildt, E.; Schnierle, B.S. Identification of functional determinants in the chikungunya virus E2 protein. PLoS Neglect. Trop. D 2017, 11, e0005318. [Google Scholar] [CrossRef]
- Vrati, S.; Fernon, C.A.; Dalgarno, L.; Weir, R.C. Location of a major antigenic site involved in Ross River virus neutralization. Virology 1988, 162, 346–353. [Google Scholar] [CrossRef]
- Akahata, W.; Yang, Z.Y.; Andersen, H.; Sun, S.; Holdaway, H.A.; Kong, W.P.; Lewis, M.G.; Higgs, S.; Rossmann, M.G.; Rao, S.; et al. A virus-like particle vaccine for epidemic Chikungunya virus protects nonhuman primates against infection. Nat. Med. 2010, 16, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front Microbiol. 2015, 5, 172. [Google Scholar] [CrossRef] [PubMed]
- Baghban, R.; Farajnia, S.; Ghasemi, Y.; Mortazavi, M.; Zarghami, N.; Samadi, N. New developments in Pichia pastoris expression system, review and update. Curr. Pharm. Biotechnol. 2018, 19, 451–467. [Google Scholar] [CrossRef] [PubMed]
- Gomes, V.A.M.; Carmo, S.T.; Carvalho, S.L.; Bahia, M.F.; Parachin, N.S. Comparison of yeasts as hosts for recombinant protein production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Aricescu, A.R.; Lu, W.; Jones, E.Y. A time- and cost-efficient system for high-level protein production in mammalian cells. Acta Cryst. D Biol. Cryst. 2006, 62 Pt 10, 1243–1250. [Google Scholar] [CrossRef]
- Kost, T.A.; Kemp, C.W. Fundamentals of Baculovirus Expression and Applications. In Advanced Technologies for Protein Complex Production and Characterization. Advances in Experimental Medicine and Biology; Vega, M.C., Ed.; Springer: Cham, Germany, 2016; pp. 187–197. [Google Scholar] [CrossRef]
- Tripathi, N.K.; Priya, R.; Shrivastava, A. Production of recombinant Chikungunya virus envelope 2 protein in Escherichia coli. Appl. Microbiol. Biotechnol. 2014, 98, 2461. [Google Scholar] [CrossRef]
- Tripathi, N.K.; Priya, R.; Shrivastava, A. Immunogenicity of Escherichia coli expressed envelope 2 protein of Chikungunya virus. Bioengineered 2014, 5, 198–203. [Google Scholar] [CrossRef][Green Version]
- Fumagalli, M.J.; de Souza, W.M.; Espósito, D.L.A.; Silva, A.; Romeiro, M.F.; Martinez, E.Z.; da Fonseca, B.A.L.; Figueiredo, L.T.M. Enzyme-linked immunosorbent assay using recombinant envelope protein 2 antigen for diagnosis of Chikungunya virus. Virol. J. 2018, 15, 112. [Google Scholar] [CrossRef]
- Kumar, P.; Pok, K.Y.; Tan, L.K.; Angela, C.; Leo, Y.S.; Ng, L.C. Development and evaluation of baculovirus-expressed Chikungunya virus E1 envelope proteins for serodiagnosis of Chikungunya infection. J. Virol. Methods 2014, 206, 67–75. [Google Scholar] [CrossRef]
- Metz, S.W.; Geertsema, C.; Martina, B.E.; Andrade, P.; Heldens, J.G.; van Oers, M.M.; Goldbach, R.W.; Vlak, J.M.; Pijlman, G.P. Functional processing and secretion of Chikungunya virus E1 and E2 glycoproteins in insect cells. Virol. J. 2011, 8, 353. [Google Scholar] [CrossRef]
- Kim, Y.C.; López-Camacho, C.; Garcia-Larragoiti, N.; Cano-Mendez, A.; Hernandez-Flores, K.G.; Domínguez-Alemán, C.A.; Mar, M.A.; Vivanco-Cid, H.; Viveros-Sandoval, M.E.; Reyes-Sandoval, A. Development of an E2 ELISA methodology to assess chikungunya seroprevalence in patients from an endemic region of mexico. Viruses 2019, 11, 407. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.K.; Shrivastava, A. Recent developments in bioprocessing of recombinant proteins: Expression hosts and process development. Front. Bioeng. Biotechnol. 2019, 7, 420. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Xiang, Y.; Akahata, W.; Holdaway, H.; Pal, P.; Zhang, X.; Diamond, M.S.; Nabel, G.J.; Rossmann, M.G. Structural analyses at pseudo atomic resolution of Chikungunya virus and antibodies show mechanisms of neutralization. eLife 2013, 2, e00435. [Google Scholar] [CrossRef] [PubMed]
