Live Visualization of Hemagglutinin Dynamics during Infection by Using a Novel Reporter Influenza A Virus

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines, Antibodies, and Drugs
2.2. IAV Infection of Cells
2.3. Plaque Assay
2.4. Virus Rescue
2.5. Virus Like Particle Formation and Hemagglutination Assay
2.6. Immunostaining
2.7. Western Blotting
2.8. Biarsenic Labelling and Live Imaging of Infected Cells
2.9. Imaging Data Analysis
2.10. Software and Statistical Analysis
3. Results
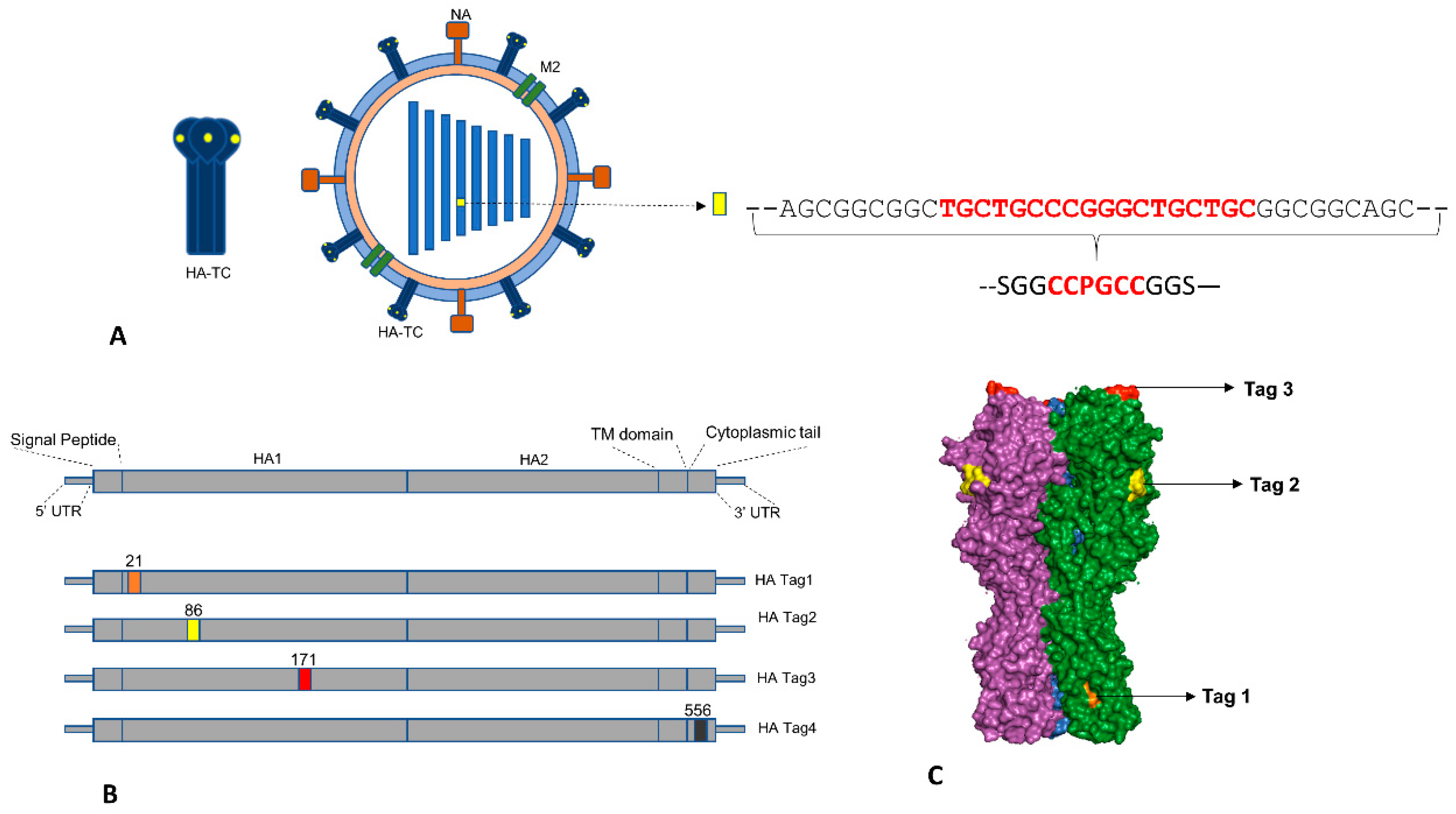
3.1. Strategy for Generation of Reporter IAV Expressing Hemagglutinin with Tetra-Cysteine Tag
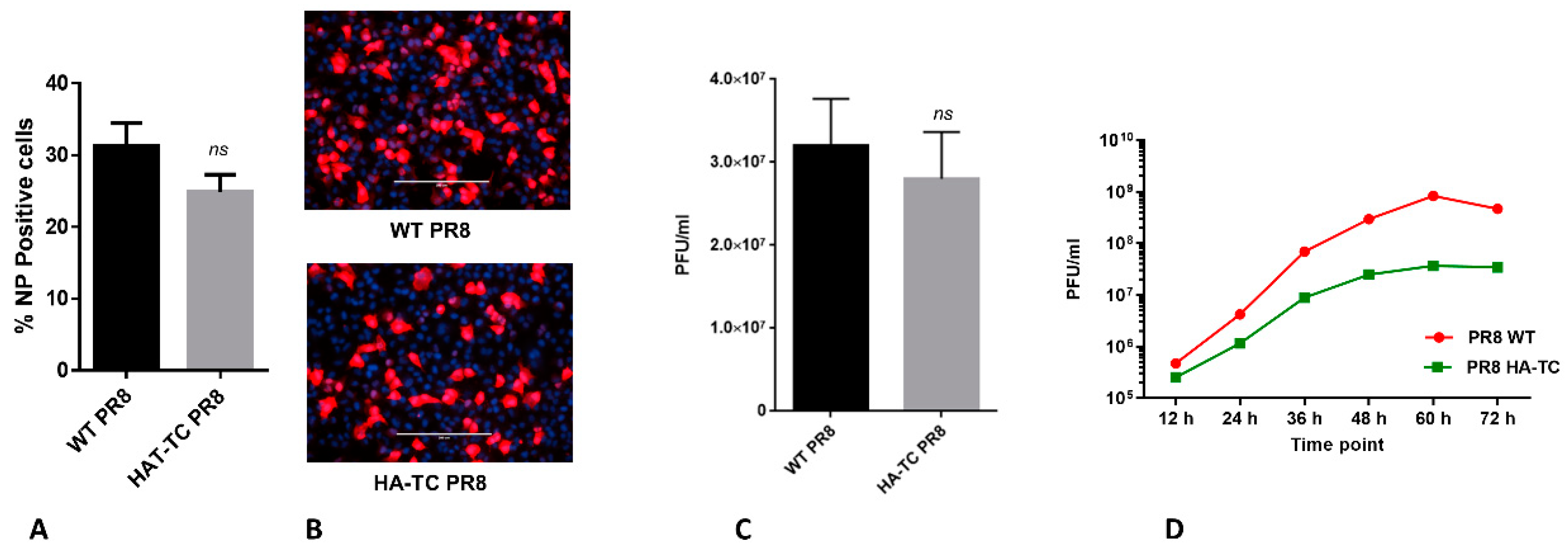
3.2. Rescue and Characterization of Reporter PR8 IAV Expressing HA-TC Protein
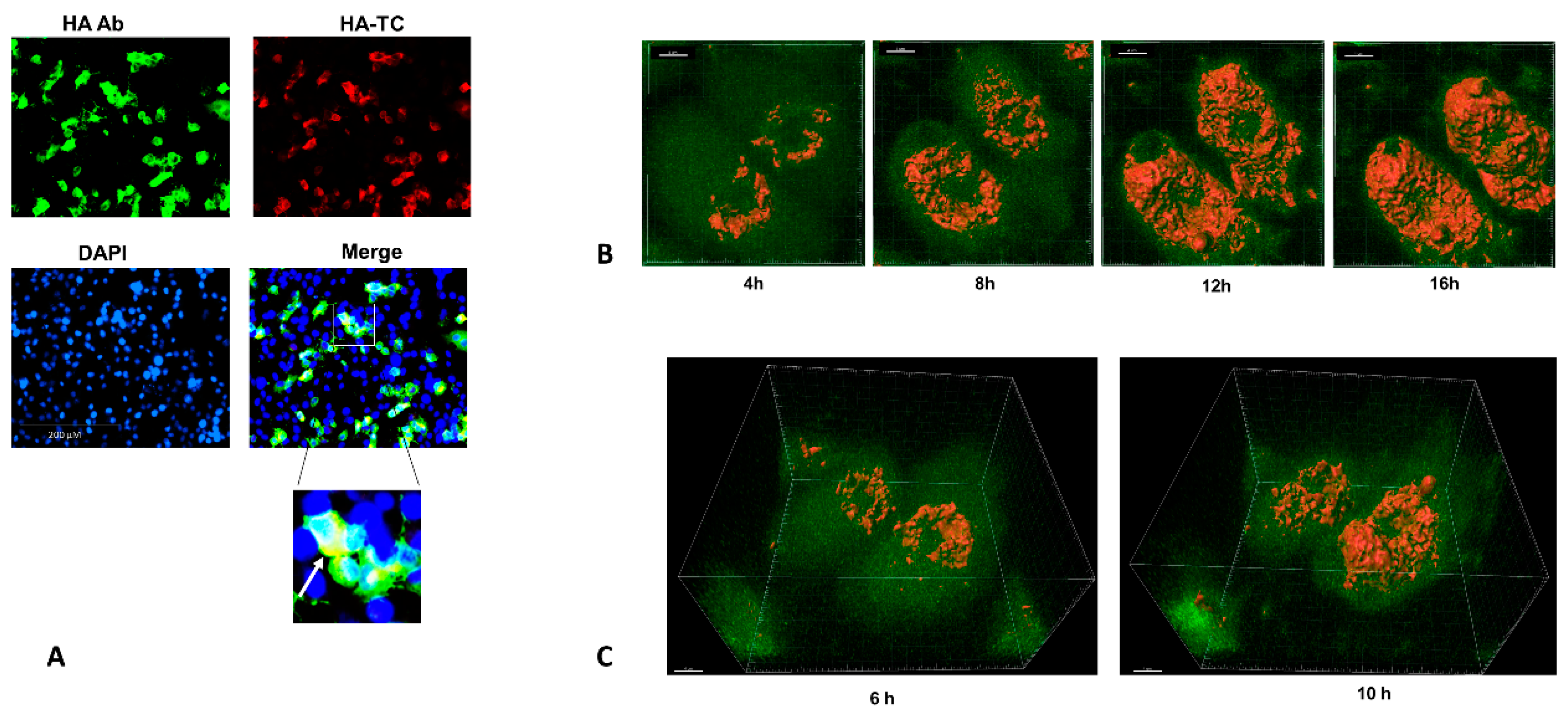
3.3. Four Dimensional Live Imaging of HA in Cells Infected with HA-TC Reporter IAV
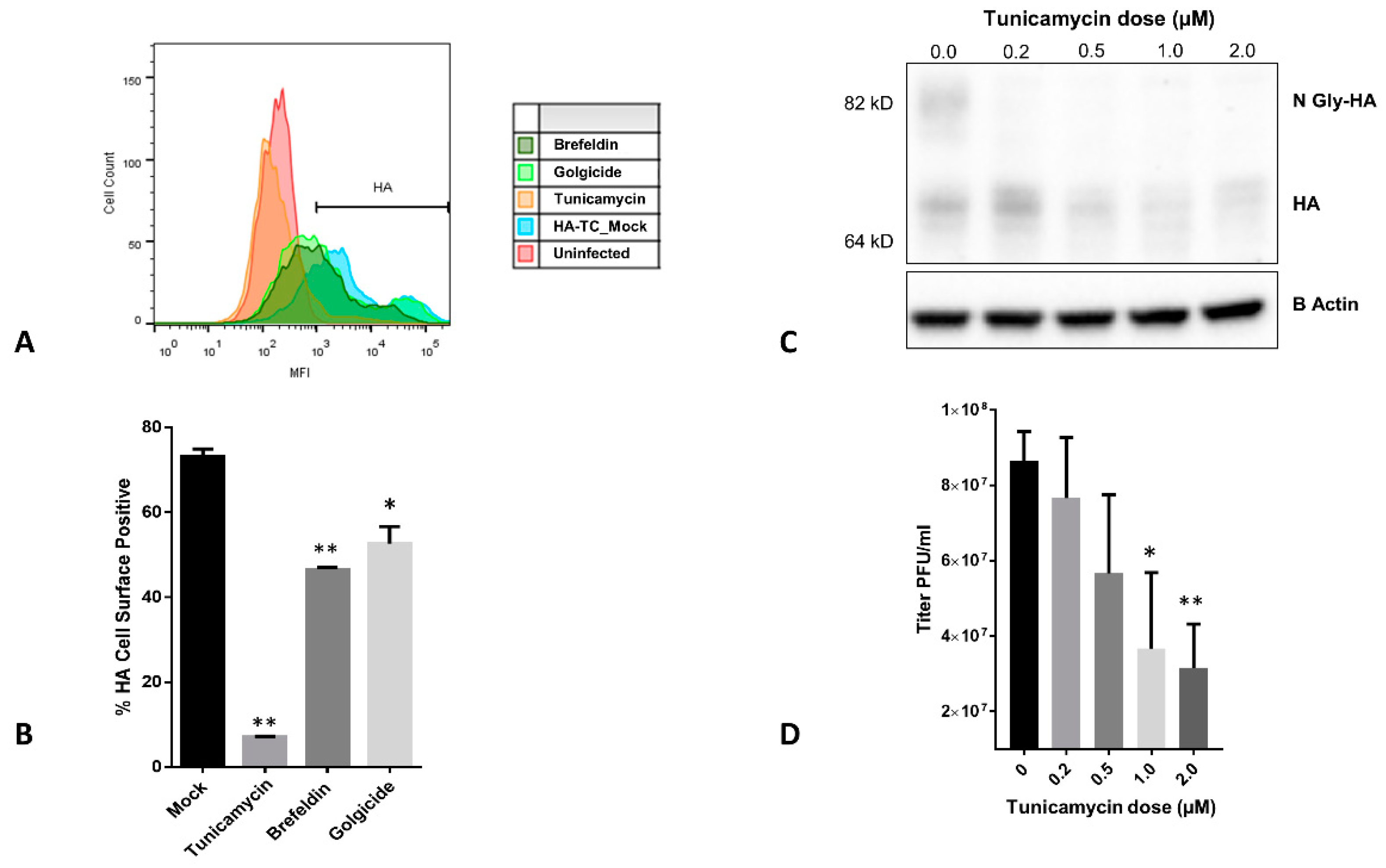
3.4. Effect of ER-Golgi Transport Inhibitors on HA Trafficking in IAV Infected Cells
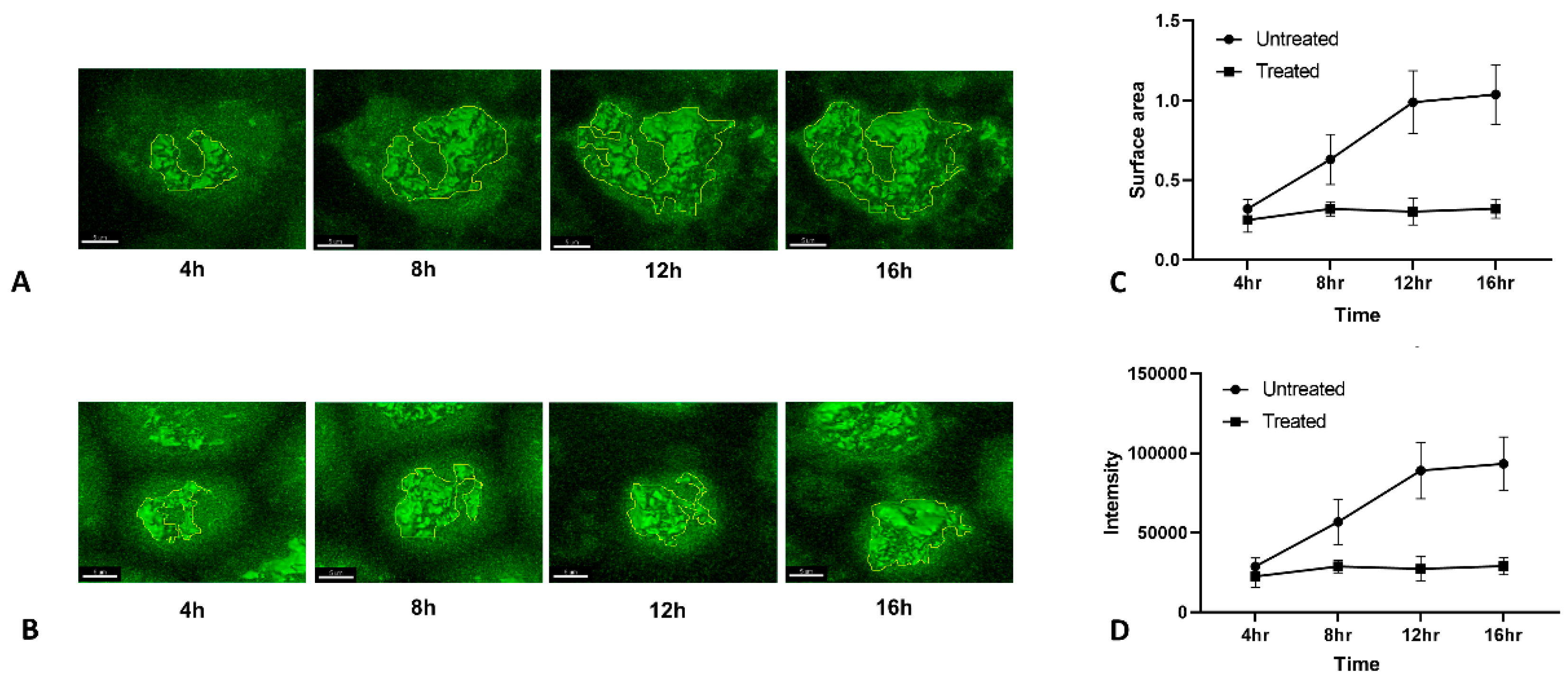
3.5. Quantiative Live Imaging of HA Transport Retardation by Tunicamycin
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Medina, R.A.; Garcia-Sastre, A. Influenza A viruses: New research developments. Nat. Rev. Microbiol. 2011, 9, 590–603. [Google Scholar] [CrossRef]
