In Silico Prediction of Human Leukocytes Antigen (HLA) Class II Binding Hepatitis B Virus (HBV) Peptides in Botswana

 , , , , , and
, , , , , and
Abstract
1. Background
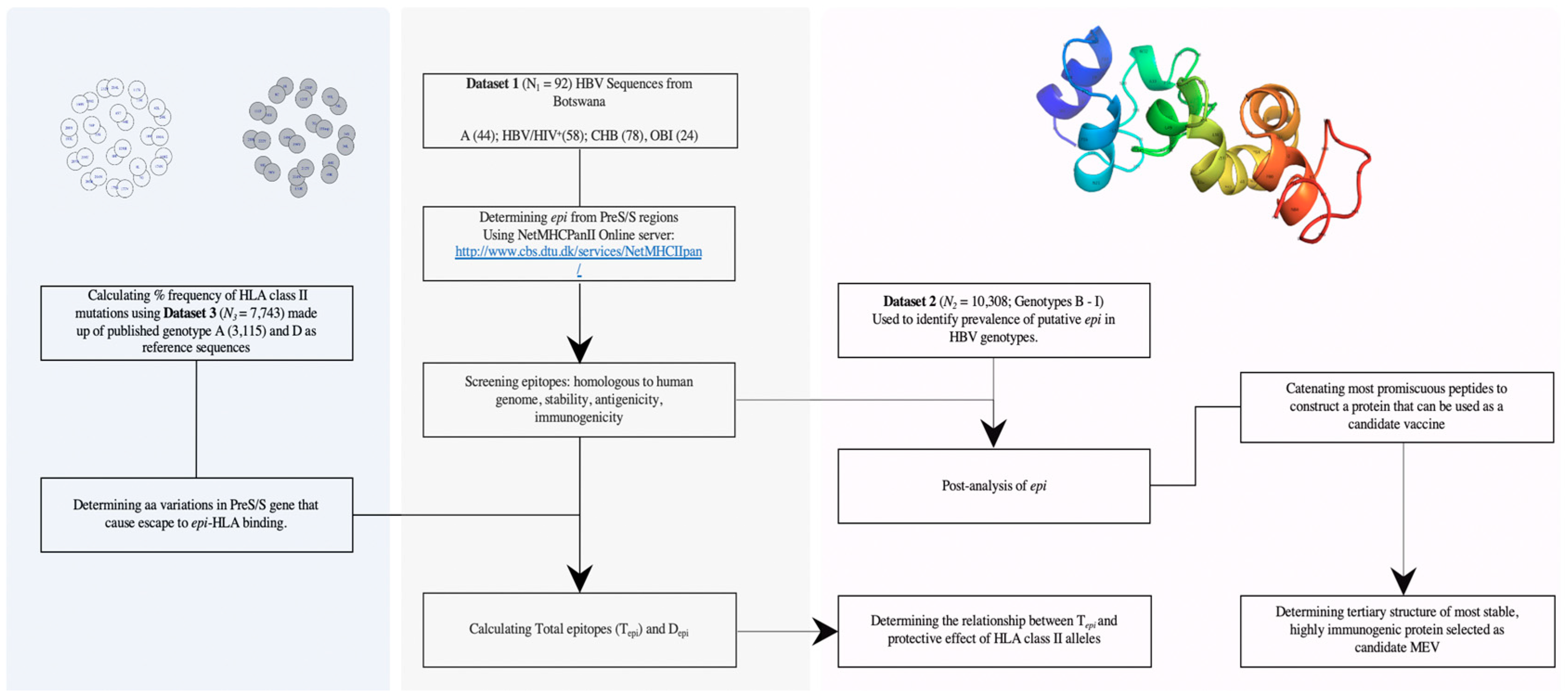
2. Materials and Methods
2.1. Mapping Peptides from HBV Surface Gene Restricted to HLA-class II Alleles
2.2. Determining Prevalence of Putative Epitopes in HBV Genotypes (A–I) Except A and D
2.3. Variations Causing Escape to HLA Class II Binding
2.4. Screening Putative Epi and Reconstruction of Tertiary Structure of the Modelled Vaccine
2.5. Determining the HLA-HBV Association Using Tepi
3. Results
3.1. Predicting T-cell Epitopes Using Consensus from N1 Dataset
3.2. Prevalence of Putative Epitopes Across all HBV Genotypes
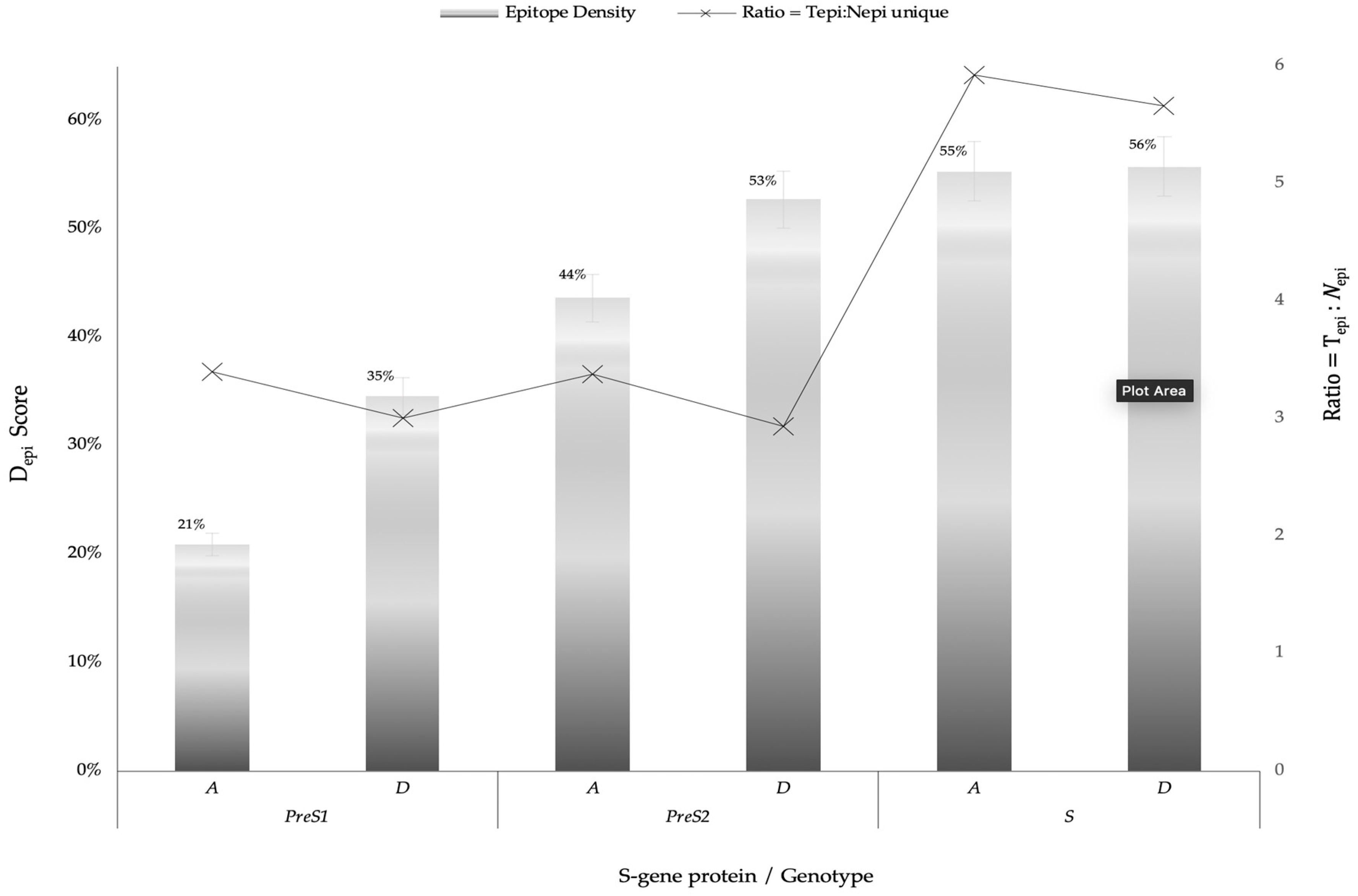
3.3. Profiles of Strong Binding Epitopes
3.4. Profiles of Most Promiscuous Epitopes
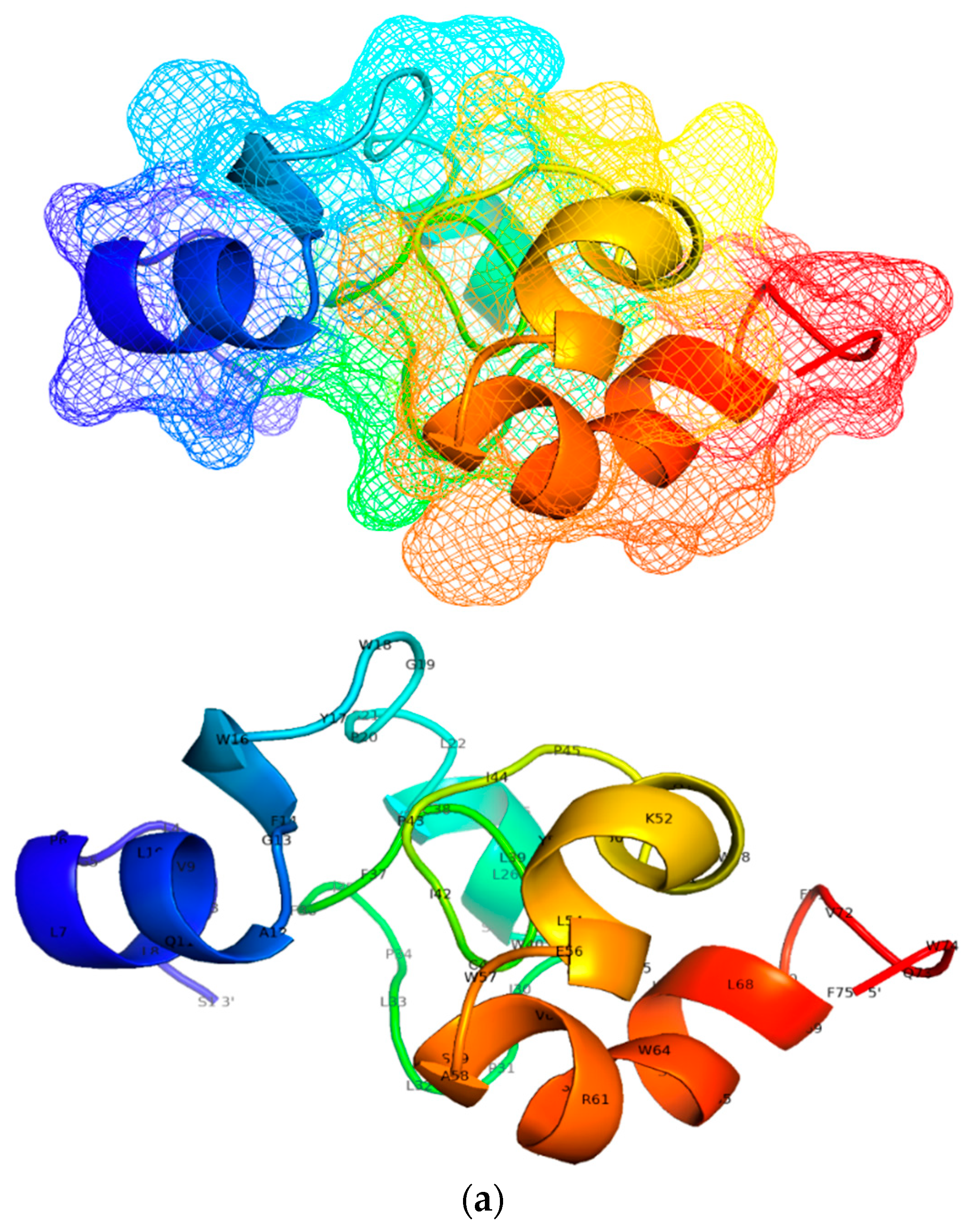
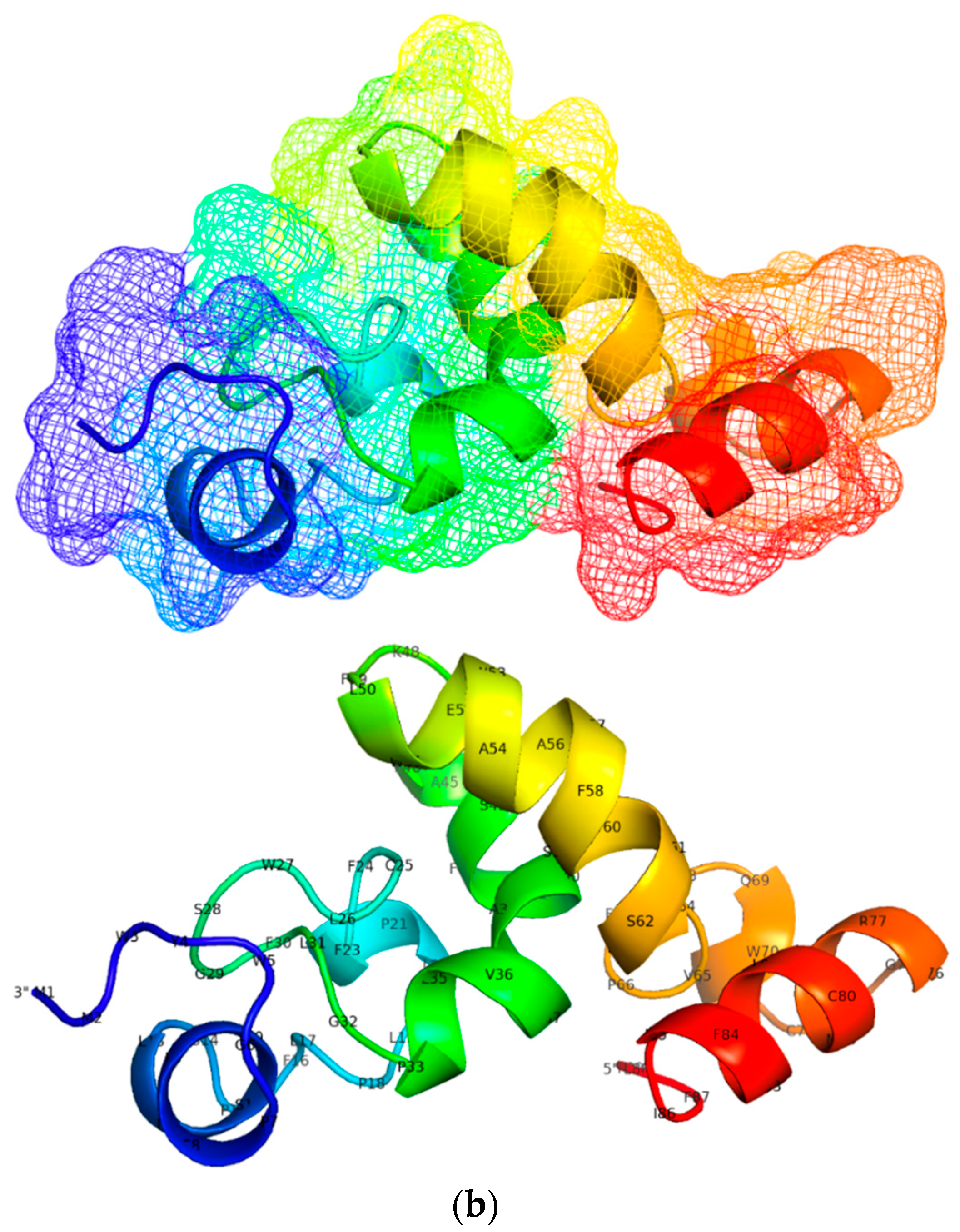
3.5. Designing and Predicting Structure of Candidate Multi-Epitope Vaccine
3.6. Mutations Associated with Escape from Class II HLA Binding
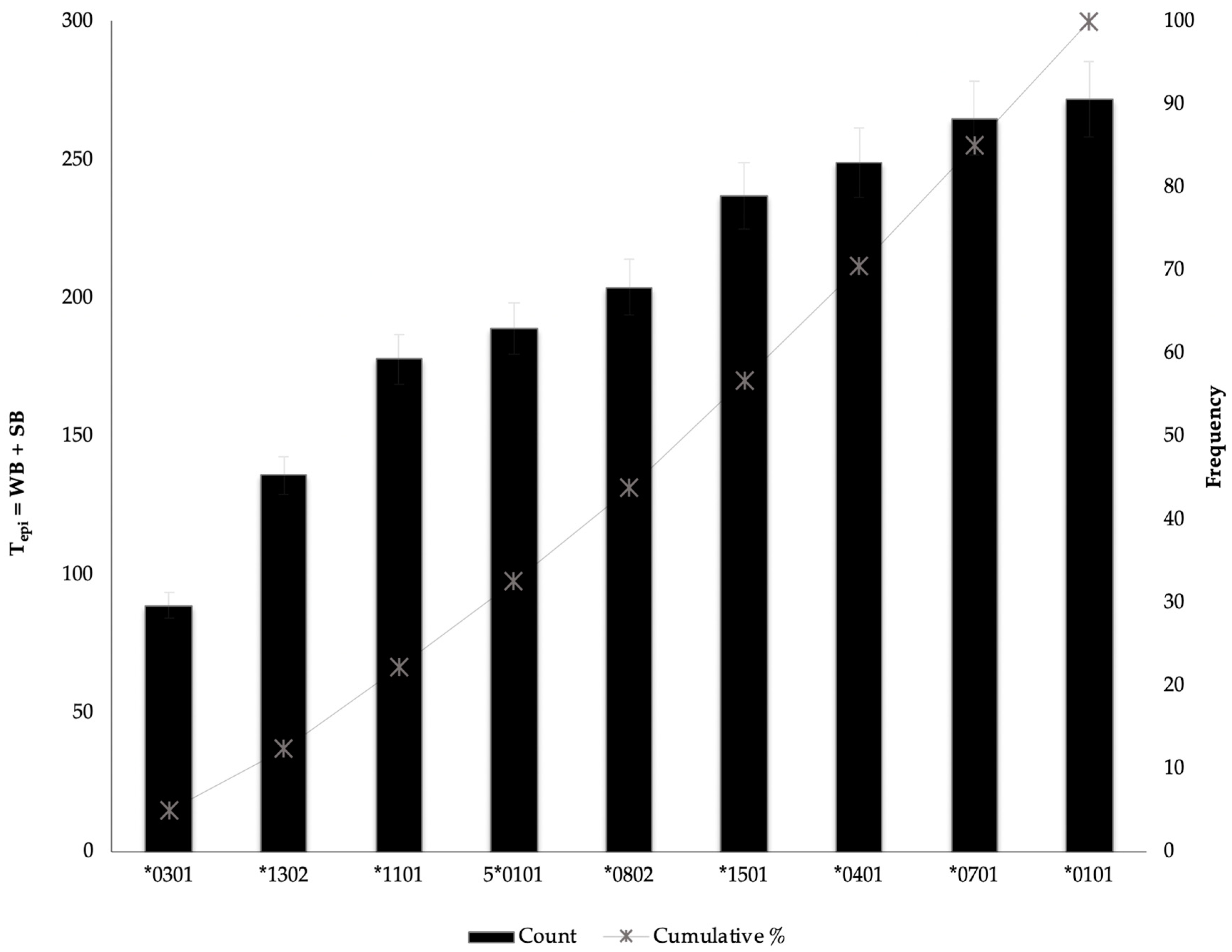
3.7. Assessing the Distribution of HBV-HLA (epi) as a Predictor of HLA Protective Effective
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HBV | hepatitis B virus |
| HLA class II | human leukocyte antigen class II |
| CHB | chronic hepatitis B infection; |
| epi | epitopes |
| SB | strong binding |
| WB | weak binding |
| MHC | major histocompatibility complex |
| aa | amino acids |
References
- Ladep, N.G.; Lesi, O.A.; Mark, P.; Lemoine, M.; Onyekwere, C.; Afihene, M.; Crossey, M.M.; Taylor-Robinson, S.D. Problem of hepatocellular carcinoma in West Africa. World J. Hepatol. 2014, 6, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Spearman, C.W.; Afihene, M.; Ally, R.; Apica, B.; Awuku, Y.; Cunha, L.; Dusheiko, G.; Gogela, N.; Kassianides, C.; Kew, M.; et al. Hepatitis B in sub-Saharan Africa: Strategies to achieve the 2030 elimination targets. Lancet Gastroenterol. Hepatol. 2017, 2, 900–909. [Google Scholar] [CrossRef]
- Kim, J.M.; Lee, K.W.; Sinn, D.H.; Choi, G.S.; Yi, N.J.; Kwon, C.H.D.; Suh, K.S.; Joh, J.W. Use of direct antiviral agents in liver transplant recipients with hepatitis C virus in Korea: 2-center experience. Ann. Surg. Treat. Res. 2018, 95, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Chisari, F.V.; Isogawa, M.; Wieland, S.F. Pathogenesis of hepatitis B virus infection. Pathol. Biol. (Paris) 2010, 58, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, W. Mechanisms of immune escape in viral hepatitis. Gut 1999, 44, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Karan, M.A.; Tascioglu, N.E.; Ozturk, A.O.; Palanduz, S.; Carin, M. The role of HLA antigens in chronic hepatitis B virus infection. J. Pak. Med. Assoc. 2002, 52, 253–256. [Google Scholar]
- Zhang, Z.H.; Wu, C.C.; Chen, X.W.; Li, X.; Li, J.; Lu, M.J. Genetic variation of hepatitis B virus and its significance for pathogenesis. World J. Gastroenterol. 2016, 22, 126–144. [Google Scholar] [CrossRef]
- Shi, Y.H. Correlation between hepatitis B virus genotypes and clinical outcomes. Jpn. J. Infect. Dis. 2012, 65, 476–482. [Google Scholar] [CrossRef]
- Thio, C.L.; Thomas, D.L.; Karacki, P.; Gao, X.; Marti, D.; Kaslow, R.A.; Goedert, J.J.; Hilgartner, M.; Strathdee, S.A.; Duggal, P.; et al. Comprehensive analysis of class I and class II HLA antigens and chronic hepatitis B virus infection. J. Virol. 2003, 77, 12083–12087. [Google Scholar] [CrossRef]
- Whitmire, J.K. Induction and function of virus-specific CD4+ T cell responses. Virology 2011, 411, 216–228. [Google Scholar] [CrossRef]
- Juno, J.A.; van Bockel, D.; Kent, S.J.; Kelleher, A.D.; Zaunders, J.J.; Munier, C.M. Cytotoxic CD4 T Cells-Friend or Foe during Viral Infection? Front. Immunol. 2017, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4(+)T cells: Differentiation and functions. Clin. Dev. Immunol. 2012, 2012, 925135. [Google Scholar] [CrossRef] [PubMed]
- Gaseitsiwe, S.; Maeurer, M.J. Identification of MHC class II binding peptides: Microarray and soluble MHC class II molecules. Methods Mol. Biol. 2009, 524, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Sette, A.; Fikes, J. Epitope-based vaccines: An update on epitope identification, vaccine design and delivery. Curr. Opin. Immunol. 2003, 15, 461–470. [Google Scholar] [CrossRef]
- Dimitrov, I.; Flower, D.R.; Doytchinova, I.A. Improving in Silico Prediction of Epitope Vaccine Candidates by Union and Intersection of Single Predictors. World J. Vaccines 2011, 1, 8. [Google Scholar] [CrossRef]
- Horvat, R.T.; El Atrouni, W.; Hammoud, K.; Hawkinson, D.; Cowden, S. Ribosomal RNA sequence analysis of Brucella infection misidentified as Ochrobactrum anthropi infection. J. Clin. Microbiol. 2011, 49, 1165–1168. [Google Scholar] [CrossRef]
- Pollicino, T.; Cacciola, I.; Saffioti, F.; Raimondo, G. Hepatitis B virus PreS/S gene variants: Pathobiology and clinical implications. J. Hepatol. 2014, 61, 408–417. [Google Scholar] [CrossRef]
- Simon, B.; Kundi, M.; Puchhammer, E. Analysis of mutations in the S gene of hepatitis B virus strains in patients with chronic infection by online bioinformatics tools. J. Clin. Microbiol 2013, 51, 163–168. [Google Scholar] [CrossRef]
- Weber, B. Diagnostic impact of the genetic variability of the hepatitis B virus surface antigen gene. J. Med. Virol. 2006, 78 (Suppl. 1), S59–S65. [Google Scholar] [CrossRef]
- Zheng, J.; Lin, X.; Wang, X.; Zheng, L.; Lan, S.; Jin, S.; Ou, Z.; Wu, J. In Silico Analysis of Epitope-Based Vaccine Candidates against Hepatitis B Virus Polymerase Protein. Viruses 2017, 9, 112. [Google Scholar] [CrossRef]
- Shi, T.D.; Wu, Y.Z.; Jia, Z.C.; Zou, L.Y.; Zhou, W. Therapeutic polypeptides based on HBV core 18-27 epitope can induce CD8+ CTL-mediated cytotoxicity in HLA-A2+ human PBMCs. World J. Gastroenterol. 2004, 10, 1902–1906. [Google Scholar] [CrossRef]
- Dimitrov, I.; Garnev, P.; Flower, D.R.; Doytchinova, I. MHC Class II Binding Prediction-A Little Help from a Friend. J. Biomed. Biotechnol. 2010, 2010, 705821. [Google Scholar] [CrossRef][Green Version]
- Rosa, D.S.; Ribeiro, S.P.; Cunha-Neto, E. CD4+ T cell epitope discovery and rational vaccine design. Arch. Immunol. Et Ther. Exp. 2010, 58, 121–130. [Google Scholar] [CrossRef]
- Schweitzer, A.; Horn, J.; Mikolajczyk, R.T.; Krause, G.; Ott, J.J. Estimations of worldwide prevalence of chronic hepatitis B virus infection: A systematic review of data published between 1965 and 2013. Lancet 2015, 386, 1546–1555. [Google Scholar] [CrossRef]
