Development and Evaluation of a duo SARS-CoV-2 RT-qPCR Assay Combining Two Assays Approved by the World Health Organization Targeting the Envelope and the RNA-Dependant RNA Polymerase (RdRp) Coding Regions

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Monoplex RT-qPCR Assays
2.2. Generation of In Vitro Transcribed RNAs (IVT RNAs)
2.3. RT-qPCR
2.4. Analytical Sensitivity
2.5. Clinical Samples for SARS-CoV-2 RNA Detection
2.6. Neutralization Assay
3. Results
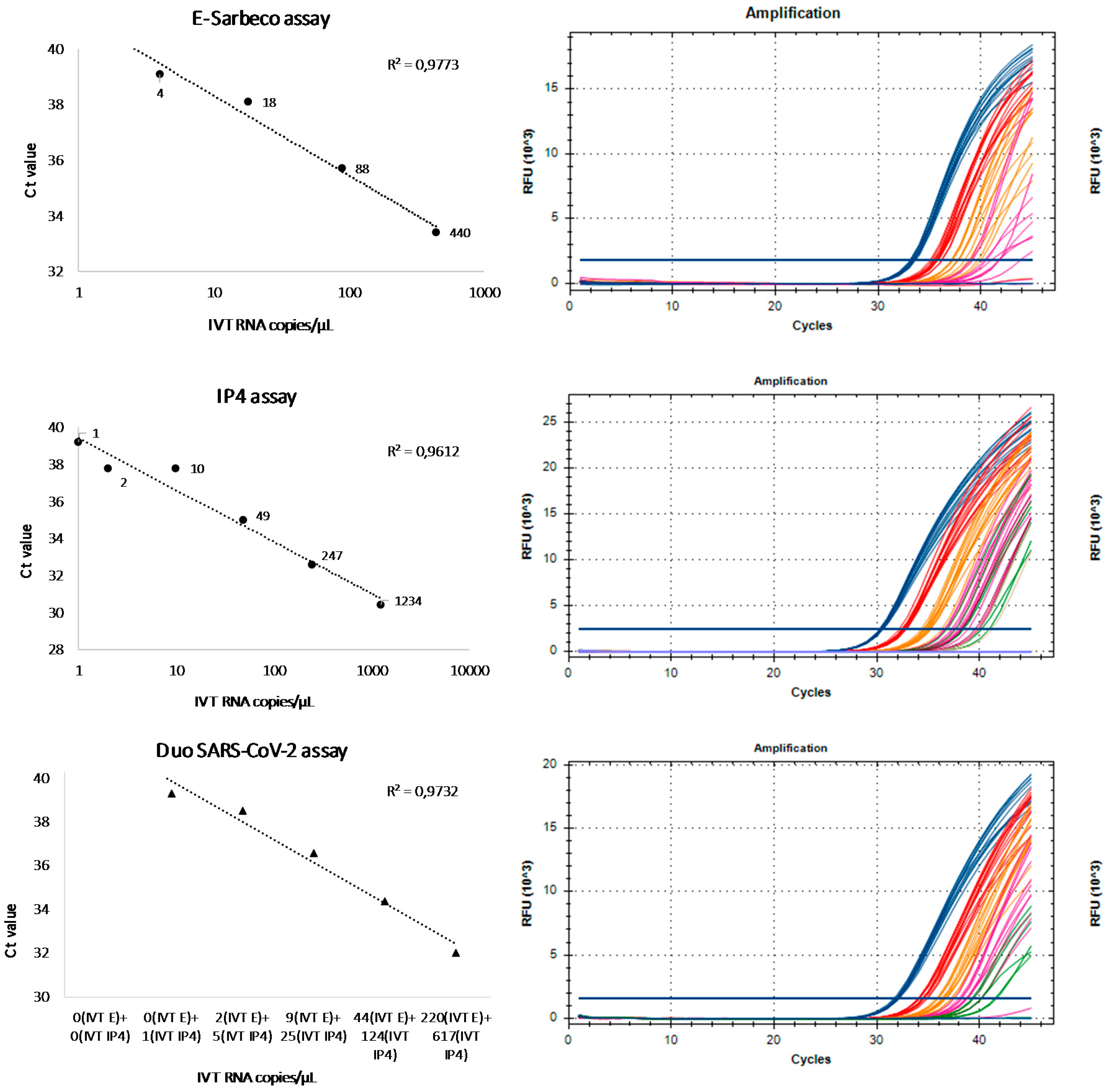
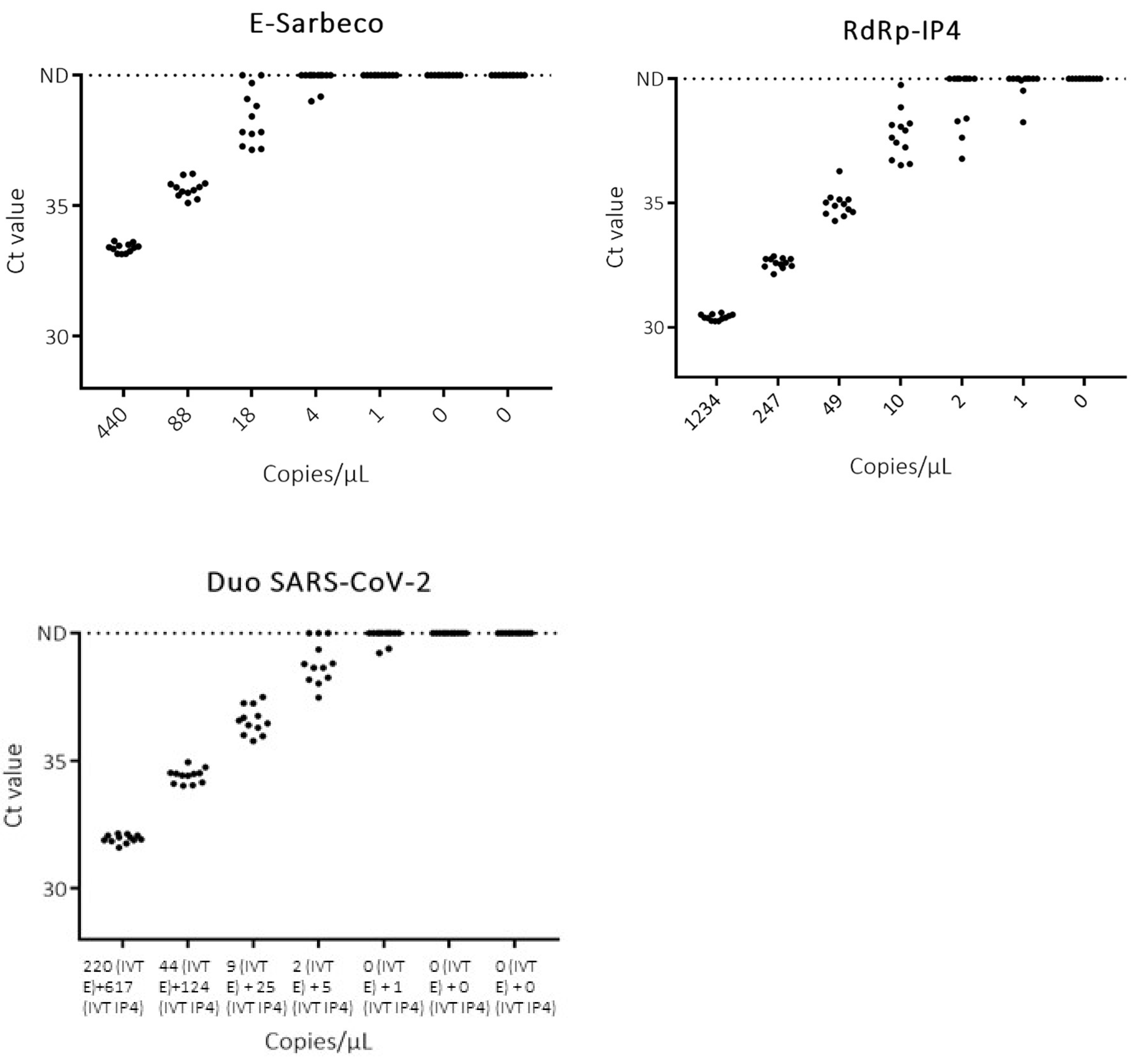
3.1. Analytical Sensitivity
3.2. Linearity and Signal Intensity
3.3. Clinical Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Molecular Assays to Diagnose COVID-19: Summary Table of Available Protocols. Available online: https://www.who.int/who-documents-detail/molecular-assays-to-diagnose-covid-19-summary-table-of-available-protocols (accessed on 25 June 2020).
