1. Introduction
Venezuelan equine encephalitis virus (VEEV), a New World alphavirus, is a member of the
Togaviridae family of RNA viruses. Infection with VEEV can progress to encephalitic disease that is fatal in ~1% of cases [
1,
2,
3] whereas Old World alphavirus infection (e.g., Sindbis, Chikungunya and Semliki Forest viruses) is primarily associated with arthritogenic disease outcomes [
4]. Although VEEV is endemic to South America, its spread to the southern United States has caused periodic outbreaks affecting both equine and human populations, with an estimated 21 outbreaks in the Americas since the 1930s [
5]. VEEV is an arbovirus transmitted naturally by mosquitoes, however aerosol infection is also possible as demonstrated by past occurrences of laboratory-acquired infections [
6] and VEEV has previously been developed as a bioweapon [
7]. Despite being classified as a category B select agent pathogen by the National Institute of Allergy and Infectious Diseases (NIAID), no FDA-approved vaccines or therapeutics against VEEV exist [
2,
8].
The VEEV genome consists of a positive-sense, single stranded RNA molecule ~11.4 kb in length that encodes two polyproteins [
1,
5]. The non-structural polyprotein consists of the four non-structural proteins nsP1-4, that drive viral RNA replication in concert with a range of host proteins [
9]. The structural polyprotein is cleaved into the viral capsid and glycoproteins, which are responsible for packaging genomes into progeny virions [
1,
10]. While multiple activities of nsP1, 2, and 4 have been described, the activities of nsP3 have only recently begun to be elucidated [
9]. Three distinct domains in nsP3 have been categorized; an N-terminal macrodomain conserved among many alphaviruses, which exhibits homology with other RNA viruses, an alphavirus unique domain (AUD), and a C-terminal hypervariable domain (HVD) [
11,
12]. The macrodomain binds ADP-ribose and possesses phosphatase and de-ADP-ribosylation activity [
12,
13]. The AUD encompasses a zinc-ion coordination site and amino acid site mutations in this domain in Sindbis virus (SINV) have revealed conserved, individual cysteine residues that are vital for viral replication [
10]. The HVD contributes to cell-specific preferences for host factor interactions between different alphaviruses [
14]. Hyperphosphorylation of several HVD threonine and serine residues is critical for viral RNA synthesis in both New World and Old World alphaviruses [
14,
15,
16,
17,
18]. Recent work has begun to elucidate the structural and functional characteristics of VEEV nsP3; however, the interplay between nsP3 and host factors is not well understood.
The nuclear factor-κB (NF-κB) signaling pathway regulates the transcription of genes associated with cellular stress, apoptosis, proliferation, and immune responses [
19,
20,
21]. The NF-κB complex is normally inactive, bound by inhibitor of κB (IκB) proteins and sequestered in the cytoplasm of eukaryotic cells [
22]. Upon stimulation, the inhibitor of κB kinase (IKK) complex is activated and phosphorylates IκB proteins, which are subsequently ubiquitinated and degraded [
22], unmasking nuclear localization signals in the p65/p50 subunits of NF-κB to allow nuclear translocation and induction of various target genes [
23]. The IKK complex, comprised of IKKα, IKKβ, and IKKγ is the master regulator of this signaling cascade and plays an important pro-inflammatory role during most viral infections [
24,
25]. Many viruses disrupt the NF-κB signaling cascade either by encoding proteins to evade the pro-inflammatory responses upregulated by the pathway or by directly hijacking the pathway through modulation of IKK complex activity [
26,
27,
28,
29]. For example, Influenza virus and adenovirus trigger NF-κB activation through accumulation of viral proteins in the endoplasmic reticulum and subsequent activation of the IKKβ subunit which is a necessary intermediate for cascade activation [
30]. Similarly, the human T-lymphotropic virus Tax oncoprotein directly interacts with the IKKγ subunit by recruiting kinase MEKK1 for phosphorylation of the IKK complex [
28]. In these two examples, the IKK complex is targeted to control induction of apoptotic regulatory genes triggered by NF-κB activation and promote transcription of cellular growth genes in order to promote viral replication [
30]. Thus, targeted inhibition of NF-κB pathway members, specifically the IKK complex, could exert antiviral effects. For example, cyclopentenone prostanoids directly bind to IKKβ, and treatment has shown reduction in IKK activity, infectious virion production, and viral gene expression in in vitro models following infection with herpes simplex virus-1 [
30]. Collectively, these examples suggest IKK complex and NF-κB are appealing targets for novel antiviral therapeutics.
Previous research has demonstrated NF-κB cascade activation and macromolecular reorganization of the IKK complex, in particular IKKβ, during infection of U-87MG astrocyte cells with the live-attenuated TC-83 vaccine strain of VEEV [
31]. Treatment with the IKKβ inhibitor BAY-11-7082 significantly decreased levels of both infectious virion particles and viral RNA copies in vitro and in vivo [
31], whereas overexpression of IKKβ increased viral titers following infection. Furthermore, infection of IKKβ
−/− cell lines demonstrated a decrease in viral replication, suggesting that IKKβ activity is important for VEEV replication. Additionally, a direct interaction between VEEV nsP3 and IKKβ was confirmed by both mass spectrometry and confocal microscopy [
31]. Inhibition of IKKβ kinase activity decreased viral replication but had no effect on downstream NF-κB activation of transcription [
31], suggesting that VEEV replication is enhanced by an activity of IKKβ, possibly mediated by phosphorylation of one or more viral proteins, potentially nsP3.