- Choga, W.T.; Anderson, M.; Zumbika, E.; Moyo, S.; Mbangiwa, T.; Phinius, B.B.; Melamu, P.; Kayembe, M.K.; Kasvosve, I.; Sebunya, T.K.; et al. Molecular characterization of hepatitis B virus in blood donors in Botswana. Virus Genes 2019, 55, 33–42. [Google Scholar] [CrossRef]
- Anderson, M.; Choga, W.T.; Moyo, S.; Bell, T.G.; Mbangiwa, T.; Phinius, B.B.; Bhebhe, L.; Sebunya, T.K.; Lockman, S.; Marlink, R.; et al. Molecular Characterization of Near Full-Length Genomes of Hepatitis B Virus Isolated from Predominantly HIV Infected Individuals in Botswana. Genes (Basel) 2018, 9, 453. [Google Scholar] [CrossRef]
- Mbangiwa, T.; Kasvosve, I.; Anderson, M.; Thami, P.K.; Choga, W.T.; Needleman, A.; Phinius, B.B.; Moyo, S.; Leteane, M.; Leidner, J.; et al. Chronic and Occult Hepatitis B Virus Infection in Pregnant Women in Botswana. Genes (Basel) 2018, 9, 259. [Google Scholar] [CrossRef]
- Anderson, M.; Gaseitsiwe, S.; Moyo, S.; Wessels, M.J.; Mohammed, T.; Sebunya, T.K.; Powell, E.A.; Makhema, J.; Blackard, J.T.; Marlink, R.; et al. Molecular characterisation of hepatitis B virus in HIV-1 subtype C infected patients in Botswana. Bmc Infect. Dis. 2015, 15, 335. [Google Scholar] [CrossRef]
- Matthews, P.C.; Beloukas, A.; Malik, A.; Carlson, J.M.; Jooste, P.; Ogwu, A.; Shapiro, R.; Riddell, L.; Chen, F.; Luzzi, G.; et al. Prevalence and Characteristics of Hepatitis B Virus (HBV) Coinfection among HIV-Positive Women in South Africa and Botswana. PLoS ONE 2015, 10, e0134037. [Google Scholar] [CrossRef]
- Wai, C.T.; Fontana, R.J. Clinical significance of hepatitis B virus genotypes, variants, and mutants. Clin. Liver Dis. 2004, 8, 321–352. [Google Scholar] [CrossRef]
- Chu, C.J.; Lok, A.S. Clinical significance of hepatitis B virus genotypes. Hepatology 2002, 35, 1274–1276. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Guo, X.R.; Wu, Y.T.; Kang, X.Z.; Zheng, Q.B.; Qi, R.Y.; Chen, B.B.; Lan, Y.; Wei, M.; Wang, S.J.; et al. A unique B cell epitope-based particulate vaccine shows effective suppression of hepatitis B surface antigen in mice. Gut 2020, 69, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Patronov, A.; Doytchinova, I. T-cell epitope vaccine design by immunoinformatics. Open Biol. 2013, 3, 120139. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Ndung’u, T.; Gaseitsiwe, S.; Sepako, E.; Doualla-Bell, F.; Peter, T.; Kim, S.; Thior, I.; Novitsky, V.A.; Essex, M. Major histocompatibility complex class II (HLA-DRB and -DQB) allele frequencies in Botswana: Association with human immunodeficiency virus type 1 infection. Clin. Diagn. Lab. Immunol. 2005, 12, 1020–1028. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jensen, K.K.; Andreatta, M.; Marcatili, P.; Buus, S.; Greenbaum, J.A.; Yan, Z.; Sette, A.; Peters, B.; Nielsen, M. Improved methods for predicting peptide binding affinity to MHC class II molecules. Immunology 2018, 154, 394–406. [Google Scholar] [CrossRef]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef]
- Bell, T.G.; Yousif, M.; Kramvis, A. Bioinformatic curation and alignment of genotyped hepatitis B virus (HBV) sequence data from the GenBank public database. Springerplus 2016, 5, 1896. [Google Scholar] [CrossRef]
- Miruka, C.O.; Matunda, N.C.; Ejekwumadu, N.J.; Mokembo, J.N. Design of a Recombinant Hepatitis B Vaccine Based on Stably Binding HLA-I Peptides. J. Biomol. Res. Ther. 2015, 4. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Racz, R.; Li, X.; Patel, M.; Xiang, Z.; He, Y. DNAVaxDB: The first web-based DNA vaccine database and its data analysis. Bmc Bioinform. 2014, 15 (Suppl. 4), S2. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Lu, J.; Li, R.; Zhao, S. Correction to: Immunogenicity of adenovirus-vector vaccine targeting hepatitis B virus: Non-clinical safety assessment in non-human primates. Virol. J. 2018, 15, 137. [Google Scholar] [CrossRef]
- Mizukoshi, E.; Sidney, J.; Livingston, B.; Ghany, M.; Hoofnagle, J.H.; Sette, A.; Rehermann, B. Cellular immune responses to the hepatitis B virus polymerase. J. Immunol. 2004, 173, 5863–5871. [Google Scholar] [CrossRef]
- Schneider, J.; Chorlton, J.; Pearce, G.; Nicola, J.; Brown, D. Compositions for Inducing an Immune Response against Hepatitis B. U.S. Patent Application No. 11/986,294, 30 October 2008. [Google Scholar]
- King, T.H.; Kemmler, C.B.; Guo, Z.; Mann, D.; Lu, Y.; Coeshott, C.; Gehring, A.J.; Bertoletti, A.; Ho, Z.Z.; Delaney, W.; et al. A whole recombinant yeast-based therapeutic vaccine elicits HBV X, S and Core specific T cells in mice and activates human T cells recognizing epitopes linked to viral clearance. PLoS ONE 2014, 9, e101904. [Google Scholar] [CrossRef]
- Zeigler, D.F.; Gage, E.; Roque, R.; Clegg, C.H. Epitope targeting with self-assembled peptide vaccines. Npj Vaccines 2019, 4, 30. [Google Scholar] [CrossRef]
- Falugi, F.; Petracca, R.; Mariani, M.; Luzzi, E.; Mancianti, S.; Carinci, V.; Melli, M.L.; Finco, O.; Wack, A.; Di Tommaso, A.; et al. Rationally designed strings of promiscuous CD4(+) T cell epitopes provide help to Haemophilus influenzae type b oligosaccharide: A model for new conjugate vaccines. Eur. J. Immunol. 2001, 31, 3816–3824. [Google Scholar] [CrossRef]
- Publicover, J.; Gaggar, A.; Jespersen, J.M.; Halac, U.; Johnson, A.J.; Goodsell, A.; Avanesyan, L.; Nishimura, S.L.; Holdorf, M.; Mansfield, K.G.; et al. An OX40/OX40L interaction directs successful immunity to hepatitis B virus. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef]
- Krebber, W.J.T.A.; Kessler, J.H.; Melief, C.J.M.; Kwappenberg, K.M.C. Synthetic Long Peptides (slp) for Therapeutic Vaccination Against Hepatitis B Virus Infection. U.S. Patent No. 10,376,576, 13 August 2019. [Google Scholar]
- Chen, L.; Zhang, Y.; Zhang, S.; Chen, Y.; Shu, X.; Lai, J.; Cao, H.; Lian, Y.; Stamataki, Z.; Huang, Y. A novel T-cell epitope in the transmembrane region of the hepatitis B virus envelope protein responds upon dendritic cell expansion. Arch. Virol. 2019, 164, 483–495. [Google Scholar] [CrossRef]
- Sette, A.; Sidney, J.; Southwood, S. Identification of Broadly Reactive DR Restricted Epitopes. U.S. Patent No. 6,413,517, 2 July 2002. [Google Scholar]
- Kent, S. Immunomodulating Compositions, Uses Therefore and Processes for Their Production. U.S. Patent No. 2007/0248584 A1, 25 October 2007. [Google Scholar]
- Godkin, A.; Davenport, M.; Hill, A.V. Molecular analysis of HLA class II associations with hepatitis B virus clearance and vaccine nonresponsiveness. Hepatology 2005, 41, 1383–1390. [Google Scholar] [CrossRef]
- Shen, K.; Shen, L.; Wang, J.; Jiang, Z.; Shen, B. Understanding Amino Acid Mutations in Hepatitis B Virus Proteins for Rational Design of Vaccines and Drugs. Adv. Protein. Chem. Struct. Biol. 2015, 99, 131–153. [Google Scholar] [CrossRef]