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.W.; Pan, Y.; Cheng, S.M.S.; Hui, K.P.Y.; Krishnan, P.; Liu, Y.; Ng, D.Y.M.; Wan, C.K.C.; Yang, P.; Wang, Q.; et al. Molecular Diagnosis of a Novel Coronavirus (2019-nCoV) Causing an Outbreak of Pneumonia. Clin. Chem. 2020, 66, 549–555. [Google Scholar] [CrossRef]
- Vogels, C.B.F.; Brito, A.F.; Wyllie, A.L.; Fauver, J.R.; Ott, I.M.; Kalinich, C.C.; Petrone, M.E.; Casanovas-Massana, A.; Muenker, M.C.; Moore, A.J.; et al. Analytical sensitivity and efficiency comparisons of SARS-COV-2 qRT-PCR assays. medRxiv 2020. [Google Scholar] [CrossRef]
- Thirion, L.; Pezzi, L.; Corcostegui, I.; Dubot-Pérès, A.; Falchi, A.; de Lamballerie, X.; Charrel, R.N. Development and Evaluation of a Duo Chikungunya Virus Real-Time RT-PCR Assay Targeting Two Regions within the Genome. Viruses 2019, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- Thirion, L.; Charrel, R.N.; Boehmann, Y.; Corcostegui, I.; Raoul, H.; de Lamballerie, X. Development and Evaluation of a Duo Zaire ebolavirus Real-Time RT-PCR Assay Targeting Two Regions within the Genome. Microorganisms 2019, 7, 652. [Google Scholar] [CrossRef] [PubMed]
- Ninove, L.; Nougairede, A.; Gazin, C.; Thirion, L.; Delogu, I.; Zandotti, C.; Charrel, R.N.; De Lamballerie, X. RNA and DNA bacteriophages as molecular diagnosis controls in clinical virology: A comprehensive study of more than 45,000 routine PCR tests. PLoS ONE 2011, 6, e16142. [Google Scholar] [CrossRef] [PubMed]
- Alwassouf, S.; Christodoulou, V.; Bichaud, L.; Ntais, P.; Mazeris, A.; Antoniou, M.; Charrel, R.N. Seroprevalence of Sandfly-Borne Phleboviruses Belonging to Three Serocomplexes (Sandfly fever Naples, Sandfly fever Sicilian and Salehabad) in Dogs from Greece and Cyprus Using Neutralization Test. PLoS Negl. Trop. Dis. 2016, 10, e0005063. [Google Scholar] [CrossRef] [PubMed]
- Ayhan, N.; Sherifi, K.; Taraku, A.; Bërxholi, K.; Charrel, R.N. High Rates of Neutralizing Antibodies to Toscana and Sandfly Fever Sicilian Viruses in Livestock, Kosovo. Emerg. Infect. Dis. 2017, 23, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Pierro, A.; Ficarelli, S.; Ayhan, N.; Morini, S.; Raumer, L.; Bartoletti, M.; Mastroianni, A.; Prati, F.; Schivazappa, S.; Cenni, P.; et al. Characterization of antibody response in neuroinvasive infection caused by Toscana virus. Clin. Microbiol. Infect. 2017, 23, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Sizmann, D.; Glaubitz, J.; Simon, C.O.; Goedel, S.; Buergisser, P.; Drogan, D.; Hesse, M.; Kröh, M.; Simmler, P.; Dewald, M.; et al. Improved HIV-1 RNA quantitation by COBAS AmpliPrep/COBAS TaqMan HIV-1 Test, v2.0 using a novel dual-target approach. J. Clin. Virol. 