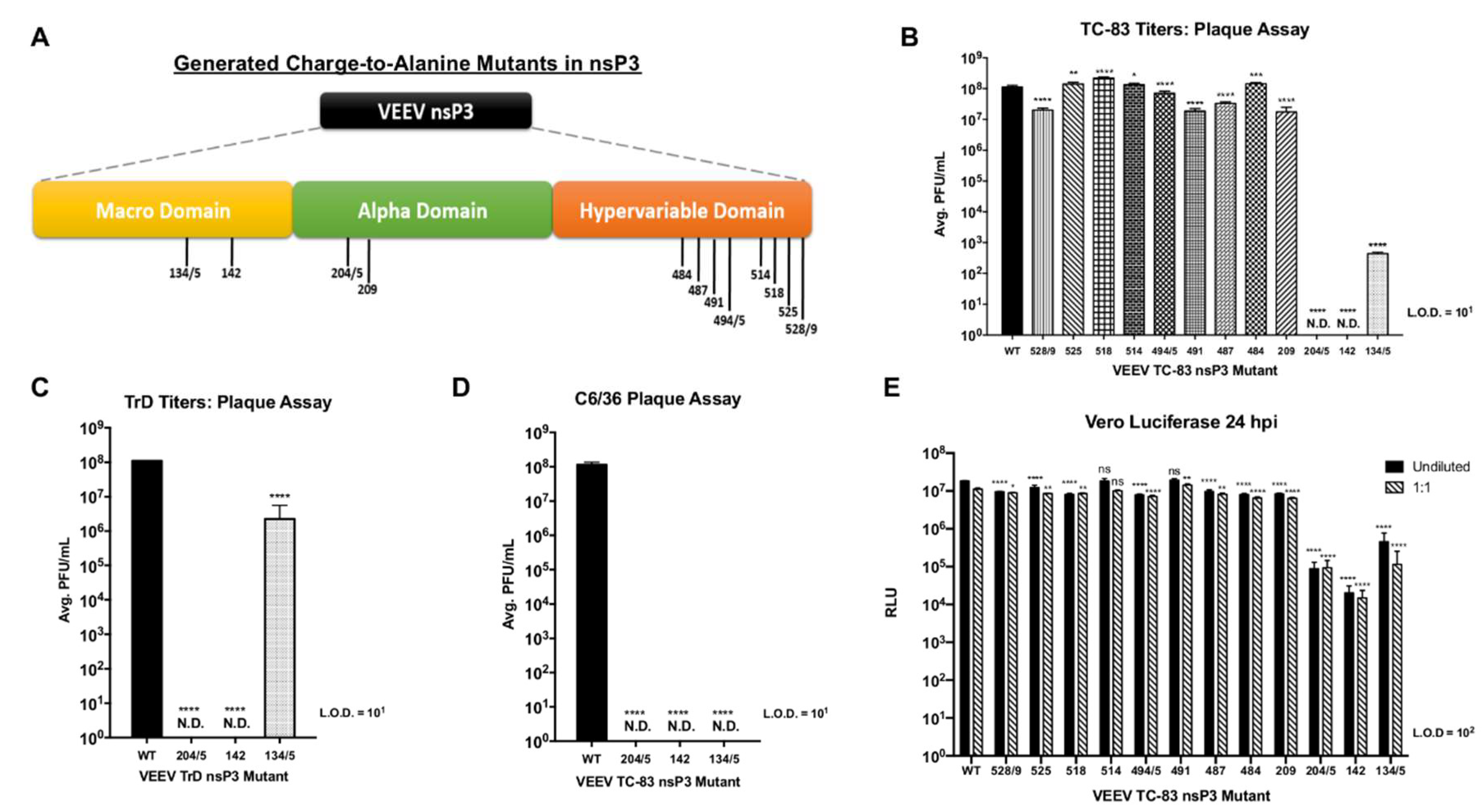
In this study, we investigated IKKβ mediated phosphorylation of nsP3 and evaluated the effects of nsP3 phosphorylation on VEEV replication. NsP3 phosphorylation levels in cells were diminished upon treatment with a small molecule inhibitor of IKKβ, BAY-11-7082, as measured by western blotting. A cell-free, in vitro kinase activity assay performed with IKKβ and purified VEEV nsP3 in the presence of 33P-γ-ATP demonstrated direct phosphorylation of VEEV nsP3 by IKKβ, with the observed phosphorylation signal significantly higher than that of autophosphorylation or a negative control protein, GAPDH. Using mass spectrometry, we identified 18 amino acids in nsP3 that exhibited phosphorylation following VEEV TC-83 infection of U-87MG astrocytes and mouse embryonic fibroblast (MEF) cells competent for IKKβ activity. Inhibition of IKKβ activity using BAY-11-7082 in U-87MG astrocytes resulted in abrogation of phosphorylation of eleven amino acids, whereas infection of IKKβ−/− MEF cells resulted in abrogation of phosphorylation of four amino acids. Charge-to-alanine substitutions were created at 12 phosphorylation sites of interest to further evaluate the effect of nsP3 phosphorylation on viral replication. We identified two nsP3 sites, 204/5 and 142, where alanine substitution abrogated production of detectable infectious particles when introduced in both the TC-83 vaccine strain and the Trinidad Donkey (TrD) epizootic wild-type strain of VEEV. Additionally, alanine substitution at site 134/5 displayed an intermediate phenotype with replication ~105-fold lower when compared to replication of TC-83 and ~30-fold lower when compared to replication of TrD. Furthermore, these mutants were unable to form detectable infectious particles following C6/36 mosquito cell infection. Analysis of positive- and negative-strand RNA levels indicated that phosphorylation-deficient mutants synthesized significantly lower levels of negative-strand during infection when compared to TC-83. Results from a cell-free, in vitro kinase activity assay displayed a reduction in phosphorylation of purified nsP3 mutant proteins with alanine substitutions at sites 204/5, 142 or 134/5 by IKKβ in comparison to wild-type nsP3. Serial passaging of the replication-deficient mutants generated revertants with a replication competent phenotype, attributed to a reversion of the alanine substitutions back to the original phosphorylation-capable TC-83 amino acids. Furthermore, phosphomimetic mutants with aspartic acid substitutions at nsP3 sites 204/5, 142, and 134/5 were generated and tested for replication competence. Phosphomimetic mutant 204/5 restored both infectious particle production and negative-strand RNA synthesis almost comparable to levels observed in TC-83 infected cells, whereas phosphomimetic mutant 134/5 partially restored the ability to produce infectious particles but did not increase negative-strand RNA synthesis. The phosphomimetic mutant 142 did not rescue viral replication. Treatment of phosphomimetic mutants with the IKKβ inhibitor BAY-11-7082 had little effect on levels of infectious particle production. Overall, we identify IKKβ as the first reported host kinase capable of phosphorylating VEEV nsP3 on residues 204/5, 142, and 134/5 and demonstrate that phosphorylation of residues 204/5 is essential for negative-strand synthesis while phosphorylation of residues 134/5 may be important for infectious particle production.
2. Materials and Methods
2.1. Cell Culture
Vero African green monkey kidney cells (ATCC, CCL-81), U-87MG human astrocytoma cells (ATCC, HTB-14), C6/36
Aedes albopictus mosquito cells (ATCC, CRL-1660), and BHK-21 baby hamster kidney cells (ATCC, CCL-10) were obtained from the American Type Culture Collection (Manassas, VA, USA). Inhibitory κB kinase KO (IKKβ
−/−) and WT MEF cells were a kind gift from Dr. Cynthia Masison from NIH/NCI [
32,
33]. Knockout of IKKβ expression in these cells was previously validated [
31]. U-87MG, Vero, BHK-21, and MEF cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, 112-013-101CS, Quality Biological, Gaithersburg, MD, USA) supplemented with 4.5 g/L glucose, 2 mM L-glutamine (MT2005CI, FisherSci, Chicago, IL, USA,), 10% heat-inactivated fetal bovine serum (10437028, ThermoFisher, Carlsbad, CA, USA) for U-87MG and MEF cells or 5% heat-inactivated fetal bovine essence (10805-184, VWR, Dixon, CA, USA) for Vero and BHK-21 cells with 10 ug/mL streptomycin and 10 U/mL penicillin (45000-652, VWR). C6/36 cells were cultured in Eagle’s Minimum Essential Medium (EMEM, 670086, VWR) supplemented with 10% heat-inactivated fetal bovine serum, 10 ug/mL streptomycin and 10 U/mL penicillin. All cell lines were cultivated at 37 °C and 5% CO
2.