- Lerner, R.A.; Green, N.; Alexander, H.; Liu, F.T.; Sutcliffe, J.G.; Shinnick, T.M. Chemically synthesized peptides predicted from the nucleotide sequence of the hepatitis B virus genome elicit antibodies reactive with the native envelope protein of Dane particles. Proc. Natl. Acad. Sci. USA 1981, 78, 3403–3407. [Google Scholar] [CrossRef]
- Milich, D.R.; Peterson, D.L.; Leroux-Roels, G.G.; Lerner, R.A.; Chisari, F.V. Genetic regulation of the immune response to hepatitis B surface antigen (HBsAg). VI. T cell fine specificity. J. Immunol. 1985, 134, 4203–4211. [Google Scholar] [PubMed]
- Krchnak, V.; Mach, O.; Maly, A. Computer prediction of B-cell determinants from protein amino acid sequences based on incidence of beta turns. Methods Enzym. 1989, 178, 586–611. [Google Scholar] [CrossRef]
- Wang, H.; Luo, H.; Wan, X.; Fu, X.; Mao, Q.; Xiang, X.; Zhou, Y.; He, W.; Zhang, J.; Guo, Y.; et al. TNF-alpha/IFN-gamma profile of HBV-specific CD4 T cells is associated with liver damage and viral clearance in chronic HBV infection. J. Hepatol. 2020, 72, 45–56. [Google Scholar] [CrossRef]
- Pajot, A.; Michel, M.L.; Mancini-Bourgine, M.; Ungeheuer, M.N.; Ojcius, D.M.; Deng, Q.; Lemonnier, F.A.; Lone, Y.C. Identification of novel HLA-DR1-restricted epitopes from the hepatitis B virus envelope protein in mice expressing HLA-DR1 and vaccinated human subjects. Microbes Infect. 2006, 8, 2783–2790. [Google Scholar] [CrossRef]
- Zhou, C.; Li, C.; Gong, G.Z.; Wang, S.; Zhang, J.M.; Xu, D.Z.; Guo, L.M.; Ren, H.; Xu, M.; Xie, Q.; et al. Analysis of immunological mechanisms exerted by HBsAg-HBIG therapeutic vaccine combined with Adefovir in chronic hepatitis B patients. Hum. Vaccin. Immunother. 2017, 13, 1989–1996. [Google Scholar] [CrossRef]
- Bahrami, A.A.; Bandehpour, M.; Khalesi, B.; Kazemi, B. Computational Design and Analysis of a Poly-Epitope Fusion Protein: A New Vaccine Candidate for Hepatitis and Poliovirus. Int. J. Pept. Res. Ther. 2020, 26, 389–403. [Google Scholar] [CrossRef]
- Sette, A.; Sidney, J.; Southwood, S.; Vitiello, M.A.; Livingston, B.D.; Celis, E.; Kubo, R.T.; Grey, H.M.; Robert, W. Inducing cellular immune responses to hepatitis B virus using peptide compositions. U.S. Patent No. 2005/0063983 A1, 24 March 2005. [Google Scholar]
- Paoletti, E.; Gettig Russell, R.; Francis, J.-C.; Cox William, I.; Pincus Steven, E.; Johnson Gerard, P.; Limbach Keith, J.; Taisne Charles, D.E.; Riviere, M.; Norton Elizabeth, K.; et al. Genetically Engineered Vaccine Strain. AU 1997/010012 A, 3 January 1997. [Google Scholar]
- Shin, Y.W.; Ryoo, K.H.; Hong, K.W.; Chang, K.H.; Choi, J.S.; So, M.; Kim, P.K.; Park, J.Y.; Bong, K.T.; Kim, S.H. Human monoclonal antibody against Hepatitis B virus surface antigen (HBsAg). Antivir. Res. 2007, 75, 113–120. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhang, T.; Luo, W.; Chen, Y.; Zhang, J.; Xia, N. Polypeptides and antibodies for treating HBV infection and related diseases. U.S Patent No. 9,751,914 B2, 5 September 2017. [Google Scholar]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. Bmc Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef]
- Klein, J.; Sato, A. The HLA system. First of two parts. N. Engl. J. Med. 2000, 343, 702–709. [Google Scholar] [CrossRef]
- Xu, D.Z.; Wang, X.Y.; Shen, X.L.; Gong, G.Z.; Ren, H.; Guo, L.M.; Sun, A.M.; Xu, M.; Li, L.J.; Guo, X.H.; et al. Results of a phase III clinical trial with an HBsAg-HBIG immunogenic complex therapeutic vaccine for chronic hepatitis B patients: Experiences and findings. J. Hepatol. 2013, 59, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Kosinska, A.D.; Bauer, T.; Protzer, U. Therapeutic vaccination for chronic hepatitis B. Curr. Opin. Virol. 2017, 23, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Lobaina, Y.; Michel, M.L. Chronic hepatitis B: Immunological profile and current therapeutic vaccines in clinical trials. Vaccine 2017, 35, 2308–2314. [Google Scholar] [CrossRef]
- Boni, C.; Janssen, H.L.A.; Rossi, M.; Yoon, S.K.; Vecchi, A.; Barili, V.; Yoshida, E.M.; Trinh, H.; Rodell, T.C.; Laccabue, D.; et al. Combined GS-4774 and Tenofovir Therapy Can Improve HBV-Specific T-Cell Responses in Patients With Chronic Hepatitis. Gastroenterology 2019, 157, 227–241 e227. [Google Scholar] [CrossRef]
- Lok, A.S. Hepatitis B: 50 years after the discovery of Australia antigen. J. Viral Hepat. 2016, 23, 5–14. [Google Scholar] [CrossRef]
- Lok, A.S.; McMahon, B.J.; Brown, R.S., Jr.; Wong, J.B.; Ahmed, A.T.; Farah, W.; Almasri, J.; Alahdab, F.; Benkhadra, K.; Mouchli, M.A.; et al. Antiviral therapy for chronic hepatitis B viral infection in adults: A systematic review and meta-analysis. Hepatology 2016, 63, 284–306. [Google Scholar] [CrossRef]
- Lok, A.S.; Pan, C.Q.; Han, S.H.; Trinh, H.N.; Fessel, W.J.; Rodell, T.; Massetto, B.; Lin, L.; Gaggar, A.; Subramanian, G.M.; et al. Randomized phase II study of GS-4774 as a therapeutic vaccine in virally suppressed patients with chronic hepatitis B. J. Hepatol. 2016, 65, 509–516. [Google Scholar] [CrossRef]
- Gaggar, A.; Coeshott, C.; Apelian, D.; Rodell, T.; Armstrong, B.R.; Shen, G.; Subramanian, G.M.; McHutchison, J.G. Safety, tolerability and immunogenicity of GS-4774, a hepatitis B virus-specific therapeutic vaccine, in healthy subjects: A randomized study. Vaccine 2014, 32, 4925–4931. [Google Scholar] [CrossRef]
- Jiang, Y.G.; Wang, Y.M.; Liu, T.H.; Liu, J. Association between HLA class II gene and susceptibility or resistance to chronic hepatitis B. World J. Gastroenterol. 2003, 9, 2221–2225. [Google Scholar] [CrossRef]
- Thio, C.L.; Carrington, M.; Marti, D.; O’Brien, S.J.; Vlahov, D.; Nelson, K.E.; Astemborski, J.; Thomas, D.L. Class II HLA alleles and hepatitis B virus persistence in African Americans. J. Infect. Dis. 1999, 179, 1004–1006. [Google Scholar] [CrossRef]
- Huang, C.J.; Wu, C.F.; Lan, C.Y.; Sung, F.Y.; Lin, C.L.; Liu, C.J.; Liu, H.F.; Yu, M.W. Impact of genetic heterogeneity in polymerase of hepatitis B virus on dynamics of viral load and hepatitis B progression. PLoS ONE 2013, 8, e70169. [Google Scholar] [CrossRef]
- Ferrari, C.; Bertoletti, A.; Penna, A.; Cavalli, A.; Valli, A.; Missale, G.; Pilli, M.; Fowler, P.; Giuberti, T.; Chisari, F.V.; et al. Identification of immunodominant T cell epitopes of the hepatitis B virus nucleocapsid antigen. J. Clin. Investig. 1991, 88, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Tonetto, P.A.; Goncales, N.S.; Fais, V.C.; Vigani, A.G.; Goncales, E.S.; Feltrin, A.; Goncales, F.L., Jr. Hepatitis B virus: Molecular genotypes and HBeAg serological status among HBV-infected patients in the southeast of Brazil. Bmc Infect. Dis. 2009, 9, 149. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; Choga, W.T.; Moyo, S.; Bell, T.G.; Mbangiwa, T.; Phinius, B.B.; Bhebhe, L.; Sebunya, T.K.; Makhema, J.; Marlink, R.; et al. In Silico Analysis of Hepatitis B Virus Occult Associated Mutations in Botswana Using a Novel Algorithm. Genes (Basel) 2018, 9, 420. [Google Scholar] [CrossRef] [PubMed]