2010, 49, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Templer, S.P.; Seiverth, B.; Baum, P.; Stevens, W.; Seguin-Devaux, C.; Carmona, S. Improved Sensitivity of a Dual-Target HIV-1 Qualitative Test for Plasma and Dried Blood Spots. J. Clin. Microbiol. 2016, 54, 1877–1882. [Google Scholar] [CrossRef]
- Van Dorp, L.; Acman, M.; Richard, D.; Shaw, L.P.; Ford, C.E.; Ormond, L.; Owen, C.J.; Pang, J.; Tan, C.C.; Boshier, F.A.; et al. Emergence of genomic diversity and recurrent mutations in SARS-CoV-2. Infect. Genet. Evol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Coronavirus Disease 2019 (COVID-19)in the EU/EEA and the UK –Eighth Update. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/covid-19-rapid-risk-assessment-coronavirus-disease-2019-eighth-update-8-april-2020.pdf (accessed on 25 June 2020).
- Wölfel, R.; Corman, V.M.; Guggemos, W.; Seilmaier, M.; Zange, S.; Müller, M.A.; Niemeyer, D.; Jones, T.C.; Vollmar, P.; Rothe, C.; et al. Virological assessment of hospitalized patients with COVID-2019. Nature 2020, 58, 465–469. [Google Scholar] [CrossRef] [PubMed]
- An analysis of SARS-CoV-2 Viral Load by Patient Age. Available online: https://zoonosen.charite.de/fileadmin/user_upload/microsites/m_cc05/virologie-ccm/dateien_upload/Weitere_Dateien/analysis-of-SARS-CoV-2-viral-load-by-patient-age.pdf (accessed on 25 June 2020).
- Thirion, L.; Dubot-Peres, A.; Pezzi, L.; Corcostegui, I.; Touinssi, M.; de Lamballerie, X.; Charrel, R.N. Lyophilized Matrix Containing Ready-to-Use Primers and Probe Solution for Standardization of Real-Time PCR and RT-qPCR Diagnostics in Virology. Viruses 2020, 12, 159. [Google Scholar] [CrossRef] [PubMed]
- Primers, Probe and Encapsidated RNA pos. Control - 2019-nCoV E gene, Large Packaging Ref-SKU 001B-03888. Available online: https://www.european-virus-archive.com/bundle/primers-probe-and-encapsidated-rna-pos-control-2019-ncov-e-gene-large-packaging (accessed on 25 June 2020).
- Lyophilized Primers and Probe for RT-PCR in the SARS-CoV-2 RdRp (IP4) gene Ref-SKU 001K-03952. Available online: https://www.european-virus-archive.com/detection-kit/lyophilized-primers-and-probe-rt-pcr-sars-cov-2-rdrp-ip4-gene (accessed on 25 June 2020).
- Primers, Probe and Encapsidated RNA pos. Control - SARS-CoV-2 RdRp (IP4) Gene, Small Packaging Ref-SKU 001B-03955. Available online: https://www.european-virus-archive.com/bundle/primers-probe-and-encapsidated-rna-pos-control-sars-cov-2-rdrp-ip4-gene-small-packaging (accessed on 25 June 2020).