- Goffard, A.; Callens, N.; Bartosch, B.; Wychowski, C.; Cosset, F.L.; Montpellier, C.; Dubuisson, J. Role of N-linked glycans in the functions of hepatitis C virus envelope glycoproteins. J. Virol. 2005, 79, 8400–8409. [Google Scholar] [CrossRef] [PubMed]
- Shental-Bechor, D.; Levy, Y. Effect of glycosylation on protein folding: A close look at thermodynamic stabilization. Proc. Natl. Acad. Sci. USA 2008, 105, 8256–8261. [Google Scholar] [CrossRef]
- Ohuchi, M.; Ohuchi, R.; Feldmann, A.; Klenk, H.D. Regulation of receptor binding affinity of influenza virus hemagglutinin by its carbohydrate moiety. J. Virol. 1997, 71, 8377–8384. [Google Scholar] [CrossRef]
- López-Camacho, C.; Kim, Y.C.; Blight, J.; Moreli, L.M.; Montoya-Diaz, E.; Huiskonen, J.T.; Kümmerer, M.B.; Reyes-Sandoval, A. Assessment of immunogenicity and neutralisation efficacacy of viral-vectored vaccines against Chikungunya virus. Viruses 2019, 11, 322. [Google Scholar] [CrossRef]
- Bagno, F.F.; Figueiredo, M.M.; Villarreal, J.; Pereira, G.C.; Godoi, L.C.; da Fonseca, F.G. Undetected Chikungunya virus co-infections in a Brazilian region presenting hyper-endemic circulation of Dengue and Zika. J. Clin. Virol. 2019, 113, 27–30. [Google Scholar] [CrossRef]
- Campbell, M.J.; Machin, D.; Walters, S.J. Medical Statistics: A Textbook for the Health Sciences, 4th ed.; John Wiley & Sons Ltd.: Chichester, UK, 2007; 203p. [Google Scholar]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef]
- Richardson, J.T. The analysis of 2 × 2 contingency tables—Yet again. Stat. Med. 2011, 30, 890–892. [Google Scholar] [CrossRef]
- Kr Verma, A.; Chandele, A.; Kaja, M.-K.; Arulandu, A.; Ray, P. Cloning, expression and purification of Chikungunya virus E2 recombinant protein in E. coli. BMC Infect. Dis. 2014, 14, P65. [Google Scholar] [CrossRef]
- Nigi, I.; Fairall, L.; Schwabe, J.W.R. Expression and purification of protein complexes suitable for structural studies using mammalian hek 293F cells. Curr. Protoc. Protein Sci. 2017, 90, 5.28.1–5.28.16. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, N.; Mukhopadhyay, S. Viral glycoproteins: Biological role and application in diagnosis. Virusdisease 2016, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Prat, C.M.; Flusin, O.; Panella, A.; Tenebray, B.; Lanciotti, R.; Leparc-Goffart, I. Evaluation of commercially available serologic diagnostic tests for chikungunya virus. Emerg. Infect. Dis. 2014, 20, 2129–2132. [Google Scholar] [CrossRef] [PubMed]
- Honjo, T.; Kinoshita, K.; Muramatsu, M. Molecular mechanism of class switch recombination: Linkage with somatic hypermutation. Annu. Rev. Immunol. 2002, 20, 165–196. [Google Scholar] [CrossRef]
- Sterner, E.; Flanagan, N.; Gildersleeve, J.C. Perspectives on anti-glycan antibodies gleaned from development of a community resource database. ACS Chem. Biol. 2016, 11, 1773–1783. [Google Scholar] [CrossRef]