- Tamori, A.; Kawada, N. HLA class II associated with outcomes of hepatitis B and C infections. World J. Gastroenterol. 2013, 19, 5395–5401. [Google Scholar] [CrossRef]
- Thursz, M.R.; Kwiatkowski, D.; Allsopp, C.E.; Greenwood, B.M.; Thomas, H.C.; Hill, A.V. Association between an MHC class II allele and clearance of hepatitis B virus in the Gambia. N. Engl. J. Med. 1995, 332, 1065–1069. [Google Scholar] [CrossRef]
- Wang, L.; Zou, Z.Q.; Wang, K. Clinical Relevance of HLA Gene Variants in HBV Infection. J. Immunol. Res. 2016, 2016, 9069375. [Google Scholar] [CrossRef]
- Al-Qahtani, A.A.; Al-Anazi, M.R.; Abdo, A.A.; Sanai, F.M.; Al-Hamoudi, W.; Alswat, K.A.; Al-Ashgar, H.I.; Khalaf, N.Z.; Eldali, A.M.; Viswan, N.A.; et al. Association between HLA variations and chronic hepatitis B virus infection in Saudi Arabian patients. PLoS ONE 2014, 9, e80445. [Google Scholar] [CrossRef]
- Yan, Z.H.; Fan, Y.; Wang, X.H.; Mao, Q.; Deng, G.H.; Wang, Y.M. Relationship between HLA-DR gene polymorphisms and outcomes of hepatitis B viral infections: A meta-analysis. World J. Gastroenterol. 2012, 18, 3119–3128. [Google Scholar] [CrossRef]
- Li, Z.K.; Nie, J.J.; Li, J.; Zhuang, H. The effect of HLA on immunological response to hepatitis B vaccine in healthy people: A meta-analysis. Vaccine 2013, 31, 4355–4361. [Google Scholar] [CrossRef]
- Yang, P.M.; Sung, J.L.; Chen, D.S. HLA-A, B, C and DR antigens in chronic hepatitis B viral infection. Hepatogastroenterology 1989, 36, 363–366. [Google Scholar]
- Czaja, A.J.; Doherty, D.G.; Donaldson, P.T. Genetic bases of autoimmune hepatitis. Dig. Dis. Sci. 2002, 47, 2139–2150. [Google Scholar] [CrossRef]
- Parkin, J.; Cohen, B. An overview of the immune system. Lancet 2001, 357, 1777–1789. [Google Scholar] [CrossRef]
- Starzl, T.E.; Zinkernagel, R.M. Antigen localization and migration in immunity and tolerance. N. Engl. J. Med. 1998, 339, 1905–1913. [Google Scholar] [CrossRef]
- Scully, C.; Georgakopoulou, E.A.; Hassona, Y. The Immune System: Basis of so much Health and Disease: 3. Adaptive Immunity. Dent. Update 2017, 44, 322–324, 327. [Google Scholar] [CrossRef]
- Finlay, B.B.; McFadden, G. Anti-immunology: Evasion of the host immune system by bacterial and viral pathogens. Cell 2006, 124, 767–782. [Google Scholar] [CrossRef]
- Thimme, R.; Wieland, S.; Steiger, C.; Ghrayeb, J.; Reimann, K.A.; Purcell, R.H.; Chisari, F.V. CD8(+) T cells mediate viral clearance and disease pathogenesis during acute hepatitis B virus infection. J. Virol. 2003, 77, 68–76. [Google Scholar] [CrossRef]
- Rehermann, B. Intrahepatic T cells in hepatitis B: Viral control versus liver cell injury. J. Exp. Med. 2000, 191, 1263–1268. [Google Scholar] [CrossRef]
- Hass, M.; Hannoun, C.; Kalinina, T.; Sommer, G.; Manegold, C.; Gunther, S. Functional analysis of hepatitis B virus reactivating in hepatitis B surface antigen-negative individuals. Hepatology 2005, 42, 93–103. [Google Scholar] [CrossRef]
- Chang, J.J.; Lewin, S.R. Immunopathogenesis of hepatitis B virus infection. Immunol. Cell Biol. 2007, 85, 16–23. [Google Scholar] [CrossRef]
- Albayrak, A.; Ertek, M.; Tasyaran, M.A.; Pirim, I. Role of HLA allele polymorphism in chronic hepatitis B virus infection and HBV vaccine sensitivity in patients from eastern Turkey. Biochem Genet. 2011, 49, 258–269. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PreS1 | PreS2 | S | |||||
|---|---|---|---|---|---|---|---|
| A (120 aa) | D (108 aa) | A (55 aa) | D (55 aa) | A (226 aa) | D (226 aa) | ||
| Total bindings (n = 1819) SB (367); WB (1452) (%) | 6; 79 (4.7) | 7; 107 (6.3) | 4; 81 (4.7) | 3; 78 (4.5) | 122; 619 (40.7) | 136; 577 (39.1%) | |
| Unique epi (n = 297) | 25 ¥ | 38 ¥ | 29 ¥ | 24 ¥ | 125 ¥ | 126 ¥ | |
| Ratio = Tepi: Nepi | 3.4 | 3.0 | 3.4 | 2.9 | 5.9 | 5.7 | |
| Most active HLA: DRB * (SB; WB) | *0802; *0101 | *0401; (*0401, *0101) | *0301; (*0401, *0101) | *0301; 5*0101 | *0702; *0401 | *0401; *1501 | |
| Genotype variation: |  |  |  | ||||
| (A|D epi; p-value) | A|D > 0.05 | A|D > 0.05 | A|D > 0.05 | ||||
| Epitope Sites in S Protein | AA Sequence | B (n = 2905) | C (n = 5575) | E (n = 1118) | F (n = 477) | G (n = 86) | H (n = 69) | I (n = 78) | Prevalence (%) = 1 − *% | Degree of Conservation ↓ (+: Variable) | epi Previously Discussed |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 17–31 | AFGKFLWEWASARFS | E = 998 | C = 32 | F = 1 | 10,0 | + | [42] | ||||
| 180–194 | AGFFLLTRILTIPQS | B = 62 | C = 4627 | E = 5 | F = 2 | G = 75 | 46,3 | ++ | [43] | ||
| 90–104 | CLIFLLVLLDYQGML | B = 2605 | C = 4661 | E = 1034 | F = 442 | G = 84 | H = 65 | I = 70 | 86,9 | ++++ | [44] |
| 69–83 | CPGYRWMCLRRFIIF | B = 2384 | C = 5064 | E = 1027 | F = 465 | G = 84 | H = 67 | I = 65 | 88,8 | ++++ | [42] |
| 19–33 | FFLLTRILTIPQSLD | B = 63 | C = 4717 | E = 5 | F = 2 | G = 76 | 47,2 | ++ | [45,46,47] | ||
| 158–172 | FGKFLWEWASARFSW | E = 996 | C = 31 | F = 1 | 10,0 | + | [48] | ||||
| 20–34 | FLLTRILTIPQSLDS | B = 63 | C = 4709 | E = 5 | F = 5 | G = 78 | 47,1 | ++ | [49] | ||
| 93–107 | FLLVLLDYQGMLPVC | B = 2616 | C = 4720 | E = 1031 | F = 446 | G = 84 | H = 66 | I = 72 | 87,7 | ++++ | [50] |
| 161–175 | FLWEWASARFSWLSL | B = 1 | E = 997 | C = 81 | F = 3 | I = 16 | 10,7 | + | [42] | ||
| 179–193 | FVQWFVGLSPTVWLS | B = 2630 | C = 4698 | E = 76 | G = 85 | I = 75 | 73,4 | +++ | [51] | ||
| 18–32 | GFFLLTRILTIPQSL | B = 63 | C = 4618 | E = 5 | F = 2 | G = 76 | 46,2 | ++ | - | ||
| 159–173 | GKFLWEWASARFSWL | E = 1002 | C = 31 | F = 3 | 10,1 | + | - | ||||
| 202–216 | GPSLYSILSPFLPLL | C = 19 | E = 2 | 0,2 | + | [48] | |||||
| 71–85 | GYRWMCLRRFIIFLF | B = 82 | C = 5065 | E = 1032 | F = 464 | G = 84 | H = 67 | I = 65 | 66,5 | +++ | - |
| 92–106 | IFLLVLLDYQGMLPV | B = 2606 | C = 4659 | E = 1030 | F = 443 | G = 84 | H = 66 | I = 70 | 86,9 | ++++ | - |
| 195–209 | IWMMWYWGPSLYSIL | B = 3 | C = 24 | E = 2 | 0,3 | + | - | ||||
| 160–174 | KFLWEWASARFSWLS | B = 1 | E = 1012 | C = 34 | F = 3 | I = 16 | 10,3 | + | - | ||
| 91–105 | LIFLLVLLDYQGMLP | B = 2611 | C = 4656 | E = 1031 | F = 443 | G = 84 | H = 66 | I = 70 | 86,9 | ++++ | [52] |
| 21–35 | LLTRILTIPQSLDSW | B = 63 | C = 4754 | E = 5 | F = 5 | G = 77 | 47,6 | ++ | [53] | ||
| 94–108 | LLVLLDYQGMLPVCP | B = 2653 | C = 4729 | E = 1032 | F = 445 | G = 84 | H = 66 | I = 72 | 88,1 | ++++ | [54] |
| 15–29 | LQAGFFLLTRILTIP | B = 62 | C = 4735 | E = 5 | F = 2 | G = 76 | 47,3 | ++ | [54] | ||
| 192–206 | LSVIWMMWYWGPSLY | B = 772 | C = 4249 | E = 889 | G = 1 | I = 50 | 57,8 | ++ | [44] | ||