- Russell, C.J.; Hu, M.; Okda, F.A. Influenza Hemagglutinin Protein Stability, Activation, and Pandemic Risk. Trends. Microbiol. 2018, 26, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, C.P.; Witte, M.D.; Theile, C.S.; Bozkurt, G.; Kundrat, L.; Blom, A.E.; Ploegh, H.L. Site-specific C-terminal and internal loop labeling of proteins using sortase-mediated reactions. Nat. Protoc. 2013, 8, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Heaton, N.S.; Sachs, D.; Chen, C.J.; Hai, R.; Palese, P. Genome-wide mutagenesis of influenza virus reveals unique plasticity of the hemagglutinin and NS1 proteins. Proc. Natl. Acad. Sci. USA 2013, 110, 20248–20253. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Rodrigues, M.; Rodriguez, D.; Rodriguez, J.R.; Esteban, M.; Palese, P.; Nussenzweig, R.S.; Zavala, F. Priming with recombinant influenza virus followed by administration of recombinant vaccinia virus induces CD8+ T-cell-mediated protective immunity against malaria. Proc. Natl. Acad. Sci. USA 1993, 90, 5214–5218. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.N.; Mueller, S.N.; Ye, L.; Bu, Z.; Yang, C.; Ahmed, R.; Steinhauer, D.A. Chimeric influenza virus hemagglutinin proteins containing large domains of the Bacillus anthracis protective antigen: Protein characterization, incorporation into infectious influenza viruses, and antigenicity. J. Virol. 2005, 79, 10003–10012. [Google Scholar] [CrossRef] [PubMed]