2.2. Expression and Purification of VEEV nsP3
Bacterial expression plasmids encoding VEEV TC-83 nsP3 wild-type (NCBI reference sequence L01443) or mutant nsP3 sequences was constructed using the pET-28a(+) vector containing an N-terminal 6X His-tag sequence and an internal T7-tag sequence with a thrombin cleavage site lac repressor and operator by Twist Biosciences (San Francisco, CA, USA). Expression and purification of nsP3 using E. coli was performed by Reaction Biology Corp. (Malvern, PA, USA).
2.3. Kinase Profiling Assay
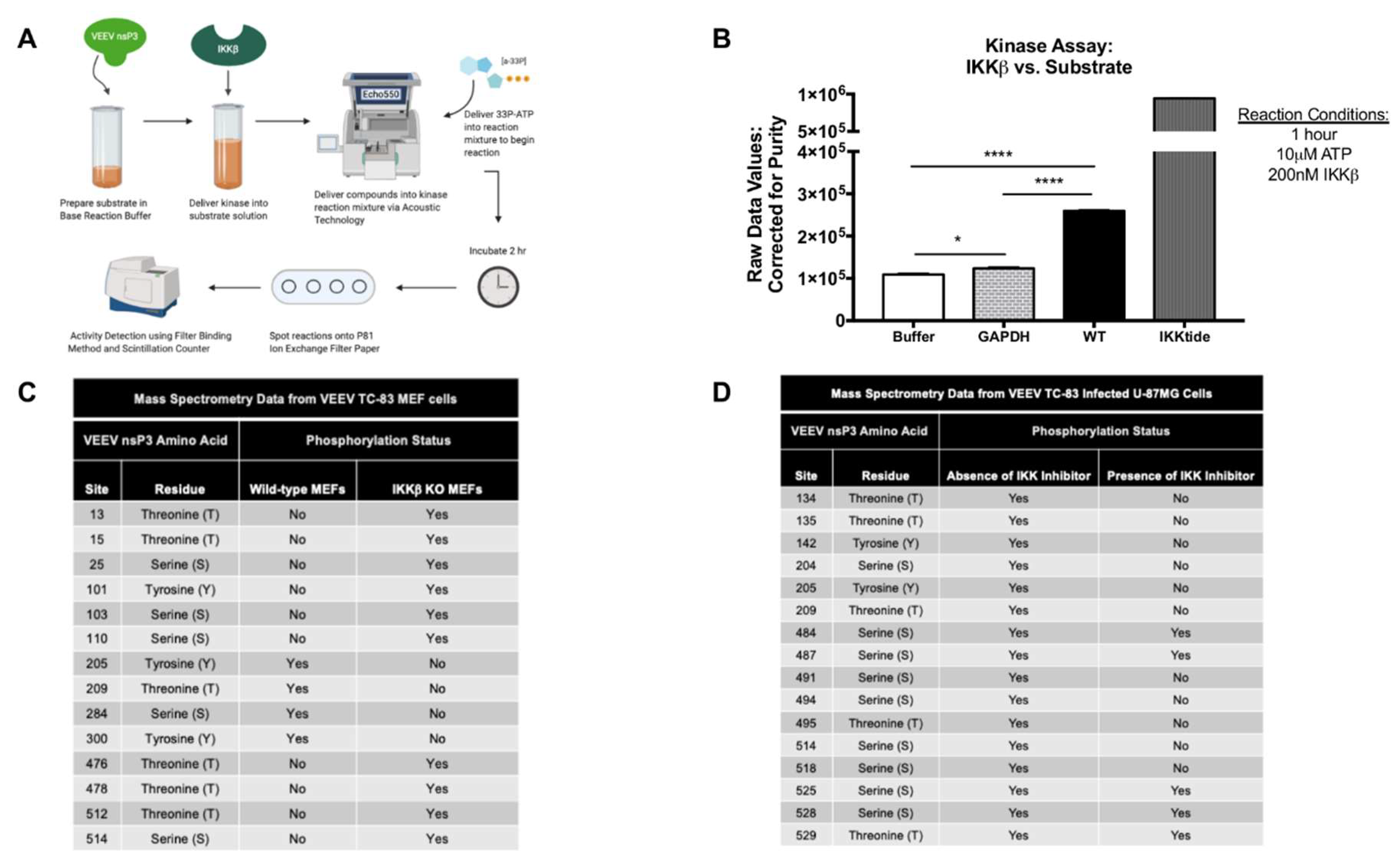
Kinase HotSpot™ Assay for nsP3 substrate vs. IKKβ (Cat #: IKKβ/IKBKB) was performed by Reaction Biology Corp. Briefly, purified VEEV nsP3 was diluted to a concentration of 2 μM in base reaction buffer in duplicate (20 mM Hepes (pH 7.5), 10 mM MgCl2, 1 mM EGTA, 0.02% Brij35, 0.02 mg/mL BSA, 0.1 mM Na3VO4, 2 mM DTT, 1% DMSO). Purified IKKβ kinase was subsequently delivered into the substrate solution at a dosage of 200 nM and mixed gently. The compound mixture was then delivered into a kinase reaction mixture using nanoliter range acoustic technology (Echo550, Reaction Biology Corp., Malvern, PA, USA) and incubated for 20 min at room temperature. 33P-γ-ATP, with a specificity of 10 mCi/mL, was delivered into the reaction mixture at a concentration of 10 μM which was subsequently incubated for 1 h at room temperature. Reactions were spotted on P81 ion exchange paper and kinase activity was subsequently detected by the filter-binding method after unreacted phosphate was removed by washing. High throughput screening using a scintillation counter quantitated kinase activity. A negative control reaction contained the IKKβ kinase alone in the absence of the nsP3 substrate (auto-phosphorylation). An inert negative control protein, GAPDH (ab82633, Abcam, Cambridge, MA) diluted to 2 μM in base reaction buffer, was also evaluated with IKKβ in the assay for evaluation of non-specific effects of the kinase. Reaction Biology Corp. positive control for IKKβ activity was a small peptide, IKKtide, used as substrate at 20 μM vs. IKKβ for efficiency evaluation. Graphs display the raw values of 33P-γ-ATP counts detected by the scintillation counter and corrected for purity of the input nsP3 substrate proteins.
2.4. Viruses and Plasmids
A plasmid encoding the full-length VEEV vaccine strain TC-83 genome was modified to express nano Luciferase as a cleavable component of the structural polyprotein (TaV-nLuc) as previously described [
34,
35], and additional nsP3 mutations were generated in this backbone using PCR mutagenesis (Quick Change Kit, Invitrogen, Carlsbad, CA, USA). A pCAGGS-backbone plasmid expressing N-terminally HA-tagged nsP3 of VEEV ZPC738 was previously described [
36]. VEEV TC-83 virus not expressing nano Luciferase was obtained from BEI Resources [
37].