| 95–109 | LVLLDYQGMLPVCPL | B = 2619 | C = 4708 | E = 1022 | F = 444 | G = 84 | H = 66 | I = 73 | 87,5 | ++++ | [55,56] |
| 162–176 | LWEWASARFSWLSLL | B = 7 | E = 1001 | C = 88 | F = 445 | G = 2 | H = 65 | I = 75 | 16,3 | + | [54] |
| 205–219 | LYSILSPFLPLLPIF | C = 21 | E = 2 | 0,2 | + | - | |||||
| 178–192 | PFVQWFVGLSPTVWL | B = 2649 | C = 4723 | E = 76 | G = 85 | I = 74 | 73,8 | +++ | - | ||
| 70–84 | PGYRWMCLRRFIIFL | B = 2375 | C = 5117 | E = 1033 | F = 466 | G = 85 | H = 67 | I = 65 | 89,3 | ++++ | [49] |
| 66–80 | PPTCPGYRWMCLRRF | B = 60 | C = 751 | E = 14 | F = 464 | G = 77 | H = 68 | 13,9 | + | [44] | |
| 203–217 | PSLYSILSPFLPLLP | C = 19 | E = 2 | 0,2 | + | - | |||||
| 67–81 | PTCPGYRWMCLRRFI | B = 60 | C = 750 | E = 14 | F = 464 | G = 77 | H = 68 | 13,9 | + | [57] | |
| 16–30 | QAGFFLLTRILTIPQ | B = 62 | C = 4645 | E = 5 | F = 2 | G = 76 | 46,5 | ++ | [58] | ||
| 181–195 | QWFVGLSPTVWLSVI | B = 2588 | C = 4310 | E = 73 | G = 6 | I = 71 | 68,4 | +++ | [58] | ||
| 38–52 | SLNFLGGTTVCLGQN | B = 5 | C = 2 | E = 2 | 0,1 | + | [55,56,57] | ||||
| 204–218 | SLYSILSPFLPLLPI | C = 19 | E = 2 | 0,2 | + | [44] | |||||
| 193–207 | SVIWMMWYWGPSLYS | B = 3 | C = 24 | E = 2 | 0,3 | + | - | ||||
| 155–169 | SWAFGKFLWEWASAR | E = 998 | C = 31 | F = 1 | 10,0 | + | - | ||||
| 68–82 | TCPGYRWMCLRRFII | B = 63 | C = 757 | E = 17 | F = 466 | G = 77 | H = 68 | 14,0 | + | - | |
| 37–51 | TSLNFLGGTTVCLGQ | B = 5 | C = 2 | E = 2 | 0,1 | + | [59,60] | ||||
| 194–208 | VIWMMWYWGPSLYSI | B = 3 | C = 24 | E = 2 | 0,3 | + | - | ||||
| 14–28 | VLQAGFFLLTRILTI | B = 61 | C = 4676 | E = 5 | F = 2 | G = 76 | 46,8 | ++ | [61]* | ||
| 180–194 | VQWFVGLSPTVWLSV | B = 2636 | C = 4584 | E = 77 | G = 6 | I = 75 | 71,6 | +++ | [62] | ||
| 156–170 | WAFGKFLWEWASARF | E = 1000 | C = 32 | F = 1 | 10,0 | + | [44] | ||||
| 163–177 | WEWASARFSWLSLLV | B = 7 | E = 1002 | C = 87 | F = 445 | G = 2 | H = 64 | I = 75 | 16,3 | + | - |
| 182–196 | WFVGLSPTVWLSVIW | B = 2506 | C = 4125 | E = 73 | G = 6 | I = 71 | 65,8 | +++ | [63] | ||
| 201–215 | WGPSLYSILSPFLPL | C = 19 | E = 2 | 0,2 | + | [52] | |||||
| 196–210 | WMMWYWGPSLYSILS | B = 3 | C = 25 | E = 2 | 0,3 | + | - | ||||
| 36–50 | WTSLNFLGGTTVCLG | B = 5 | C = 2 | E = 2 | 0,1 | + | - | ||||
| 35–49 | WWTSLNFLGGTTVCL | B = 5 | C = 2 | E = 2 | 0,1 | + | - | ||||
| 199–213 | WYWGPSLYSILSPFL | C = 18 | E = 2 | 0,2 | + | - | |||||
| 206–220 | YSILSPFLPLLPIFF | C = 19 | E = 2 | 0,2 | + | - | |||||
| 200–214 | YWGPSLYSILSPFLP | C = 18 | E = 2 | 0,2 | + | - |
| S: Residues | AA Sequence | Geno | Core aa | HLA Class II Alleles | S: Residues | AA Sequence | Geno | Core aa | HLA Class II Alleles | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 69–83 | CPGYRWMCLRRFIIF | A | YRWMCLRRF | *0101 | *0301 | *0401 | *0701 | *0802 | *1302 | 68–82 | TCPGYRWMCLRRFII | D | YRWMCLRRF | *0101 | *0301 | *0401 | *0701 | *0802 | *1302 | *1501 | ||||
| CPGYRWMCLRRFIIF | D | 168–182 | VRFSWLSLLVPFVQW | A | WLSLLVPFV | *0101 | *0401 | *0701 | *0802 | *1101 | *1302 | *1501 | 5*0101 | |||||||||||
| 68–82 | ICPGYRWMCLRRFII | A | 14–28 | VLQAGFFLLTRILTI | A | FLLTRILTI | *0301 | *0401 | *0802 | *1302 | *1501 | 5*0101 | ||||||||||||
| 70–84 | PGYRWMCLRRFIIFL | D | VLQAGFFLLTRILTI | D | ||||||||||||||||||||
| PGYRWMCLRRFIIFL | A | *0101 | *0301 | *1501 | 165–179 | WASARFSWLSLLVPF | D | FSWLSLLVP | *0101 | *0401 | *0802 | *1101 | 5*0101 | |||||||||||
| 71–85 | GYRWMCLRRFIIFLF | A | 174–188 | SLLVPFVQWFVGLSP | A | FVQWFVGLS | *0101 | *0802 | *1101 | *1501 | ||||||||||||||
| GYRWMCLRRFIIFLF | D | SLLVPFVQWFVGLSP | D | |||||||||||||||||||||
| 67–85 | PICPGYRWMCLRRFI | A | 6–20 | SGFLGPLLVLQAGFF | A | LGPLLVLQA | *0101 | *0701 | *0802 | *1101 | *1501 | 5*0101 | ||||||||||||
| 66–84 | PPICPGYRWMCLRRF | A | *0101 | *1501 | 5*0101 | SGFLGPLLVLQAGFF | D | *0701 | ||||||||||||||||
| 67–81 | PTCPGYRWMCLRRFI | D | *1501 | 7–21 | GFLGPLLVLQAGFFL | D | ||||||||||||||||||
| 147–161 | CTCIPIPSSWAFAKY | A | CIPIPSSWA | *0101 | *0401 | *0701 | *0802 | *1501 | GFLGPLLVLQAGFFL | A | ||||||||||||||
| CTCIPIPSSWAFGKF | D | 5–19 | TSGFLGPLLVLQAGF | D | ||||||||||||||||||||
| 145–159 | GNCTCIPIPSSWAFA | A | TSGFLGPLLVLQAGF | A | ||||||||||||||||||||
| 146–160 | NCTCIPIPSSWAFAK | A | 93–107 | FLLVLLDYQGMLPVC | D | LVLLDYQGM | *0101 | *0401 | *0802 | *1101 | *1501 | |||||||||||||
| NCTCIPIPSSWAFGK | D | 5*0101 | FLLVLLDYQGMLPVC | A | *0802 | *1101 | ||||||||||||||||||
| 177–191 | VPFVQWFVGLSPTVW | D | WFVGLSPTV | *0101 | *0401 | *0701 | *0802 | *1101 | 5*0101 | 92–106 | IFLLVLLDYQGMLPV | A | *0802 | *1101 | ||||||||||
| VPFVQWFVGLSPTVW | A | IFLLVLLDYQGMLPV | D | *0802 | *1101 | |||||||||||||||||||
| 168–182 | ARFSWLSLLVPFVQW | D | WLSLLVPFV | *0101 | *0401 | *0701 | *0802 | *1101 | *1501 | 5*0101 | 182–196 | WFVGLSPTVWLSVIW | D | VGLSPTVWL | *0301 | *0401 | *0802 | *1302 | *1501 | |||||
| 169–183 | RFSWLSLLVPFVQWF | D | WFVGLSPTVWLSVIW | A | *0301 | *0401 | *0802 | *1302 | ||||||||||||||||
| RFSWLSLLVPFVQWF | A | 97–111 | LLDYQGMLPVCPLIP | A | YQGMLPVCP | *0101 | *0401 | *0701 | *1101 | *1501 | 5*0101 | |||||||||||||
| 167–181 | SARFSWLSLLVPFVQ | D | LLDYQGMLPVCPLIP | D | *0401 | *0701 | *1101 | |||||||||||||||||
| 176–189 | LVPFVQWFVGLSPTV | A | WFVGLSPTV | *0101 | *0401 | *0701 | *0802 | 5*0101 | 96–110 | VLLDYQGMLPVCPLI | A | *0401 | *0701 | *1101 | ||||||||||
| LVPFVQWFVGLSPTV | D | VLLDYQGMLPVCPLI | D | *0401 | *0701 | *1101 | ||||||||||||||||||
| 166–180 | ASARFSWLSLLVPFV | D | *0101 | *0401 | *0701 | *0802 | 5*0101 | 72–86 | YRWMCLRRFIIFLFI | A | WMCLRRFII | *0701 | *0802 | *1101 | *1501 | 5*0101 | ||||||||
| 170–184 | FSWLSLLVPFVQWFV | A | *1101 | YRWMCLRRFIIFLFI | D | WMCLRRFII | ||||||||||||||||||
| FSWLSLLVPFVQWFV | D | 194–208 | VIWMMWYWGPSLYNI | A | WYWGPSLYN | *0101 | *0401 | *0701 | *0802 | *1101 | 5*0101 | |||||||||||||
| 167–181 | SVRFSWLSLLVPFVQ | A | 5*0101 | |||||||||||||||||||||
| Epitope Site | AA Sequence | HBV Protein | Core AA | HLA Class II Alleles | Previously Discussed epi |
|---|---|---|---|---|---|
| 34–48 | PVPNIASHISSISSR | PreS2A | IASHISSIS | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1302, 1*1501, 5*0101 | - |
| 39–53 | ASHISSISSRTGDPA | PreS2A | ISSISSRTG | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 38–52 | IASHISSISSRTGDP | PreS2A | ISSISSRTG | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 37–51 | TTASPLSSIFSRIGD | PreS2D | LSSIFSRIG | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 38–52 | TASPLSSIFSRIGDP | PreS2D | LSSIFSRIG | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 39–53 | ASPLSSIFSRIGDPA | PreS2D | LSSIFSRIG | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 208–222 | ILSPFIPLLPIFFCL | SA | FIPLLPIFF | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 209–223 | LSPFIPLLPIFFCLW | SA | FIPLLPIFF | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 207–221 | NILSPFIPLLPIFFC | SA | FIPLLPIFF | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1302, 5*0101 | - |