- Griffin, B.A.; Adams, S.R.; Tsien, R.Y. Specific covalent labeling of recombinant protein molecules inside live cells. Science 1998, 281, 269–272. [Google Scholar] [CrossRef]
- Charlier, C.M.; Wu, Y.J.; Allart, S.; Malnou, C.E.; Schwemmle, M.; Gonzalez-Dunia, D. Analysis of borna disease virus trafficking in live infected cells by using a virus encoding a tetracysteine-tagged p protein. J. Virol. 2013, 87, 12339–12348. [Google Scholar] [CrossRef]
- Das, S.C.; Panda, D.; Nayak, D.; Pattnaik, A.K. Biarsenical labeling of vesicular stomatitis virus encoding tetracysteine-tagged m protein allows dynamic imaging of m protein and virus uncoating in infected cells. J. Virol. 2009, 83, 2611–2622. [Google Scholar] [CrossRef]
- Du, J.; Bhattacharya, B.; Ward, T.H.; Roy, P. Trafficking of bluetongue virus visualized by recovery of tetracysteine-tagged virion particles. J. Virol. 2014, 88, 12656–12668. [Google Scholar] [CrossRef]
- Gousset, K.; Ablan, S.D.; Coren, L.V.; Ono, A.; Soheilian, F.; Nagashima, K.; Ott, D.E.; Freed, E.O. Real-time visualization of HIV-1 GAG trafficking in infected macrophages. PLoS Pathog. 2008, 4, e1000015. [Google Scholar] [CrossRef]
- Li, Y.; Lu, X.; Li, J.; Berube, N.; Giest, K.L.; Liu, Q.; Anderson, D.H.; Zhou, Y. Genetically engineered, biarsenically labeled influenza virus allows visualization of viral NS1 protein in living cells. J. Virol. 2010, 84, 7204–7213. [Google Scholar] [CrossRef] [PubMed]
