Identification of Circovirus Genome in a Chinstrap Penguin (Pygoscelis antarcticus) and Adélie Penguin (Pygoscelis adeliae) on the Antarctic Peninsula

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
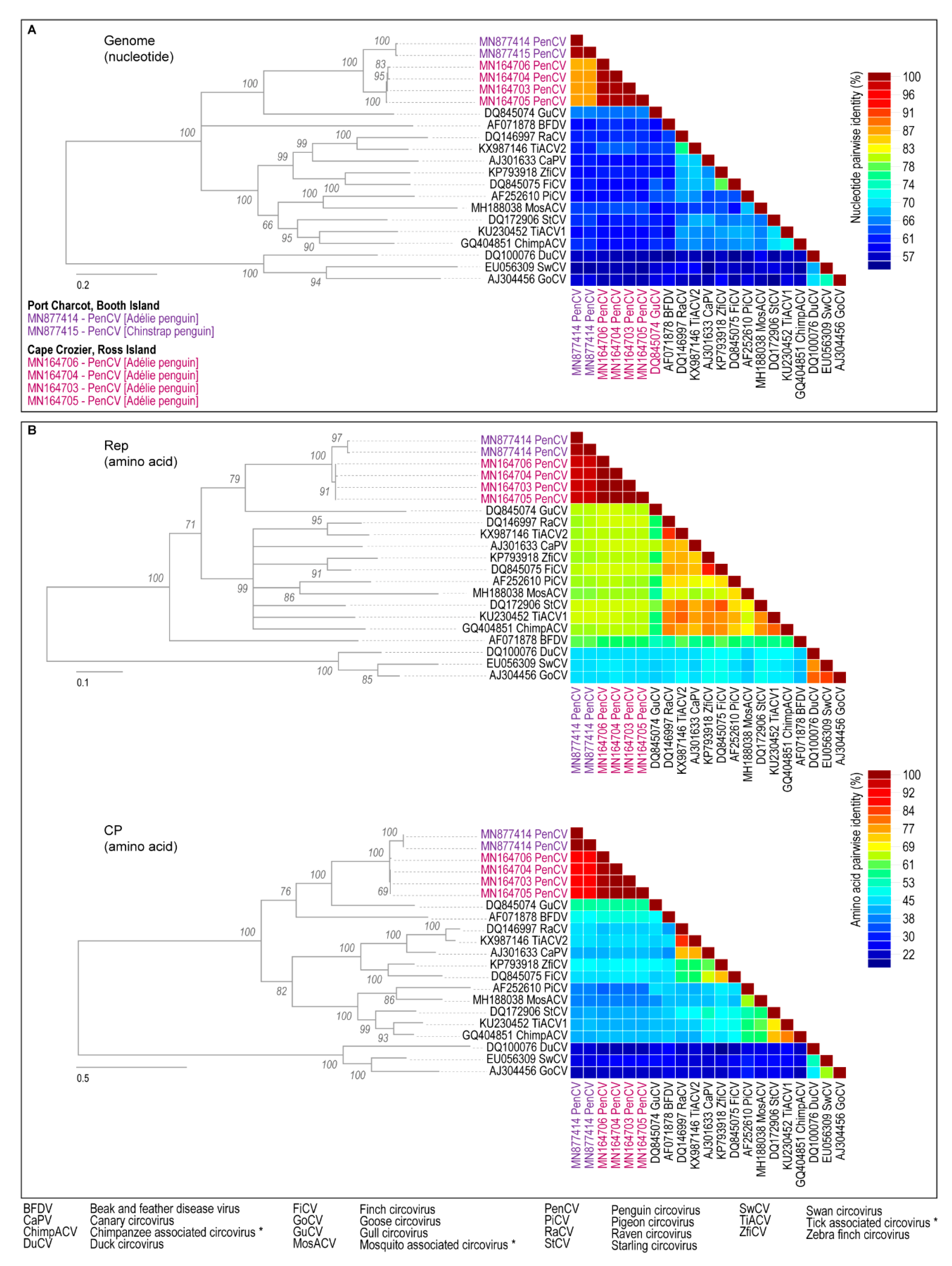
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Barbosa, A.; Palacios, M.J. Health of Antarctic birds: A review of their parasites, pathogens and diseases. Polar Biol. 2009, 32, 1095. [Google Scholar] [CrossRef]
- Grimaldi, W.W.; Seddon, P.J.; Lyver, P.O.; Nakagawa, S.; Tompkins, D.M. Infectious diseases of Antarctic penguins: Current status and future threats. Polar Biol. 2015, 38, 591–606. [Google Scholar] [CrossRef]
- Morandini, V.; Dugger, K.M.; Ballard, G.; Elrod, M.; Schmidt, A.; Ruoppolo, V.; Lescroel, A.; Jongsomjit, D.; Massaro, M.; Pennycook, J.; et al. Identification of a novel adelie penguin circovirus at cape crozier (Ross Island, Antarctica). Viruses 2019, 11, 1088. [Google Scholar] [CrossRef]
- Neira, V.; Tapia, R.; Verdugo, C.; Barriga, G.; Mor, S.; Ng, T.F.F.; Garcia, V.; Del Rio, J.; Rodrigues, P.; Briceno, C.; et al. Novel avulaviruses in penguins, Antarctica. Emerg. Infect. Dis. 2017, 23, 1212–1214. [Google Scholar] [CrossRef]
- Olivares, F.; Tapia, R.; Galvez, C.; Meza, F.; Barriga, G.P.; Borras-Chavez, R.; Mena-Vasquez, J.; Medina, R.A.; Neira, V. Novel penguin Avian avulaviruses 17, 18 and 19 are widely distributed in the Antarctic Peninsula. Transbound. Emerg. Dis. 2019, 66, 2227–2232. [Google Scholar] [CrossRef] [PubMed]
- Smeele, Z.E.; Ainley, D.G.; Varsani, A. Viruses associated with Antarctic wildlife: From serology based detection to identification of genomes using high throughput sequencing. Virus Res. 2018, 243, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Wille, M.; Aban, M.; Wang, J.; Moore, N.; Shan, S.; Marshall, J.; Gonzalez-Acuna, D.; Vijaykrishna, D.; Butler, J.; Wang, J.; et al. Antarctic penguins as reservoirs of diversity for avian avulaviruses. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Yinda, C.K.; Esefeld, J.; Peter, H.U.; Matthijnssens, J.; Zell, R. Penguin megrivirus, a novel picornavirus from an Adelie penguin (Pygoscelis adeliae). Arch. Virol. 2019, 164, 2887–2890. [Google Scholar] [CrossRef]
- Alexander, D.J.; Manvell, R.J.; Collins, M.S.; Brockman, S.J.; Westbury, H.A.; Morgan, I.; Austin, F.J. Characterization of paramyxoviruses isolated from penguins in Antarctica and sub-Antarctica during 1976-1979. Arch. Virol. 1989, 109, 135–143. [Google Scholar] [CrossRef]
- Austin, F.J.; Webster, R.G. Evidence of ortho- and paramyxoviruses in fauna from Antarctica. J. Wildl. Dis. 1993, 29, 568–571. [Google Scholar] [CrossRef]