2.5. In Vitro Transcription
Plasmids encoding nsP3 mutant viruses were linearized using MluI digestion and purified using a Minelute PCR Purification Kit (28006, Qiagen, Louisville, KY, USA,). RNA was transcribed from linearized DNA using an Invitrogen Sp6 MegaScript Kit (AM1330, FisherSci,) per manufacturer’s instructions. RNA was purified using a Qiagen RNeasy Kit (74104) and quantitated using a NanoDrop spectrophotometer (ThermoFisher, Waltham, MA, USA).
2.6. Electroporation and Viral Cultivation
BHK-21 or Vero cells were seeded and grown to 90% confluency in T-75 flasks. Electroporation with 1µg of in vitro transcribed RNA was performed using BHK-21 cells for TC-83 and Vero cells for TrD at 860 V, 25 µF, 950 Ω in BTX 0.2 cm gap cuvettes (USA Scientific, Ocala, FL, USA, 9104-5050) using an ECM 630 Electroporation System (BTX). Electroporated cells were added to a T-75 flask and supernatants collected at 6, 18, and 24 h post electroporation to harvest viral stocks. Supernatants were subjected to 0.2-micron filtering (09-719-006, FisherSci) and frozen at −80 °C.
2.7. Viral Infections and IKKβ Inhibitor Treatment
For all infections, the appropriate cell line was seeded in 96-well plates 24 h prior as follows: U-87MG’s at 1 × 104 and Vero at 5 × 104 per well. C6/36 cells were seeded in 24-well plates 24 h prior at 1 × 105 per well. Vero cells were seeded in 6-well plates 24 h prior at 5 × 105 per well. Viruses were either undiluted, diluted 1:1 in the appropriate culture media, or diluted to the indicated multiplicity of infection (MOI). After a 1-h infection, virus was removed and the appropriate cell media was added. Plates were incubated at 37 °C, 5% CO2 for indicated durations. For inhibitor treatment, U-87MG and MEF cells were seeded at a density of 3 × 105 cells per well in 6-well plates. The IKKβ inhibitor BAY-11-7082 (S2913, SelleckChem, Houston, TX, USA) was dissolved in DMSO, diluted in media for a final concentration of 1 µM for U-87MG and MEF cells or 10 µM for Vero cells and cells were pre-treated for 2 h prior to infection. U-87MG and MEF cells were infected with VEEV TC-83 for 1 h at indicated MOI. Vero cells were infected with viruses as described above for 1 h. Conditioned media containing the inhibitor or standard media was added to cells after removal of virus. Plates were incubated at 37 °C, 5% CO2 for indicated durations.
2.8. Plaque Assay
Vero cells were plated in 12-well plates at a density of 2 × 105 cells/well for 24 h. Viral stocks or infection supernatants were serially diluted to 10−8 in culture media and overlaid on cells for 1 h. Cells were covered with Eagle’s Minimum Essential Medium (without phenol red, supplemented with 5% fetal bovine essence, non-essential amino acids, 1 mM sodium pyruvate (45000-710, VWR), 2 mM L-glutamine, 20 U/mL penicillin, and 20 µg/mL streptomycin) containing 0.6% agarose. 48 h post-infection, cells were fixed with 10% formaldehyde (F79P-4, FisherSci) for 1 h. Medium was removed, wells were washed with diH2O, and cells were stained with a 1% crystal violet (FisherSci, C581-25) in a 20% ethanol solution (BP2818-4, FisherSci). For plaque assay of VEEV TrD nsP3 mutants, virus was serially diluted in virus diluent (phosphate-buffered saline, 1% donor calf serum) and overlaid on BHK-J cells seeded at a density of 1 × 106 cells/well in 6-well plates for 24 h. Plates were stained with neutral red and plaques were counted. All viral entities were titered in triplicate unless otherwise specified.
2.9. Luciferase and Bradford Protein Assay
At indicated time points, supernatants were collected or cellular lysates were obtained using 1X Passive Lysis Buffer (E1941, Promega, Madison, WI, USA) and the Nano-Glo Luciferase Assay System (N1130, Promega) was used to measure luciferase activity per the manufacturer’s instructions. An aliquot of cellular lysates were mixed with Bradford Reagent (5000006, Bio-Rad, Hercules, CA, USA) per the manufacturer’s instructions. A standard curve for total protein was established using Bovine Serum Albumin (BSA, BP1600, FisherSci) diluted in Passive Lysis Buffer at concentrations of 1, 2.5, 5, 10, 15, 20, 25 µg/µL. Mock-infected cells were used to establish the limit of detection for luciferase assays. Luminescence and absorbance was measured using a DTX 880 multimode plate reader (Beckman Coulter, Brea, CA, USA). Intracellular luciferase was normalized to total µg of protein. Overall, sensitivity of the DTX 880 multimode plate reader was somewhat limited due to the age and performance of the machine with lower than anticipated RLU values observed following infection.
2.10. RNA Extraction
At indicated time points, cells were washed with phosphate buffered saline (PBS, L0119-0500, VWR) and lysed with TRIzol Reagent (Invitrogen). Intracellular RNA was extracted using MagBead Direct-zol RNA kit or Direct-zol Miniprep RNA kit (Zymo Research, Irvine, CA, USA) per the manufacturer’s instructions. Extracted viral RNA was stored at −80 °C prior to use.
2.11. Primer/Probes and cDNA
All primers and probes for detection of viral RNA were designed and obtained from Integrated DNA Technologies (IDT) utilizing the PrimerQuest Tool. Primer/probe sets contained a double-quenched ZEN/IBFQ probe with a 6-FAM fluorescent dye attachment at the 5′ end. Capsid primer/probe set, initially described by Julander et al. [
38] was used and is represented in
Table 1. Quantitative data for nsP3 is represented for each non-structural RT-PCR figure and the primer/probe set is represented in
Table 1. 18S rRNA endogenous control primer/probe set was obtained from ThermoFisher (4333760T). cDNA specific to negative-strand RNA for VEEV TC-83 was generated using a forward primer containing a T7 promoter sequence attached at the 5′ end and a high-capacity cDNA reverse transcription kit (4368814, ThermoFisher) per the manufacturer’s instructions. For qPCR of negative-strand viral RNA, forward primer specific to the T7 promoter sequence and reverse primer specific to VEEV TC-83 were utilized. All primer sets for negative-strand viral RNA quantitation are represented in
Table 1.
2.12. Semi-Quantitative RT-PCR
Semi-quantitative RT-PCR for targets of capsid, nsP3, and 18S used thermal cycling conditions adapted from Verso 1-step RT-qPCR kit (AB4101C, ThermoFisher) per the manufacturer’s instructions: 1 cycle at 50 °C for 20 min, 1 cycle at 95 °C for 15 min, 40 cycles at 95 °C for 15 s with 51 °C (capsid), 59 °C (nsP3), 60 °C (18S) for 1 min using StepOnePlus™ Real Time PCR system. qRT-PCR for detection of viral negative-strand used thermal cycling conditions adapted from PowerUp SYBR Green (A25742, ThermoScientific) per the manufacturer’s instructions: 1 cycle at 50 °C for 2 min, 1 cycle at 95 °C for 2 min, 40 cycles at 95 °C for 15 s, 60 °C for 15 s and 72 °C for 1 min using StepOnePlus™ Real Time PCR system. A no template cDNA control and mock infections were included for all analyses and established the limits of detection. Semi-quantitative RT-PCR values were calculated using the ΔΔCt method [
39] with viral entities normalized to 18S levels.