- Panchal, R.G.; Ruthel, G.; Kenny, T.A.; Kallstrom, G.H.; Lane, D.; Badie, S.S.; Li, L.; Bavari, S.; Aman, M.J. In vivo oligomerization and raft localization of Ebola virus protein VP40 during vesicular budding. Proc. Natl. Acad. Sci. USA 2003, 100, 15936–15941. [Google Scholar] [CrossRef] [PubMed]
- Vahey, M.D.; Fletcher, D.A. Low-Fidelity Assembly of Influenza A Virus Promotes Escape from Host Cells. Cell 2019, 176, 281–294.e219. [Google Scholar] [CrossRef] [PubMed]
- Whitt, M.A.; Mire, C.E. Utilization of fluorescently-labeled tetracysteine-tagged proteins to study virus entry by live cell microscopy. Methods 2011, 55, 127–136. [Google Scholar] [CrossRef]
- Lippincott-Schwartz, J.; Yuan, L.; Tipper, C.; Amherdt, M.; Orci, L.; Klausner, R.D. Brefeldin A’s effects on endosomes, lysosomes, and the TGN suggest a general mechanism for regulating organelle structure and membrane traffic. Cell 1991, 67, 601–616. [Google Scholar] [CrossRef]
- Saenz, J.B.; Sun, W.J.; Chang, J.W.; Li, J.; Bursulaya, B.; Gray, N.S.; Haslam, D.B. Golgicide A reveals essential roles for GBF1 in Golgi assembly and function. Nat. Chem. Biol. 2009, 5, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Van Raam, B.J.; Lacina, T.; Lindemann, R.K.; Reiling, J.H. Secretory stressors induce intracellular death receptor accumulation to control apoptosis. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef]