- Baumeister, E.; Leotta, G.; Pontoriero, A.; Campos, A.; Montalti, D.; Vigo, G.; Pecoraro, M.; Savy, V. Serological evidences of influenza A virus infection in Antarctica migratory birds. Int. Congr. Ser. 2004, 1263, 737–740. [Google Scholar] [CrossRef]
- Gardner, H.; Kerry, K.; Riddle, M.; Brouwer, S.; Gleeson, L. Poultry virus infection in Antarctic penguins. Nature 1997, 387, 245. [Google Scholar] [CrossRef] [PubMed]
- Mac Donald, J.W.; Conroy, J.W.H. Virus disease resembling puffinosis in the gentoo penguin Pygoscelis papua on Signy Island South Orkney Islands. Br. Antarct. Surv. Bull. 1971, 80–82. [Google Scholar]
- Morgan, I.R.; Westbury, H.A. Virological studies of Adelie Penguins (Pygoscelis adeliae) in Antarctica. Avian Dis. 1981, 25, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Morgan, I.R.; Westbury, H.A. Studies of viruses in penguins in the Vestfold hills. Hydrobiologia 1988, 165, 263–269. [Google Scholar] [CrossRef]
- Morgan, I.R.; Westbury, H.A.; Caple, I.W.; Campbell, J. A survey of virus infection in sub-Antarctic penguins on Macquarie Island, Southern Ocean. Aust. Vet. J. 1981, 57, 333–335. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C.; Vijaykrishna, D.; Butler, J.; Baas, C.; Maurer-Stroh, S.; Silva-de-la-Fuente, M.C.; Medina-Vogel, G.; Olsen, B.; Kelso, A.; Barr, I.G.; et al. Detection of evolutionarily distinct avian influenza a viruses in Antarctica. mBio 2014, 5, e01098-14. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Kim, J.H.; Park, Y.M.; Shin, O.S.; Kim, H.; Choi, H.G.; Song, J.W. A novel adenovirus in Chinstrap penguins (Pygoscelis Antarctica) in Antarctica. Viruses 2014, 6, 2052–2061. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, J.H.; Seo, T.K.; No, J.S.; Kim, H.; Kim, W.K.; Choi, H.G.; Kang, S.H.; Song, J.W. Genetic and molecular epidemiological characterization of a novel adenovirus in Antarctic penguins collected between 2008 and 2013. PLoS ONE 2016, 11, e0157032. [Google Scholar] [CrossRef]
- Thomazelli, L.M.; Araujo, J.; Oliveira, D.B.; Sanfilippo, L.; Ferreira, C.S.; Brentano, L.; Pelizari, V.H.; Nakayama, C.; Duarte, R.; Hurtado, R.; et al. Newcastle disease virus in penguins from King George Island on the Antarctic region. Vet. Microbiol. 2010, 146, 155–160. [Google Scholar] [CrossRef]
- Varsani, A.; Kraberger, S.; Jennings, S.; Porzig, E.L.; Julian, L.; Massaro, M.; Pollard, A.; Ballard, G.; Ainley, D.G. A novel papillomavirus in Adelie penguin (Pygoscelis adeliae) faeces sampled at the Cape Crozier colony, Antarctica. J. Gen. Virol. 2014, 95, 1352–1365. [Google Scholar] [CrossRef]