2.13. Transfections
U-87MG cells were seeded at a density of 3 × 105 cells per well in 6-well plates. Transfections used Attractene Transfection Reagent (301005, Qiagen) with 1 µg of DNA plasmid transfected per well as per manufacturer’s instructions. The IKKβ inhibitor, BAY-11-7082, was diluted to 1 µM with culture media and added to the appropriate treatment well at the time of transfection. A pcDNA3.1 (+) plasmid was included as a negative control. Plates were incubated at 37 °C, 5% CO2 for 24 h.
2.14. Western Blot
For preparation of whole cell lysates from infected or transfected cells, media was removed and cells washed twice with PBS. Cells were lysed with clear lysis buffer (CLB) supplemented with 10 mM 1,4-dithiothreitol (DTT, P2325, Invitrogen). CLB contained 50 mM Tris-HCL at pH 7.5 (351-006-101, Quality Biological), 120 mM NaCl (S271-500, FisherSci), 5 mM EDTA (351-027-721, Quality Biological), 0.5% NP-40 (492016, EMD Millipore, Burlington, MA, USA), 50 mM NaF (S7920, Sigma-Aldrich, Milwaukee, WI, USA), 0.2 mM Na3VO4 (450243, Sigma-Aldrich), protease inhibitor tablet (88666, ThermoFisher) and phosphatase inhibitor cocktail (78420, ThermoFisher). Protein lysates were vortexed every 5 min for 20 min and supernatants were collected after centrifugation at 10,000 rpm for 10 min at 4 °C. Sample supernatants were diluted 1:1 in 2× Laemmli buffer (1610737, Bio-Rad) supplemented with DTT and boiled for 10 min. Lysates were separated on 4–20% Tris-Glycine Gels and transferred to polyvinyl difluoride (PVDF) membranes via wet transfer for 2 h at 250 mA. Membranes were blocked at room temperature for 30 min using 5% BSA in Tris-buffered saline with 0.1% Tween-20 (TBS-T). Anti-phospho serine antibody (ab9332, Abcam), anti-phosphothreonine antibody (ab9337, Abcam), anti-phospho tyrosine antibody (ab17302, Abcam, 61-5800, Invitrogen) or anti-HA tag antibody (ab18181, Abcam) were diluted in 5% BSA in TBS-T at 4 µg/mL, 2 µg/mL, 1:1000, and 1:1000 respectively, and incubated on individual membranes overnight at 4 °C. Following 3 5-min washes with TBS-T, membranes were incubated with respective secondary HRP-conjugated antibodies (PI32460, FisherSci) diluted 1:10,000 in 5% BSA in TBS-T at room temperature for 1 h and then washed twice with TBS-T and twice with TBS. Membranes were imaged using SuperSignal West Femto Maximum Sensitivity Substrate Kit (34095, ThermoFisher) and a Bio-Rad Molecular Imager ChemiDoc XRS system. Phosphorylation signals were calculated using NIH ImageJ software and values were normalized to both the actin loading control and the HA-tag for all lanes and quantitation is presented as percent fold-change vs. wild-type.
2.15. Liquid Chromatography-Mass Spectrometry
LC-MS/MS was performed as previously described [
40]. Briefly, VEEV TC-83 infected U-87MG or MEF cells were trypsinized (25200056, ThermoFisher) and washed twice with PBS using a centrifugal spin of 1500 rpm for 5 min during each wash. Cellular pellets were lysed using 8 M urea, disulfide bonds were reduced with 1 M DTT and alkylated with iodacetamide. Trypsin digestion was performed for 4 h at 37 °C and peptides were eluted with ZipTip purification (Z720070, Millipore,). LTQ-tandem MS/MS with nanospray reverse-phase liquid chromatography (ThermoFisher) was performed. Post sample injection, column washes proceeded for 5 min with 0.1% formic acid at 200 nL/min. Peptides were then eluted on a 50-min linear gradient from 0–40% acetonitrile in 0.1% formic acid followed by a 5-min holding step in 80% acetonitrile in 0.1% formic acid. LTQ-MS operated in data-dependent mode and each full MS scan was followed with five MS-MS scans where the five most abundant molecular ions were dynamically selected and fragmented by collision-induced dissociation using normalized collision energy of 35%. Mass spectra were fitted against NCBI reference sequence NP_740698.1 or L01443 for putative VEEV nsP3 post translational modification analysis with Sequest Bioworks software (ThermoFisher).
2.16. Serial Passaging of Virus
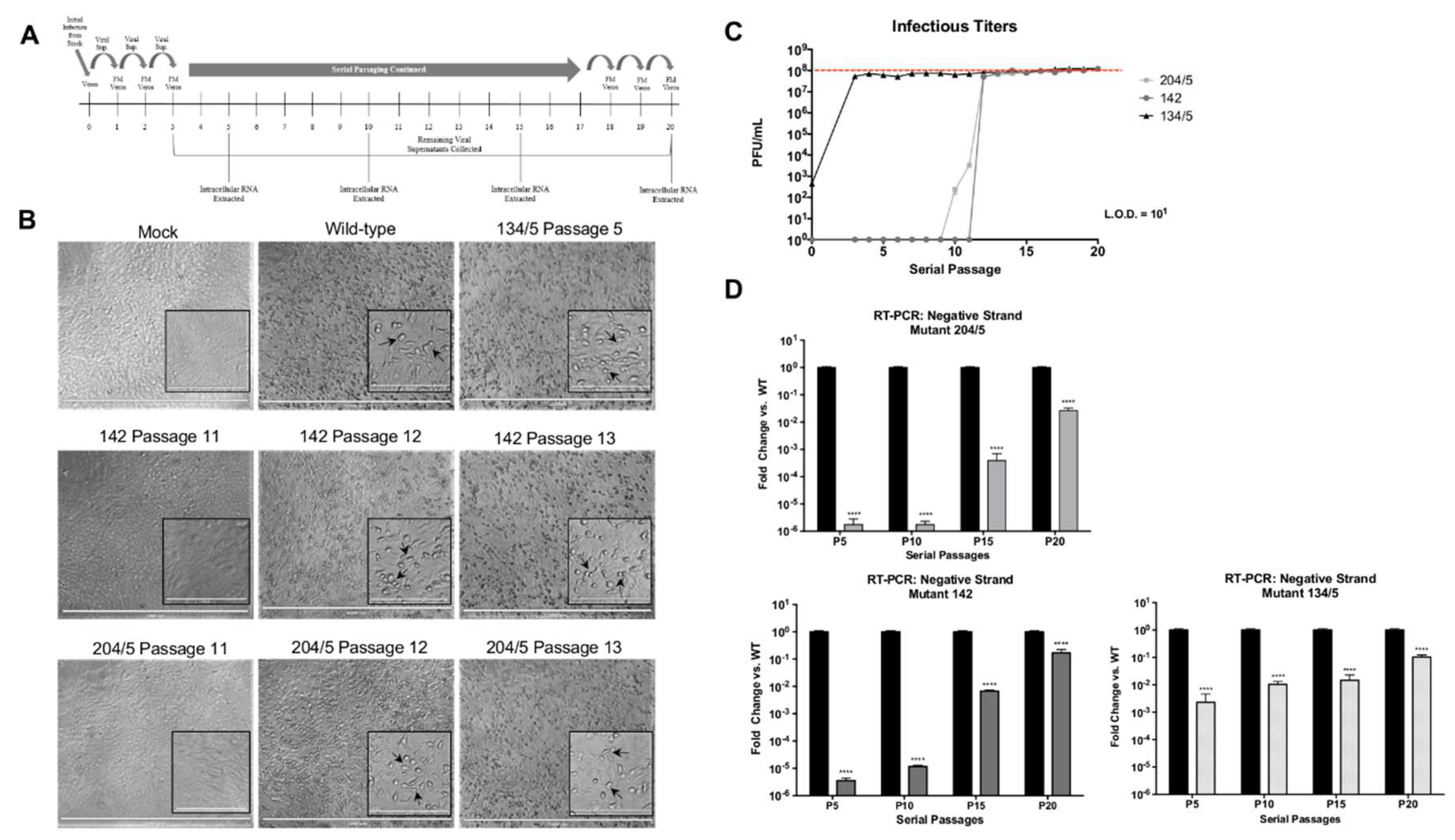
Vero cells were seeded in 6-well plates at varying densities of 5 × 105, 2.5 × 105, and 1.5 × 105 per well to ensure 90% confluency prior to infection. On the first day of infection, cells were infected in duplicate with 250 µL of undiluted virus. Following a 1 h infection, 250 µL of fresh medium (FM) was added. Plates were incubated for 24 h. The following day, 250 µL of supernatants from the initial plate were added to fresh 6-well plates containing 250 µL of FM. The newly infected plates were incubated for 24 h and this was continued for 20 passages. At passages 5, 10, 15, and 20, all media was removed, wells washed with PBS, and cells lysed with TRIzol reagent. Intracellular RNA was extracted as previously described. For each passage, the remaining viral supernatants were collected and evaluated for infectious titers via plaque assay.
2.17. Sequencing
cDNA for the intracellular RNA extracted at serial passages 5, 10, 15, 20 was generated using a high-capacity cDNA reverse transcription kit (4368814, ThermoFisher) and a T7-tagged reverse primer targeted to the 3′ end of nsP3 or the 3′UTR containing the CSE sequence on the positive-strand of VEEV TC-83. cDNA was amplified using a T7 primer and a forward primer targeting the 5′ end of the nsP3 and CSE sequences of VEEV TC-83 using PlatinumSuperFi PCR master mix kit (12358-050, Invitrogen): 1 cycle at 98 °C for 30 s, 35 cycles at 98 °C for 5 s, 51 °C for 10 s, 72 °C for 4 min, and 1 cycle at 72 °C for 5 min. All primer sets are represented in
Table 2. Forward primers were designed every 800 base-pairs along the nsP3 and 3′UTR segment of the VEEV TC-83 genome and used to sequence viral cDNA, performed using Sanger Sequencing by Macrogen Corp. (Rockville, MD, USA). SnapGene Viewer was utilized to view chromatograms.
2.18. Bioinformatics
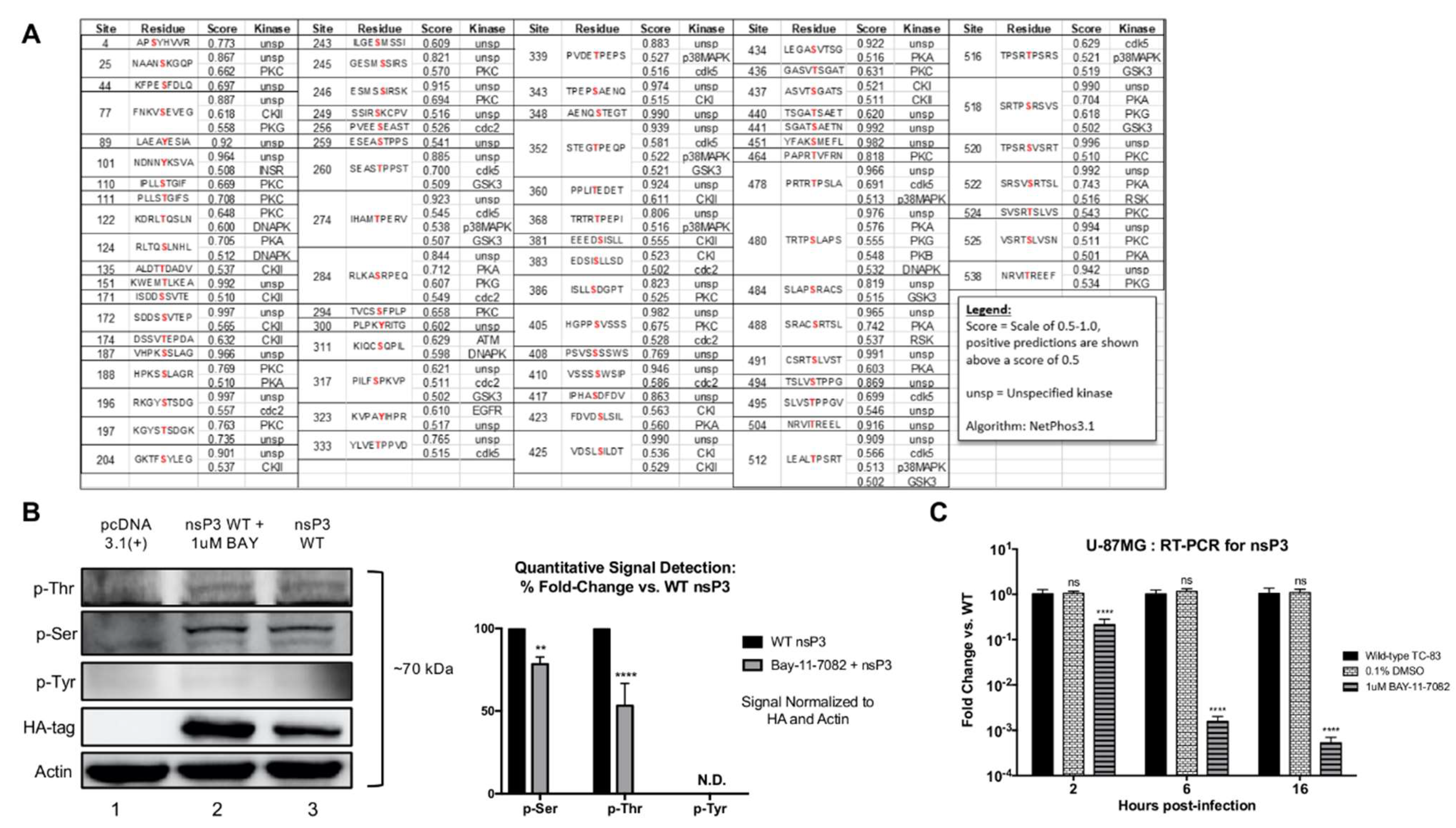
Phosphorylation site predictions on VEEV nsP3 were performed with the VEEV TC-83 sequence (L01443) and the NetPhos 3.1 Server with results filtered for hits scored above 0.5. All sequence alignments utilized Clustal Omega (EMBL-EBI). DNA to protein analysis was performed with ExPASy SIB Bioinformatics Resource Portal. The following sequences obtained from the NCBI nucleotide database were used for alignments: VEEV Enzootic strain NP_740698.1, VEEV TrD strain L01442, VEEV TC-83 strain L01443, EEEV FL93 Strain ABL84686.1, SINV TR339 Strain (William Klimstra), CHIKV LR Strain AQX78116.1.