- Baumann, J.; Ignashkova, T.I.; Chirasani, S.R.; Ramirez-Peinado, S.; Alborzinia, H.; Gendarme, M.; Kuhnigk, K.; Kramer, V.; Lindemann, R.K.; Reiling, J.H. Golgi stress-induced transcriptional changes mediated by MAPK signaling and three ETS transcription factors regulate MCL1 splicing. Mol. Biol Cell 2018, 29, 42–52. [Google Scholar] [CrossRef]
- Manicassamy, B.; Manicassamy, S.; Belicha-Villanueva, A.; Pisanelli, G.; Pulendran, B.; Garcia-Sastre, A. Analysis of in vivo dynamics of influenza virus infection in mice using a GFP reporter virus. Proc. Natl. Acad. Sci. USA 2010, 107, 11531–11536. [Google Scholar] [CrossRef]
- Horton, R.M.; Cai, Z.L.; Ho, S.N.; Pease, L.R. Gene splicing by overlap extension: Tailor-made genes using the polymerase chain reaction. Biotechniques 1990, 8, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Watanabe, T.; Kawaoka, Y. Plasmid-driven formation of influenza virus-like particles. J. Virol. 2000, 74, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Nogales, A.; Avila-Perez, G.; Rangel-Moreno, J.; Chiem, K.; DeDiego, M.L.; Martinez-Sobrido, L. A Novel Fluorescent and Bioluminescent Bireporter Influenza A Virus To Evaluate Viral Infections. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Kuo, S.C.; Lampen, J.O. Tunicamycin—An inhibitor of yeast glycoprotein synthesis. Biochem. Biophys. Res. Commun. 1974, 58, 287–295. [Google Scholar] [CrossRef]