| 149–163 | CIPIPSSWAFAKYLW | SA | IPSSWAFAK | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 6–20 | SGFLGPLLVLQAGFF | SA | LGPLLVLQA | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 200–214 | YWGPSLYNILSPFIP | SA | LYNILSPFI | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1302, 1*1501 | - |
| 199–213 | WYWGPSLYNILSPFI | SA | LYNILSPFI | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 164–178 | EWASVRFSWLSLLVP | SA | VRFSWLSLL | 1*0101, 1*0401, 1*0802, 1*1101, 1*1302, 1*1501, 5*0101 | [64] |
| 168–182 | VRFSWLSLLVPFVQW | SA | WLSLLVPFV | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1302, 1*1501, 5*0101 | [44] |
| 169–183 | RFSWLSLLVPFVQWF | SA | WLSLLVPFV | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | [65] |
| 156–170 | WAFAKYLWEWASVRF | SA | YLWEWASVR | 1*0101, 1*0301, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | [49] |
| 208–222 | ILSPFLPLLPIFFCL | SD | FLPLLPIFF | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 209–223 | LSPFLPLLPIFFCLW | SD | FLPLLPIFF | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 155–169 | SWAFGKFLWEWASAR | SD | FLWEWASAR | 1*0101, 1*0301, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501 | - |
| 6–20 | SGFLGPLLVLQAGFF | SD | LGPLLVLQA | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | [44] |
| 199–212 | WYWGPSLYSILSPFL | SD | LYSILSPFL | 1*0101, 1*0401, 1*0802, 1*1101, 1*1302, 1*1501, 5*0101 | - |
| 167–181 | SARFSWLSLLVPFVQ | SD | WLSLLVPFV | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 168–182 | ARFSWLSLLVPFVQW | SD | WLSLLVPFV | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | [44] |
| 169–183 | RFSWLSLLVPFVQWF | SD | WLSLLVPFV | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | [42,49] |
| 197–211 | MMWYWGPSLYSILSP | SD | WYWGPSLYS | 1*0101, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| 68–82 | TCPGYRWMCLRRFII | SD | YRWMCLRRF | 1*0101, 1*0301, 1*0401, 1*0701, 1*0802, 1*1101, 1*1501, 5*0101 | - |
| Protein | AA Sequence | S Protein AA Residues (epi) | Escape Mutations | Count in A (n = 3115) | Count in D (n = 4628) | Count in Other Genotypes |
|---|---|---|---|---|---|---|
| SA | MENITSGFLGPQLV | 1–14 | L12Q | 13 | 1 | 3 |
| 1–14 | I4T | 13 | 25 | 91∆ | ||
| 1–14 | I4Stop | 10 | - | - | ||
| SA | GPLLVLQAGFFLLTR | 10–24 | L15Stop | 2 | - | 17 |
| SA | LNFPGGSPVCLGQNS | 39–53 | L42P | 13 | 6 | 53 |
| SA | TRILTIPQ *LDSWWT | 23 -37 | S31Stop | 6 | 12 | - |
| SA | DLWWTSLNFLGDPPV | 33–47 | G44D | 4 | - | 16 |
| DSWWTSLNFLGESPV | G44E | 105 | 87 | 826 | ||
| IPIPSSWGFAKYLWE | 150–164 | A157G | 10 | 5 | 3 | |
| SA | IPIPSSWAFVKYLWE | A159V | 20 | 3 | 234 | |
| SA | PPICPGYRWMCQRRF | 66–80 | L77R | 7 | 2 | 43 |
| L77Q | 6 | - | 18 | |||
| SA | PPICPGYRWMCLR *F | R79Stop | 9 | 10 | 20 | |
| R79H | 13 | 58 | 49 | |||
| SA | LIFLLVLLDYQDMLP | 91–105 | G102D | 1 | 2 | 3 |
| SA | CLIFLLVLLDYQGML | 90–104 | D99Stop | 15 | 5 | 8 |
| Y100C | 196 | 12 | 17 | |||
| M103I | 31 | 31 | 29 | |||
| SA | SLLVPFVQWFVGLTP LLVPFVQWFEGLSPT | 174–188 | S187T | 1 | - | - |
| G185E | 6 | 15 | 19 | |||
| V184E | 1 | 2 | 2 | |||
| L186P | 10 | - | 5 | |||
| SA | SPTVWLLAIWMMWYW | 187–193 | S193L | 104 | 257 | 612 |
| A194V | 494 | ∆ * | ||||
| SA | GPSLYNISSPFIPLL | 202–216 | L209S | 4 | 9 | 12 |
| SD | ENITSGCLGPLLVLQAGF | 2–16 | F8C | - | 1 | - |
| SA | YLWEWASVRFSWPSL | 161–175 | L173P | 1 | 3 | 11 |
| SA | SPFIPLLLIFFCLWV | 210–224 | P214L | 11 | 41 | 23 |
| SD | SSWAFGKFLWEWASA | 154–168 208–222 | K160N | 1 | 7 | 9 |
| SD | ILSPYILLLPIFFCI | F212Y + L213I + P214L | -; -; 11 | 40; 202; 41 | 3; ∆ *;23 | |
| NITSGFLGLLLVLQA | 4 -18 | P11L | 2 | 4 | 3 | |
| SD | MENITSGFLGPLLVL | 1–15 | T5P; N3S + I4T + T5A | 2; - | 4; 2 | 4; -29 |
| SD | SWWTSLNFLGETTVC | 34–48 | G44E | 105 | 87 | 826 |
| SD | SWWTSLNFRGGTTVC | L42R | 14 | 8 | 14 | |
| SD | LSVIWMMWYWGPNLY | 192–206 | S204N | 136 | 194 | 848 |
| List of aa Variations within Genotype A and D epi | M1E, M1L, M1K, M1I, M1V, M1T, M1R, E2H, E2Q, E2*, E2V, E2K, E2A, E2G, E2D, N3H, N3E, N3P, N3A, N3C, N3D, N3I, N3R, N3Y, N3K, N3T, N3G, N3S, I4L, I4R, I4Y, I4Q, I4F, I4M, I4H, I4K, I4A, I4P, I4S, I4N, I4V, I4T, T5Y, T5Q, T5L, T5R, T5E, T5K, T5V, T5P, T5I, T5S, T5A, S6G, S6F, S6A, S6P, S6T, S6L, G7C, G7D, G7*, G7A, G7V, G7K, G7E, G7R, F8C, F8A, F8G, F8V, F8Y, F8I, F8H, F8P, F8S, F8L, L9R, L9K, L9V, L9I, L9H, L9Q, L9P, G10D, G10*, G10V, G10Q, G10T, G10A, G10K, G10E, G10R, P11A, P11T, P11H, P11L, L12V, L12E, L12M, L12R, L12P, L12Q, L13Q, L13V, L13I, L13F, L13R, L13H, L13P, V14R, V14L, V14E, V14I, V14M, V14G, V14A, L15E, L15K, L15T, L15*, L15I, L15F, L15V, L15S, Q16K, Q16E, Q16H, Q16L, Q16R, Q16P, A17R, A17T, A17S, A17P, A17V, A17G, A17E, G18W, G18K, G18E, G18A, G18R, G18V, F19I, F19V, F19L, F19S, F19Y, F19C, F20I, F20Y, F20L, F20S, L21V, L21M, L21F, L21*, L21W, L21S, L22Q, L22M, L22V, L22F, L22S, L22*, L22W, T23F, T23R, T23Q, T23S, T23P, T23A, T23I, R24N, R24T, R24Q, R24E, R24I, R24G, R24S, R24K, I25R, I25S, I25F, I25N, I25A, I25T, I25V, L26Y, L26I, L26F, L26Q, L26P, L26H, L26R, T27R, T27P, T27S, T27A, T27K, T27I, I28K, I28L, I28V, I28T, I28M, P29E, P29A, P29F, P29S, P29Q, P29T, P29L, Q30M, Q30S, Q30P, Q30A, Q30L, Q30H, Q30R, Q30K, S31K, S31I, S31D, S31T, S31C, S31G, S31R, S31N, L32V, L32G, L32Q, L32R, L32I, L32P, D33V, D33H, D33Y, D33E, D33N, D33G, S34*, S34A, S34T, S34W, S34P, S34L, W35P, W35C, W35*, W35G, W35R, W35L, W36E, W36G, W36R, W36*, W36L, T37D, T37P, T37L, T37I, T37S, T37N, T37A, S38E, S38A, S38Y, S38F, S38P, L39I, L39V, L39R, L39H, L39F, L39P, N40Q, N40H, N40K, N40I, N40D, N40S, F41Q, F41I, F41Y, F41C, F41L, F41S, L42*, L42S, L42I, L42V, L42Q, L42R, L42P, G43V, G43W, G43K, G43R, G43E, G44Q, G44K, G44R, G44A, G44D, G44V, G44E, S45M, S45D, S45G, S45E, S45H, S45Q, S45R, S45I, S45K, S45N, S45V, S45L, S45P, S45A, S45T, P46N, P46R, P46S, P46I, P46A, P46H, P46L, P46T, V47Q, V47N, V47L, V47P, V47M, V47E, V47R, V47K, V47A, V47G, V47T, C48W, C48L, C48F, C48R, C48G, C48S, C48Y, L49A, L49T, L49C, L49S, L49I, L49V, L49F, L49H, L49R, L49P, G50C, G50W, G50P, G50R, G50D, G50V, G50S, G50A, Q51E, Q51K, Q51H, Q51P, Q51R, Q51L, N52T, N52G, N52H, N52K, N52I, N52D, N52S, S53G, S53M, S53*, S53T, S53P, S53W, S53L, Q54K, Q54*, Q54H, Q54L, Q54P, Q54R, S55T, S55Y, S55A, S55P, S55F, S55C, P56A, P56*, P56S, P56R, P56H, P56L, P56Q, T57S, T57A, T57I, S58Y, S58A, S58L, S58T, S58P, S58F, S58C, N59L, N59I, N59R, N59H, N59D, N59K, N59S, H60D, H60S, H60N, H60Y, H60Q, H60R, H60P, S61T, S61P, S61*, S61L, P62S, P62Q, P62L, T63V, T63F, T