2.19. Imaging
Undiluted viral supernatants from serial passages 11–13 for mutants 204/5 and 142 and passage 5 for mutant 134/5 were used to infect Vero cells in a 96-well plate for 24 h. Wild-type VEEV TC-83 and mock infection controls were included. At 24 hpi, media was removed, cells washed with PBS, and fixed with 4% paraformaldehyde. Cells were imaged using EVOS™ FL Auto Imaging System (ThermoFisher). Images were taken using 4X objective with brightness adjusted to 50% and photo viewer clarity set to 100. Inset images were taken using 20× objective with brightness adjusted to 70% and photo viewer clarity set to 100. Scale bars for 4× and 20× images are 1000 µm and 200 µm, respectively.
2.20. Statistical Analyses
All graphs represent the mean ± SD for all data obtained. Prism 7 (Graph Pad, San Diego, CA, USA) was used for all statistical analyses and statistical significance was determined using Two-Way ANOVA with Dunnett’s Post Test unless otherwise stated. Significance values are indicated using asterisks for * p < 0.0332, ** p < 0.0021, *** p < 0.0002, and **** p < 0.0001.
4. Discussion
4.1. The Role of IKKβ during the Alphavirus Replication Cycle
The nsP3 of both Old World and New World alphaviruses has been studied due to its capacity for hyperphosphorylation, variability among alphaviruses, and its interaction with a variety of host proteins [
9,
12]. However, cellular kinases responsible for phosphorylating various sites in nsP3 have yet to be determined [
12]. We have previously shown that inhibiting IKKβ activity leads to greatly reduced viral replication following VEEV infection [
31]. Here, we extend our previous observations to demonstrate IKKβ-mediated phosphorylation of VEEV nsP3 and elucidate the role of this post-translational modification in the viral replication cycle. First, we demonstrated by western blot that nsP3 phosphorylation levels decrease upon treatment with the IKKβ inhibitor BAY-11-7082. We also used a cell-free, in vitro kinase activity assay containing purified IKKβ and VEEV nsP3 to quantitatively identify nsP3 as a substrate targeted by IKKβ. To our knowledge, this is the first identified cellular kinase responsible for phosphorylating VEEV nsP3.
33P-ATP counts for nsP3 were significantly higher than those measured from potential IKKβ autophosphorylation and non-specific IKKβ kinase activity. The lower level of nsP3 phosphorylation vs. the positive control in this assay may be attributed to a lower input concentration of nsP3 substrate vs. the control peptide. Given that the control substrate is a small, highly purified peptide, a greater number of peptide molecules present at a given concentration when compared to the full-length nsP3 protein would also result in a higher observed signal. Furthermore, VEEV nsP3 may not have acquired all post-translational modifications when expressed in a bacterial system. Alternatively, the presence of nsP3 alone, in the absence of other non-structural proteins, may affect the proficiency of its interaction with IKKβ. Similar methodology was employed for SINV nsP3 in which kinase activity was detected following incubation of nsP3 and
33P-ATP, with the kinase speculated to be casein kinase II (CKII) [
53]. Interestingly, our phosphorylation prediction analysis also identified CKII as potentially phosphorylating two sites, 135 and 204. Using mass spectrometry, we have identified 13 serine, threonine, and tyrosine phosphorylation sites on VEEV nsP3 in two distinct cell types that appear to be phosphorylated by IKKβ. Evaluation of mutant nsP3 proteins, 204/5, 142, and 134/5, by a cell-free, in vitro kinase activity assay confirmed that these sites can be phosphorylated by IKKβ kinase activity. Additionally, nsP3 residues that are phosphorylated regardless of the presence or absence of the IKKβ inhibitor provide evidence that nsP3 can be phosphorylated by the activity of at least two host kinases. Indeed, our phosphorylation prediction analysis identified a variety of kinases potentially phosphorylating nsP3, including at the same residues, for example, CKII and IKKβ at sites 135 and 204. Given that VEEV replication is not completely abrogated upon BAY-11-7082 treatment or in IKKβ
−/− cells [
31], multiple cellular kinases probably phosphorylate nsP3 in the absence of IKKβ activity. Thus, our data identifies IKKβ as one kinase phosphorylating VEEV nsP3 at multiple sites but does not demonstrate that this interaction is absolutely essential for VEEV replication.
Our conclusions regarding the role of nsP3 phosphorylation during VEEV replication are supported by those obtained from a previous study using VEEV, in which 53 potential phosphorylation sites in the nsP3 HVD were mutated. That study revealed minimal effects on viral replication and concluded that the phosphorylation state of this domain is inconsequential for replication [
14]. Similarly, the mutation of eight phosphorylation sites in nsP3 HVD in our study also demonstrated minimal effects on viral replication. However, our study extends the analysis of nsP3 phosphorylation to the macrodomain and the AUD of nsP3 and identifies phosphorylation sites in both these domains that are critical for VEEV replication.
4.2. NsP3 Residues 204/5, 142, and 135 Are Critical for Negative-Strand Synthesis
Our research identified three phosphorylation sites in VEEV nsP3 at amino acid positions 204/5, 142, and 134/5 at which charge-to-alanine mutations dramatically diminished viral replication when introduced into the TC-83 or TrD strains of VEEV. Furthermore, the abrogation of phosphorylation-capable residues at any one of these sites reduced negative-strand synthesis during VEEV TC-83 infection, suggesting that nsP3 activity during replication could require multiple phosphorylation events. However, the collective impact of these residues may be cell-type dependent and phosphorylation at these residues may also be temporally regulated. For example, sites 142 and 134/5 were phosphorylated in astrocyte cells at 8 hpi but absent in the wild-type MEF cells at 6 hpi. This phenomenon could suggest that differential phosphorylation at certain nsP3 residues may occur at different time points post-infection in different cell types during replication. Furthermore, all three mutants were incapable of virion production following infection of mosquito cells, suggesting that these sites are required for viral growth in multiple cell types.