- Lakdawala, S.S.; Wu, Y.; Wawrzusin, P.; Kabat, J.; Broadbent, A.J.; Lamirande, E.W.; Fodor, E.; Altan-Bonnet, N.; Shroff, H.; Subbarao, K. Influenza a virus assembly intermediates fuse in the cytoplasm. PLoS Pathog. 2014, 10, e1003971. [Google Scholar] [CrossRef]
- Lambert Emo, K.; Hyun, Y.M.; Reilly, E.; Barilla, C.; Gerber, S.; Fowell, D.; Kim, M.; Topham, D.J. Live Imaging of Influenza Infection of the Trachea Reveals Dynamic Regulation of CD8+ T Cell Motility by Antigen. PLoS Pathog. 2016, 12, e1005881. [Google Scholar] [CrossRef]
- Popp, M.W.; Karssemeijer, R.A.; Ploegh, H.L. Chemoenzymatic site-specific labeling of influenza glycoproteins as a tool to observe virus budding in real time. PLoS Pathog. 2012, 8, e1002604. [Google Scholar] [CrossRef]
- Sato, R.; Okura, T.; Kawahara, M.; Takizawa, N.; Momose, F.; Morikawa, Y. Apical Trafficking Pathways of Influenza A Virus HA and NA via Rab17- and Rab23-Positive Compartments. Front. Microbiol. 2019, 10, 1857. [Google Scholar] [CrossRef]
- Davtyan, H.; Ghochikyan, A.; Cadagan, R.; Zamarin, D.; Petrushina, I.; Movsesyan, N.; Martinez-Sobrido, L.; Albrecht, R.A.; Garcia-Sastre, A.; Agadjanyan, M.G. The immunological potency and therapeutic potential of a prototype dual vaccine against influenza and Alzheimer’s disease. J. Transl. Med. 2011, 9, 127. [Google Scholar] [CrossRef]