63N, T63S, T63A, T63I, S64W, S64P, S64F, S64Y, S64C, C65G, C65F, C65S, C65Y, C65R, P66S, P66L, P66T, P66A, P66Q, P66H, P67T, P67A, P67R, P67S, P67L, P67Q, I68D, I68P, I68V, I68F, I68S, I68N, I68A, I68T, C69F, C69G, C69S, C69R, C69Y, C69W, C69*, P70S, P70H, P70R, P70A, P70T, P70L, G71S, G71D, G71W, G71V, Y72S, Y72D, Y72N, Y72H, Y72F, Y72C, R73L, R73C, R73P, R73H, W74C, W74G, W74R, W74*, W74S, W74L, M75K, M75L, M75R, M75S, M75V, M75T, M75I, C76G, C76*, C76R, C76W, C76S, C76F, C76Y, L77P, L77G, L77M, L77V, L77Q, L77R, R78I, R78G, R78W, R78P, R78L, R78Q, R79P, R79G, R79L, R79C, R79S, R79H, F80G, F80N, F80Y, F80I, F80L, F80S, I81S, I81Y, I81N, I81F, I81M, I81V, I81T, I82F, I82N, I82V, I82M, I82T, I82L, F83Y, F83I, F83L, F83C, F83S, L84F, L84H, L84P, F85Q, F85R, F85P, F85L, F85Y, F85S, F85C, I86M, I86K, I86L, I86F, I86V, I86T, L87M, L87V, L87R, L87Q, L87P, L88R, L88V, L88M, L88Q, L88P, L89V, L89Y, L89T, L89R, L89I, L89Q, L89P, C90G, C90W, C90N, C90I, C90*, C90Y, C90F, C90S, L91G, L91F, L91P, L91R, L91H, I92H, I92N, I92V, I92M, I92L, I92S, I92T, F93R, F93W, F93A, F93Y, F93I, F93L, F93C, F93S, L94G, L94*, L94V, L94F, L94M, L94W, L94S, L95P, L95F, L95M, L95V, L95S, L95*, L95W, V96I, V96E, V96N, V96F, V96L, V96C, V96D, V96A, V96G, L97C, L97R, L97V, L97H, L97F, L97P, V98M, V98W, V98A, V98Q, V98P, V98R, V98L, D99H, D99K, D99V, D99A, D99E, D99N, D99G, Y100P, Y100*, Y100K, Y100D, Y100H, Y100N, Y100L, Y100W, Y100S, Y100F, Y100C, Q101P, Q101L, Q101*, Q101K, Q101H, Q101R, G102N, G102R, G102A, G102D, G102C, G102S, G102V, M103K, M103R, M103L, M103T, M103V, M103I, L104Q, L104M, L104G, L104V, L104W, L104S, L104F, P105T, P105S, P105H, P105A, P105L, P105R, V106L, V106R, V106Q, V106D, V106E, V106G, V106I, V106A, C107A, C107W, C107*, C107G, C107S, C107Y, C107R, P108A, P108T, P108S, P108L, P108H, L109R, L109V, L109I, L109Q, L109M, L109P, I110R, I110H, I110V, I110N, I110F, I110S, I110P, I110T, I110M, I110L, A111N, A111R, A111T, A111Q, A111S, A111L, A111P, G112V, G112K, G112N, G112E, G112A, G112R, S113C, S113R, S113F, S113L, S113Y, S113K, S113P, S113N, S113A, S113T, T114V, T114*, T114L, T114R, T114N, T114M, T114K, T114A, T114P, T114S, T115P, T115S, T115A, T115I, T115N, T116S, T116P, T116A, T116I, T116N, S117Y, S117A, S117V, S117R, S117I, S117G, S117N, S117T, T118W, T118N, T118G, T118I, T118E, T118P, T118S, T118K, T118R, T118M, T118A, T118V, G119S, G119W, G119T, G119*, G119C, G119V, G119E, G119R, P120H, P120N, P120R, P120L, P120A, P120Q, P120S, P120T, C121T, C121G, C121F, C121L, C121Y, C121S, C121W, C121R, K122G, K122V, K122E, K122N, K122T, K122I, K122S, K122Q, K122R, T123G, T123V, T123P, T123S, T123N, T123I, T123A, C124G, C124R, C124W, C124Y, C124S, C124F, T125P, T125I, T125A, T125M, T126P, T126Q, T126K, T126G, T126M, T126L, T126V, T126N, T126A, T126S, T126I, P127F, P127R, P127V, P127H, P127S, P127A, P127I, P127T, P127L, A128T, A128R, A128G, A128V, Q129E, Q129K, Q129L, Q129P, Q129N, Q129H, Q129R, G130V, G130*, G130D, G130K, G130A, G130E, G130S, G130R, G130N, N131Q, N131G, N131D, N131H, N131K, N131S, N131I, N131A, N131P, N131T, S132H, S132Y, S132P, S132C, S132F, M133R, M133G, M133K, M133V, M133Q, M133S, M133I, M133L, M133T, F134K, F134D, F134W, F134T, F134Q, F134C, F134R, F134V, F134H, F134S, F134L, F134N, F134I, F134Y, P135S, P135R, P135T, P135H, P135L, S136T, S136C, S136A, S136L, S136P, S136F, S136Y, C137S, C137R, C137*, C137Y, C137W, C138S, C138*, C138W, C138G, C138R, C138Y, C139S, C139G, C139*, C139Y, C139W, C139R, T140P, T140K, T140M, T140A, T140L, T140I, T140S, K141N, K141Q, K141T, K141I, K141R, K141E, P142T, P142A, P142H, P142R, P142I, P142S, P142L, T143W, T143F, T143A, T143P, T143M, T143L, T143S, D144Y, D144N, D144V, D144A, D144G, D144E, G145D, G145V, G145I, G145S, G145K, G145E, G145A, G145R, N146I, N146T, N146K, N146S, N146D, C147W, C147S, C147R, C147G, C147Y, T148S, T148I, T148A, C149S, C149G, C149W, C149Y, C149R, I150S, I150N, I150M, I150V, I150T, P151S, P151R, P151T, P151L, P151H, I152H, I152F, I152L, I152T, I152V, P153A, P153T, P153L, P153Q, P153S, S154Q, S154A, S154H, S154V, S154T, S154P, S154L, S155T, S155A, S155Y, S155F, S155P, W156S, W156C, W156G, W156R, W156*, W156L, A157S, A157N, A157D, A157P, A157V, A157T, A157G, F158I, F158Y, F158S, F158L, A159P, A159Q, A159L, A159S, A159T, A159R, A159E, A159V, A159G, K160Q, K160T, K160E, K160G, K160S, K160N, K160R, Y161K, Y161C, Y161V, Y161H, Y161I, Y161L, Y161S, Y161F, L162S, L162*, L162R, L162V, L162I, L162P, L162Q, W163G, W163C, W163R, E164*, E164K, E164A, E164V, E164D, E164G, W165Q, W165*, W165G, W165C, W165S, W165R, W165L, A166S, A166D, A166P, A166T, A166V, A166G, S167Y, S167E, S167T, S167G, S167P, S167L, V168L, V168I, V168F, V168G, V168D, V168S, V16887T, V168P, V168A, R169G, R169L, R169S, R169C, R169P, R169H, F170V, F170L, F170S, S171N, S171Y, S171C, S171L, S171P, S171F, W172M, W172F, W172G, W172R, W172S, W172L, W172C, W172*, P173I, P173Y, P173S, P173V, P173F, P173L, S174G, S174K, S174C, S174R, S174T, S174I, S174N, L175I, L175*, L175F, L175S, L176I, L176V, L176Q, L176P, V177P, V177I, V177S, V177G, V177E, V177M, V177L, V177A, P178G, P178R, P178H, P178A, P178L, P178S, P178Q, F179I, F179C, F179L, F179Y, F179S, V180P, V180L, V180E, V180F, V180I, V180A, Q181K, Q181E, Q181L, Q181H, Q181R, W182F, W182G, W182S, W182R, W182C, W182L, W182*, F183V, F183Y, F183I, F183L, F183S, F183C, V184L, V184P, V184D, V184F, V184I, V184E, V184G, V184A, G185K, G185L, G185W, G185R, G185A, G185E, L186M, L186A, L186R, L186C, L186S, L186I, L186V, L186F, L186P, L186H, S187A, S187T, S187C, S187P, S187F, S187L, P188A, P188S, P188R, P188T, P188H, P188L, T189L, T189N, T189A, T189S, T189P, T189I, V190S, V190E, V190P, V190D, V190G, V190I, V190F, V190A, W191K, W191S, W191*, W191G, W191L, W191R, W191C, L192R, L192H, L192V, L192F, L192P, S193*, S193T, S193P, S193F, S193L, A194G, A194D, A194S, A194T, A194F, A194L, A194I, A194V, I195K, I195V, I195L, I195T, I195M, W196G, W196V, W196K, W196R, W196*, W196S, W196L, M197L, M197K, M197R, M197V, M197I, M197T, M198L, M198V, M198R, M198K, M198T, M198I, W199P, W199C, W199R, W199G, W199S, W199*, W199L, Y200H, Y200*, Y200L, Y200N, Y200S, Y200C, Y200F, W201C, W201L, W201G, W201S, W201R, W201*, G202V, G202E, G202R, G202A, P203H, P203T, P203G, P203S, P203A, P203L, P203Q, P203R, S204Q, S204H, S204C, S204D, S204I, S204T, S204G, S204K, S204R, S204N, L205Q, L205R, L205V, L205P, L205M, Y206G, Y206P, Y206Q, Y206*, Y206I, Y206W, Y206V, Y206D, Y206R, Y206L, Y206N, Y206S, Y206F, Y206H, Y206C, N207G, N207K, N207C, N207D, N207P, N207H, N207I, N207T, N207R, N207S, I208L, I208C, I208F, I208S, I208V, I208N, I208T, L209K, L209A, L209F, L209*, L209G, L209M, L209S, L209W, L209V, S210M, S210G, S210I, S210T, S210K, S210R, S210N, P211A, P211T, P211S, P211R, P211L, P211H, F212C, F212S, F212L, F212Y, I213A, I213V, I213S, I213F, I213T, I213M, I213L, P214G, P214R, P214T, P214H, P214S, P214Q, P214L, L215D, L215I, L215V, L215T, L215M, L215Q, L215R, L215P, L216T, L216G, L216C, L216R, L216Y, L216V, L216I, L216S, L216F, L216*, P217A, P217T, P217Q, P217S, P217L, I218P, I218K, I218S, I218F, I218N, I218T, I218L, F219C, F219I, F219L, F219S, F220V, F220I, F220S, F220W, F220Y, F220L, F220C, C221V, C221R, C221W, C221L, C221S, C221G, C221F, C221Y, H222Q, H222V, H222S, H222R, H222F, H222I, H222P, H222L, W223F, W223G, W223C, W223R, W223*, W223L, V224L, V224I, V224E, V224G, V224A, Y225A, Y225*, Y225N, Y225D, Y225L, Y225I, Y225H, Y225C, Y225F, Y225S, I226R, I226H, I226L, I226F, I226N, I226V, I226T, I226M, I226S, | |||||