- Cifuentes-Muñoz, N.; Salazar-Quiroz, N.; Tischler, N.D. Hantavirus Gn and Gc envelope glycoproteins: Key structural units for virus cell entry and virus assembly. Viruses 2014, 6, 1801–1822. [Google Scholar] [CrossRef]
- Varki, A.; Gagneux, P. Biological Functions of Glycans. In Essentials of Glycobiology [Internet], 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017; Chapter 7. Available online: https://www.ncbi.nlm.nih.gov/books/NBK453034/ (accessed on 14 March 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IgG-E.coli | IgG-HEK-293T | IgM-E.coli | IgM-HEK-293T | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Reference | Positive | Negative | Sum | Positive | Negative | Sum | Positive | Negative | Sum | Positive | Negative | Sum | |
| Positive | 66 | 2 | 68 | 69 | 1 | 70 | 42 | 10 | 52 | 57 | 10 | 67 | |
| Negative | 2 | 55 | 57 | 3 | 83 | 86 | 17 | 42 | 59 | 1 | 76 | 77 | |
| Sum | 68 | 57 | 125 | 72 | 84 | 156 | 59 | 52 | 111 | 58 | 86 | 144 | |
| Cut-off | 0.279 | 0.225 | 0.328 | 0.409 | |||||||||
| Sensitivity(95% CI) | 97 (90–100)% | 99 (92–100)% | 81 (67–90%) | 85 (74–93)% | |||||||||
| Especificity(95% CI) | 96 (88–100)% | 97 (90–99)% | 71 (58–82)% | 98 (93–100)% | |||||||||
| PPV (95% CI) | 97 (89–99)% | 96 (88–100)% | 72 (62–79)% | 98 (89–100)% | |||||||||
| NPV (95% CI) | 96 (88–99)% | 99 (92–99)% | 81 (70–88)% | 88 (81–93)% | |||||||||
| AC 95% CI) | 97 (92–99)% | 97 (94–99)% | 76 (67–83)% | 92 (87–96)% | |||||||||
| AUC(95% CI) | 0.987 (0.972–1.000) | 0.999 (0.998–1.000) | 0.817 (0.747–0.888) | 0.9251 (0.877–0.974) | |||||||||
| k(95% CI) | 0.936 (0.873–0.998) | 0.948 (0.898–0.998) | 0.515 (0.358–0.673) | 0.845 (0.758–0.932) | |||||||||
| IgG-E.coli | ||||
| Positive | Negative | Sum | ||
| IgG-HEK | Positive | 65 | 2 | 67 |
| Negative | 2 | 78 | 80 | |
| Sum | 67 | 80 | 147 | |
| k (95% CI) = 0.945 (0.892–0.998) | ||||
| IgM-E.coli | ||||
| Positive | Negative | Sum | ||
| IgM-HEK | Positive | 43 | 13 | 56 |
| Negative | 3 | 25 | 28 | |
| Sum | 46 | 38 | 84 | |
| k (95% CI) = 0.607 (0.439–0.774) | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bagno, F.F.; Godói, L.C.; Figueiredo, M.M.; Sérgio, S.A.R.; Moraes, T.d.F.S.; Salazar, N.d.C.; Kim, Y.C.; Reyes-Sandoval, A.; da Fonseca, F.G. Chikungunya E2 Protein Produced in E. coli and HEK293-T Cells—Comparison of Their Performances in ELISA. Viruses 2020, 12, 939. https://doi.org/10.3390/v12090939
Bagno FF, Godói LC, Figueiredo MM, Sérgio SAR, Moraes TdFS, Salazar NdC, Kim YC, Reyes-Sandoval A, da Fonseca FG. Chikungunya E2 Protein Produced in E. coli and HEK293-T Cells—Comparison of Their Performances in ELISA. Viruses. 2020; 12(9):939. https://doi.org/10.3390/v12090939
Chicago/Turabian StyleBagno, Flávia Fonseca, Lara Carvalho Godói, Maria Marta Figueiredo, Sarah Aparecida Rodrigues Sérgio, Thaís de Fátima Silva Moraes, Natália de Castro Salazar, Young Chan Kim, Arturo Reyes-Sandoval, and Flávio Guimarães da Fonseca. 2020. "Chikungunya E2 Protein Produced in E. coli and HEK293-T Cells—Comparison of Their Performances in ELISA" Viruses 12, no. 9: 939. https://doi.org/10.3390/v12090939
APA StyleBagno, F. F., Godói, L. C., Figueiredo, M. M., Sérgio, S. A. R., Moraes, T. d. F. S., Salazar, N. d. C., Kim, Y. C., Reyes-Sandoval, A., & da Fonseca, F. G. (2020). Chikungunya E2 Protein Produced in E. coli and HEK293-T Cells—Comparison of Their Performances in ELISA. Viruses, 12(9), 939. https://doi.org/10.3390/v12090939