- Rust, M.J.; Lakadamyali, M.; Zhang, F.; Zhuang, X. Assembly of endocytic machinery around individual influenza viruses during viral entry. Nat. Struct. Mol. Biol. 2004, 11, 567–573. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Anjos Borges, L.G.; Pisanelli, G.; Khatun, O.; García-Sastre, A.; Tripathi, S. Live Visualization of Hemagglutinin Dynamics during Infection by Using a Novel Reporter Influenza A Virus. Viruses 2020, 12, 687. https://doi.org/10.3390/v12060687
dos Anjos Borges LG, Pisanelli G, Khatun O, García-Sastre A, Tripathi S. Live Visualization of Hemagglutinin Dynamics during Infection by Using a Novel Reporter Influenza A Virus. Viruses. 2020; 12(6):687. https://doi.org/10.3390/v12060687
Chicago/Turabian Styledos Anjos Borges, Luiz Gustavo, Giuseppe Pisanelli, Oyahida Khatun, Adolfo García-Sastre, and Shashank Tripathi. 2020. "Live Visualization of Hemagglutinin Dynamics during Infection by Using a Novel Reporter Influenza A Virus" Viruses 12, no. 6: 687. https://doi.org/10.3390/v12060687
APA Styledos Anjos Borges, L. G., Pisanelli, G., Khatun, O., García-Sastre, A., & Tripathi, S. (2020). Live Visualization of Hemagglutinin Dynamics during Infection by Using a Novel Reporter Influenza A Virus. Viruses, 12(6), 687. https://doi.org/10.3390/v12060687