| Epitope Position | Core AA Sequence | HLA_ DRB*1/5_ Genotype A | Epitopes Position | Core AA Sequence | HLA_ DRB*1/5_ Genotype D | Mutations Relative to Protein: (WT, aa#, Mut) | |
|---|---|---|---|---|---|---|---|
| A | D | ||||||
| 84–92 | ILATVPAVP ¥ | *0802 | 73–81 | ILQTLPANP | I84V, I84T, A86V, A86T, V88M, V88L, A90P, A90V, P92L | I73M, L75H, L77M, L77V, A79E | |
| 85–93 | LATVPAVPP ¥ | *˜0802 | 74–82 | LQTLPANPP | I84V, I84T, A86V, A86T, V88M, V88L, A90P, A90V, P92L | I73M, L75H, L77M, L77V, A79E | |
| 34–42 | FGANSNNPD | 23–30 | FRANTANP ¥ | *0401 | - | R24K | |
| 16–24 | LSVPNPLGF | 5–13 | LSTSNPLGF ¥ | *0401, *1302 | - | - | |
| 24–32 | FFPDHQLDP | 13–21 | FFPDHQLDP | *0401, | F25L | - | |
| 63–71 | FGPGFTPPH | 52–60 | FGLGFTPPH | *0101, *0401, *0701, 5*0101 | F67L | L54M, L54P, F56L | |
| 34–42 | FGANSNNPD | 23–30 | FRANTANPD | *0101, *0401, *0802, *1302 | - | R24K | |
| 67–75 | FTPPHGGVL | 56–64 | FTPPHGGLL | *0101, *0701, | F67L, G73R | - | |
| 28–36 | HQLDPAFGA | 17–25 | HQLDPAFRA | *0301, *0401, | - | R24K | |
| 84–92 | ILATVPAVP | *0101, *0301, *0401, *0701, *0802, *1101, *1302, *1501, 5*0101 | 73–81 | ILQTLPANP | *0101, *0301, *0401, *0701, *0802,*1302, | I84V, I84T, A86V, A86T, V88M, V88L, A90P, A90V, P92L | I73M, L75H, L77M, L77V, A79E |
| 85–93 | LATVPAVPP | *0401, *0802, 5*0101 | 74–82 | LQTLPANPP | *0802, 5*0101 | I84V, I84T, A86V, A86T, V88M, V88L, A90P, A90V, P92L | I73M, L75H, L77M, L77V, A79E |
| 74–82 | LLGWSPQAQ | *1501 | 63–71 | *0101, *0401, *0802, *1501 | W77stop, S78R, P79S, A81S, A81P | S67N, P68S | |
| 16–24 | LSTSNPLGF | 5–13 | *0301, *0701, *1302 | - | - | ||
| 12–21 | MGTNLSVPN | *0401, *0802, *1302 | 1–10 | MGQNLSTSNP | - | G2E | |
| 15–22 | NLSVPNPLG | *0401, | 4–12 | NLSTSNPLGF | - | - | |
| 83–90 | QGILATVPA | *0101, *0802 | 71–79 | QGILQTLPA | *0101, | G83D, I84V, I84K, A86T, A86V, V88L, V88M, A90T, A90V | I73M, L75H, L77M, L77V, A79E |
| 14–22 | TNLSVPNPL | *0101, *0701, *1302 | 3–11 | QNLSTSNPL | *0101, *0401, | - | - |
| 4–12 | WSAKPRKGM | *1101, 5*0101 | - | S5P, A6S, A6T, K10N, K10I | - | ||
| 77–85 | WSPQAQGIL | *0101, *0701, | 66–75 | WSPQAQGIL | *0101, *0701, | W77stop, S78R, P79S, A81P, A81S, G83D, I84V, I84K | P68S, |
| Genotype | AA Sequence Core | AA Sequence of epi | Pres2 epi Residues | Count of epi with Core AA Sequence | HLA Class II Alleles | Mutations in the Core Sequence |
|---|---|---|---|---|---|---|
| A | ALQDPRVRG | AFHQALQDPRVRGLYFPA | 7–24 | 7 | *0301∆ | R16K, V17I |
| A | ASHISSISS | VPNTASHISSISSRT | 35–49 | *0401 | A39T, I45S | |
| D | ASPLSSIFS | VPTTASPLSSIFSRIG | 35–50 | 2 | *0101, *0401 | L42I, L42S |
| A | FHQALQDPR | NSTAFHQALQDPRVRG | 4–19 | 2 | *0301, 5*0101 | A11T, R16K |
| D | FHQTLQDPR | NSTTFHQTLQDPRVR | 4–18 | 1 | *0301 | - |
| D | FSRIGDPAL | SPLSSIFSRIGDPALN | 40–55 | 1 | *0101, *0401, *0701, 5*0101 | R48T, P52H, A53V, L54P |
| A | HISSISSRT | VPNTASHISSISSRTG | 35–50 | 4 | *0101, *0401, *0701 | I45S, R48T |
| D | IFSRIGDPA | SPLSSIFSRIGDPALN | 40–55 | 2 | *0802, *1302, *1501 | L42I, L42S |
| A | ISSISSRTG | PNTASHISSISSRTGDPALN | 36–55 | 6 | *0101, *0401, *0701, *0802, *1101, *1501, 5*0101 | I45S, R48T |
| A | LNPVPNTAS | SSSGTLNPVPNTASHISSI | 27–45 | 5 | *0401, *0802 | L32H, L32R, P34L, N37H, N37T, I38T, A38T |
| D | LSSIFSRIG | PTTASPLSSIFSRIGDPALN | 36–55 | 6 | *0101, *0401, *0701, *0802, *1101, *1501, 5*0101 | P52L, A53V |
| A | MQWNSTAFH | MQWNSTAFHQALQDP | 1–15 | 1 | *1302 | Q2I, A7T |
| MQWNSTTFHQTLQDP | - | - | S5F, S5Y, T6A, T7I, T7N | |||
| D | PLSSIFSRI | VPTTASPLSSIFSRI | 35–49 | 1 | *0701 | L42I, L42S |
| A | PVPNTASHI | SSGTLNPVPNTASHISSI | 27–45 | 6 | *1302 | P34L, N37H, N37T |
| D | PVPTTASPL | VNPVPTTASPLSSIF | 32–46 | 2 | *0701 | P34H, P36L, L42I, L42S |
| A | QALQDPRVR | TAFHQALQDPRVRGLYF | 6–22 | 3 | 5*0101 | - |
| D | QTLQDPRVR | STTFHQTLQDPRVRGLYF | 5–22 | 4 | 5*0101 | R22K |
| D | TLQDPRVRG | STTFHQTLQDPRVRGLYFPA | 6 | *0301 | R18K, G19D, G19A | |
| A | VRGLYFPAG | DPRVRGLYFPAGGSSSG | 14–30 | 3 | *0802, *1101, *1501 | V17I, Y21N, Y21S, F22L, F22P, F22T, P23A |
| D | 3 | *0802, *1101, *1501 | R18K, G19D, G19A, F2222Q, F22H, F22P, F22L | |||
| A | WNSTAFHQA | MQWNSTAFHQALQDP | 1–15 | 1 | *0401, *0701 | A7T, A11T |
| D | WNSTTFHQT | MQWNSTTFHQTLQDP | 1 | *0701 | S5F, S5Y, T6A, T7I, T7N | |
| A | YFPAGGSSS | PRVRGLYFPAGGSSSGTLNP | 15–34 | 6 | *0101, *0401, *1101, 5*0101 | Y21N, Y21S, F22L, F22P, F22T, P23A, S29L |
| D | YFPAGGSSS | PRVRGLYFPAGGSSSGTVNP | 6 | *0101, *0401, 5*0101, *1101 | F22Q, F22H, F22P, F22L |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choga, W.T.; Anderson, M.; Zumbika, E.; Phinius, B.B.; Mbangiwa, T.; Bhebhe, L.N.; Baruti, K.; Kimathi, P.O.; Seatla, K.K.; Musonda, R.M.; et al. In Silico Prediction of Human Leukocytes Antigen (HLA) Class II Binding Hepatitis B Virus (HBV) Peptides in Botswana. Viruses 2020, 12, 731. https://doi.org/10.3390/v12070731
Choga WT, Anderson M, Zumbika E, Phinius BB, Mbangiwa T, Bhebhe LN, Baruti K, Kimathi PO, Seatla KK, Musonda RM, et al. In Silico Prediction of Human Leukocytes Antigen (HLA) Class II Binding Hepatitis B Virus (HBV) Peptides in Botswana. Viruses. 2020; 12(7):731. https://doi.org/10.3390/v12070731
Chicago/Turabian StyleChoga, Wonderful Tatenda, Motswedi Anderson, Edward Zumbika, Bonolo B. Phinius, Tshepiso Mbangiwa, Lynnette N. Bhebhe, Kabo Baruti, Peter Opiyo Kimathi, Kaelo K. Seatla, Rosemary M. Musonda, and et al. 2020. "In Silico Prediction of Human Leukocytes Antigen (HLA) Class II Binding Hepatitis B Virus (HBV) Peptides in Botswana" Viruses 12, no. 7: 731. https://doi.org/10.3390/v12070731
APA StyleChoga, W. T., Anderson, M., Zumbika, E., Phinius, B. B., Mbangiwa, T., Bhebhe, L. N., Baruti, K., Kimathi, P. O., Seatla, K. K., Musonda, R. M., Bell, T. G., Moyo, S., Blackard, J. T., & Gaseitsiwe, S. (2020). In Silico Prediction of Human Leukocytes Antigen (HLA) Class II Binding Hepatitis B Virus (HBV) Peptides in Botswana. Viruses, 12(7), 731. https://doi.org/10.3390/v12070731