Our data supports research performed in a heat-resistant (HR) strain of SINV where a mutation at position 268 from alanine to valine in the N-terminal conserved region of nsP3 rendered the virus defective in negative-strand synthesis. Furthermore, double recombinant genomes allowed reactivation, suggesting that the impairment of negative-strand synthesis was due to prevention of active replication complex formation early during replication [
41]. Similarly, a glycine substitution at site 68 resulted in blockage of overall SINV nsP3 phosphorylation and was correlated with diminished negative-strand synthesis [
54]. Furthermore, two deletions in Semliki Forest virus nsP3 at positions 344 and 345 in the HVD, individually and doubly deleted, resulted in a considerably diminished rate of viral RNA synthesis, albeit with successful virion production [
15]. One potential mechanism by which nsP3 phosphorylation may support viral replication is by enabling interactions with host factors that may be involved in the viral replication cycle. For instance, Fragile X syndrome family proteins interact with VEEV nsP3 whereas Chikungunya virus (CHIKV) and SINV nsP3 exploit the G3BP family of proteins to enable formation of replication complexes [
17]. Mutation of residues 260 and 261 in CHIKV nsP3 can abrogate viral replication, and in silico modeling suggests these sites may play a role in nsP3 structural integrity and/or interaction with host cellular components [
55]. More recently, the HVD of CHIKV nsP3 was shown to contain two SH3 domain-binding motifs and point mutations in these sites not only reduced viral replication but also eliminated interaction with SH3 domain-containing host proteins CD2AP, BIN1, and SH3KBP1 [
56]. Binding of host proteins with the SH3 domains was found to cause allosteric effects in the HVD of CHIKV nsP3, which affected the interaction with G3BP family proteins [
56]. Our data parallels the mechanistic results described in these studies where mutations in nsP3 that abrogate phosphorylation result in decreased negative-strand RNA synthesis which may occur due to abrogation of interactions between nsP3 and host proteins facilitated by nsP3 phosphorylation.
4.3. The Evolutionary Importance of Phosphorylation Competence at Positions 204/5, 142, and 134/5
To further highlight the importance of replication-deficient nsP3 sites 204/5, 142, and 134/5 for successful viral replication, we serially passaged these mutants to generate revertants that restored infectivity and negative-strand synthesis. Interestingly, all three revertants restored the alanine mutations back to the original phosphorylation-capable residues present in wild-type TC-83 at these sites, which suggests that viral fitness requires the presence of phosphorylation competent amino acids at these sites. However, mutant 134/5 did not mutate amino acid site 134 back to the original threonine and instead mutated to glycine, indicating that of the pair, amino acid site 135 is probably more important for successful VEEV replication. Furthermore, our phosphorylation prediction analysis identified only site 135 as a residue targeted for phosphorylation. Intriguingly, alignment of SINV, CHIKV, and eastern equine encephalitis virus (EEEV) nsP3 sequences with VEEV nsP3 shows conservation of residues at 204/5, 142, and 135 among these alphaviruses, potentially suggesting that phosphorylation at these residues represents an evolutionarily conserved mechanism for successful replication.
Additionally, data from phosphomimetic mutants suggests differential roles for phosphorylation at individual sites in the replication cycle. Phosphomimetic mutant 204D/5D restored infectious particle production and this correlated with a restoration of negative-strand RNA synthesis indicating that phosphorylation at positions 204/5 in VEEV nsP3 is more important for negative-strand RNA synthesis than the identity of the amino acid residues. Indeed, as aspartic acid substitution at this site mimics a constantly phosphorylated residue, this suggests that constant phosphorylation of site 204/5 is potentially required for negative-strand synthesis. Additionally, treatment of 204D/5D with the IKKβ inhibitor BAY-11-7082 displayed a minor reduction in infectious particle production further suggesting that phosphorylation at this site is important for successful replication and negative strand synthesis. The lack of inhibition observed following treatment of 204D/5D infected cells with BAY-11-7082 may reflect phosphorylation of nsP3 by multiple kinases that presumably overcome the inhibitory effect of BAY-11-7082 on IKKβ activity.
In contrast to the rescue of replication observed with mutant 204D/5D, the phosphomimetic mutant 142D failed to restore infectious particle production or negative-strand RNA synthesis rendering interpretation of the role of phosphorylation at this site inconclusive. A mimic at this site can only represent a constitutively phosphorylated residue. Given this, we can only conclude that alanine substitution at this site abrogates viral replication and negative strand synthesis, that the aspartic acid substitution may have destabilized nsP3, that a residue mimicking constant phosphorylation at nsP3 site 142 does not appear to restore viral replication, and/or that the use of a phosphomimetic at this site cannot duplicate the natural function of a phosphorylated residue at this site in nsP3.
Most interestingly, phosphomimetic mutant 134D/5D infection resulted in a significant rescue of infectious particle production, but not negative-strand RNA synthesis. Treatment of 134D/5D with BAY-11-7082 demonstrated no significant reduction in infectious particle production supporting our previous conclusion that phosphorylation at this site is important for viral replication. Taken together with the alanine substitution data where detectable infectious particle production but negative-strand RNA levels comparable to mutants 204/5 and 142 are observed, this suggests that phosphorylation at nsP3 site 134/5 is important for infectious particle production but the identity of the amino acid residue at this site may be important for negative-strand synthesis. Alternatively, a threshold effect may apply such that the small observed increase in negative-strand level above a given amount during 134D/5D infection may be sufficient to increase infectious particle production. Another possibility, given the large observed increase in PFU production with a correspondingly small change in negative strand levels, is that phosphorylation at nsP3 site 134/5 results in an increase in the efficiency of viral genome packaging into virions. In a study using capsid protein mutants that altered its ability to package viral genomes, compensatory mutations in non-structural protein 2 (nsP2) were observed that corresponded with increased viral titers [
57]. Results suggested that nsP2 may be involved in RNA encapsidation or could mediate presentation of viral genomes for packaging. Therefore, phosphorylation of nsP3 at site 134/5 could mediate nsP3 functions different from its role in viral RNA synthesis.
Lastly, defining the mechanism by which negative-strand synthesis is curtailed may provide insights into the initiation and regulation of this activity during infection. It is unknown as to whether these mutations affect replication complex formation, binding to the viral genome at the positive-strand 3′ CSE promoter, or curtail the rate of initiation or elongation of negative-strand synthesis, and whether they do so by perturbing nsP3 function or abrogating interactions with host factors involved in negative-strand synthesis. Further, determining the means by which an increase in infectious particle production can occur without a significant increase in negative-strand RNA levels could describe a role for nsP3 outside of the viral replication complex. Future experiments using these mutants will be performed to further elucidate the molecular mechanisms by which nsP3 may regulate negative-strand synthesis and infectious particle production.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}