- Varsani, A.; Porzig, E.L.; Jennings, S.; Kraberger, S.; Farkas, K.; Julian, L.; Massaro, M.; Ballard, G.; Ainley, D.G. Identification of an avian polyomavirus associated with Adelie penguins (Pygoscelis adeliae). J. Gen. Virol. 2015, 96, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, M.; Delwart, E.; Rosario, K.; Segales, J.; Varsani, A.; Ictv Report, C. ICTV virus taxonomy profile: Circoviridae. J. Gen. Virol. 2017, 98, 1997–1998. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.; Colominas-Ciuro, R.; Coria, N.; Centurion, M.; Sandler, R.; Negri, A.; Santos, M. First record of feather-loss disorder in Antarctic penguins. Antarct. Sci. 2015, 27, 69–70. [Google Scholar] [CrossRef]
- Gavryushkina, A.; Heath, T.A.; Ksepka, D.T.; Stadler, T.; Welch, D.; Drummond, A.J. Bayesian total-evidence dating reveals the recent crown radiation of Penguins. Syst. Biol. 2017, 66, 57–73. [Google Scholar] [CrossRef]
- Lynch, H.J.; Naveen, R.; Fagan, W.F. Censuses of penguin, blue-eyed shag Phalacrocorax atriceps and southern giant petrel Macronectes giganteus populations on the Antarctic Peninsula, 2001–2007. Mar. Ornithol. 2008, 36, 83–97. [Google Scholar]
- Humphries, G.R.W.; Naveen, R.; Schwaller, M.; Che-Castaldo, C.; McDowall, P.; Schrimpf, M.; Lynch, H.J. Mapping Application for Penguin Populations and Projected Dynamics (MAPPPD): Data and tools for dynamic management and decision support. Polar Rec. 2017, 53, 160–166. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef]
- Stover, B.C.; Muller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Yang, Z.; Wong, W.S.; Nielsen, R. Bayes empirical bayes inference of amino acid sites under positive selection. Mol. Biol. Evol. 2005, 22, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Rosario, K.; Breitbart, M.; Harrach, B.; Segales, J.; Delwart, E.; Biagini, P.; Varsani, A. Revisiting the taxonomy of the family Circoviridae: Establishment of the genus Cyclovirus and removal of the genus Gyrovirus. Arch. Virol. 2017, 162, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Naveen, R.; Forrest, S.C.; Dagit, R.G.; Blight, L.K.; Trivelpiece, W.Z.; Trivelpiece, S.G. Censuses of penguin, blue-eyed shag, and southern giant petrel populations in the Antarctic Peninsula region, 1994–2000. Polar Rec. 2000, 36, 323–334. [Google Scholar] [CrossRef]
- Julian, L.; Piasecki, T.; Chrzastek, K.; Walters, M.; Muhire, B.; Harkins, G.W.; Martin, D.P.; Varsani, A. Extensive recombination detected among beak and feather disease virus isolates from breeding facilities in Poland. J. Gen. Virol. 2013, 94, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Smyth, J.A.; Todd, D.; Scott, A.; Beckett, A.; Twentyman, C.M.; Brojer, C.; Uhlhorn, H.; Gavier-Widen, D. Identification of circovirus infection in three species of gull. Vet. Rec. 2006, 159, 212–214. [Google Scholar] [CrossRef] [PubMed]
- Cimino, M.A.; Lynch, H.J.; Saba, V.S.; Oliver, M.J. Projected asymmetric response of Adelie penguins to Antarctic climate change. Sci. Rep. 2016, 6, 28785. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Colony and Antarctic Site Inventory Subarea | Coordinates | Penguin Species (Individuals Sampled) | PenCV Dentification in Adults Sampled | PenCV Identification in Chicks Sampled |
|---|---|---|---|---|
| Half Moon Island (HMI), South Shetland Islands | 60°36′ S, 59°55′ W | Chinstrap (10) | 0/10 | - |
| Yankee Harbor (YH), Greenwich Island, South Shetland Islands | 62°32′ S, 59°47′ W | Gentoo (9) | 0/6 | 0/3 |
| Baily Head (BH), Deception Island, South Shetland Islands | 62°58′ S, 60°30′ W | Chinstrap (10) | 0/5 | 0/5 |
| Kinnes Cove (KC, Madder Cliff), Joinville Island, Northeast Antarctic Peninsula | 63°18′ S, 56°29′ W | Adélie (14) | 0/7 | 0/7 |
| Georges Point (GP), Ronge Island, Northwest Antarctic Peninsula | 64°40′ S, 62°40′ W | Chinstrap (6) | 0/3 | 0/3 |
| Gentoo (6) | 0/3 | 0/3 | ||
| Port Charcot (PC), Booth Island, Southwest Antarctic Peninsula | 65°05′ S, 64°00′ W | Adélie (3) | 1/2 | 0/1 |
| Chinstrap (6) | 1/3 | 0/3 | ||
| Gentoo (6) | 0/3 | 0/3 | ||
| Moot Point (MP), Southwest Antarctic Peninsula | 65°12′ S, 64°06′ W | Gentoo (5) | 0/4 | 0/1 |
| Total by Species | Adélie (17) | 1/9 | 0/8 | |
| Chinstrap (32) | 1/21 | 0/11 | ||
| Gentoo (26) | 0/16 | 0/10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levy, H.; Fiddaman, S.R.; Djurhuus, A.; Black, C.E.; Kraberger, S.; Smith, A.L.; Hart, T.; Varsani, A. Identification of Circovirus Genome in a Chinstrap Penguin (Pygoscelis antarcticus) and Adélie Penguin (Pygoscelis adeliae) on the Antarctic Peninsula. Viruses 2020, 12, 858. https://doi.org/10.3390/v12080858
Levy H, Fiddaman SR, Djurhuus A, Black CE, Kraberger S, Smith AL, Hart T, Varsani A. Identification of Circovirus Genome in a Chinstrap Penguin (Pygoscelis antarcticus) and Adélie Penguin (Pygoscelis adeliae) on the Antarctic Peninsula. Viruses. 2020; 12(8):858. https://doi.org/10.3390/v12080858
Chicago/Turabian StyleLevy, Hila, Steven R. Fiddaman, Anni Djurhuus, Caitlin E. Black, Simona Kraberger, Adrian L. Smith, Tom Hart, and Arvind Varsani. 2020. "Identification of Circovirus Genome in a Chinstrap Penguin (Pygoscelis antarcticus) and Adélie Penguin (Pygoscelis adeliae) on the Antarctic Peninsula" Viruses 12, no. 8: 858. https://doi.org/10.3390/v12080858
APA StyleLevy, H., Fiddaman, S. R., Djurhuus, A., Black, C. E., Kraberger, S., Smith, A. L., Hart, T., & Varsani, A. (2020). Identification of Circovirus Genome in a Chinstrap Penguin (Pygoscelis antarcticus) and Adélie Penguin (Pygoscelis adeliae) on the Antarctic Peninsula. Viruses, 12(8), 858. https://doi.org/10.3390/v12080858