{kind=link}
{kind=link}
{kind=link}
| Reference | Institute | Primer/Probe | 5′→3′ Sequence | Target | Position a | Amplicon Size | Concentration |
|---|---|---|---|---|---|---|---|
| [3] | Charité (Berlin) | E_Sarbeco_F | ACAGGTACGTTAATAGTTAATAGCGT | E gene | 26,269–26,294 | 113 nt | 400 nM |
| E_Sarbeco_R | ATATTGCAGCAGTACGCACACA | 26,360–26,381 | 400 nM | ||||
| E_Sarbeco_P | FAM-ACACTAGCCATCCTTACTGCGCTTCG-QSY | 26,332–26,357 | 200 nM | ||||
| [2] | Pasteur (Paris) | nCoV_IP4-14059Fw | GGTAACTGGTATGATTTCG | RdRp gene | 14,080–14,098 | 107 nt | 400 nM |
| nCoV_IP4-14146Rv | CTGGTCAAGGTTAATATAGG | 14,167–14,186 | 400 nM | ||||
| nCoV_IP4-14084Probe | FAM-TCATACAAACCACGCCAGG-QSY | 14,105–14,123 | 200 nM |
| E-Sarbeco Assay (Charité) | ||||
| IVT RNA copies/µL | Total samples tested, No. | Positive samples, No. | % Detected | Ct, Mean (SD) |
| 440 | 12 | 12 | 100 | 33.4 (0.2) |
| 88 | 12 | 12 | 100 | 35.7 (0.3) |
| 18 | 12 | 10 | 83 | 38.1 (1.0) |
| 4 | 12 | 2 | 17 | 39.1 (0.1) |
| 1 | 12 | 0 | 0 | - |
| 0 | 12 | 0 | 0 | - |
| 0 | 12 | 0 | 0 | - |
| RdRp-IP4 Assay (Institut Pasteur) | ||||
| IVT RNA copies/µL | Total samples tested, No. | Positive samples, No. | % Detected | Ct, Mean (SD) |
| 1234 | 12 | 12 | 100 | 30.4 (0.1) |
| 247 | 12 | 12 | 100 | 32.6 (0.2) |
| 49 | 12 | 12 | 100 | 35 (0.5) |
| 10 | 12 | 12 | 100 | 37.8 (1) |
| 2 | 12 | 4 | 33 | 37.8 (0.7) |
| 1 | 12 | 3 | 25 | 39.2 (0.9) |
| 0 | 12 | 0 | 0 | - |
| Duo SARS-CoV-2 Assay (This Study) | ||||
| IVT RNA copies/µL | Total samples tested, No. | Positive samples, No. | % Detected | Ct, Mean (SD) |
| 220 (IVT RNA E-Sarbeco) + 617 (IVT RNA RdRp-IP4) | 12 | 12 | 100 | 32 (0.2) |
| 44 (IVT RNA E-Sarbeco) + 124 (IVT RNA RdRp-IP4) | 12 | 12 | 100 | 34.4 (0.3) |
| 9 (IVT RNA E-Sarbeco) + 25 (IVT RNA RdRp-IP4) | 12 | 12 | 100 | 36.6 (0.6) |
| 2 (IVT RNA E-Sarbeco) + 5 (IVT RNA RdRp-IP4) | 12 | 9 | 75 | 38.5 (0.6) |
| 0 (IVT RNA E-Sarbeco) + 1 (IVT RNA RdRp-IP4) | 12 | 2 | 17 | 39.3 (0.1) |
| 0 (IVT RNA E-Sarbeco) + 0 (IVT RNA RdRp-IP4) | 12 | 0 | 0 | - |
| 0 (IVT RNA E-Sarbeco) + 0 (IVT RNA RdRp-IP4) | 12 | 0 | 0 | - |
| Sample ID | Onset to Sample Collection (Days) | E-Sarbeco Assay (Ct Value) | RdRp-IP4 Assay (Ct Value) | Duo SARS-CoV-2 Assay (Ct Value) | VNT Titre | |||
|---|---|---|---|---|---|---|---|---|
| #1 | 16 | Positive | 35.2 | Positive | 34.9 | Positive | 35.2 | 80 |
| #2 | 9 | Positive | 31.4 | Positive | 31.2 | Positive | 31.2 | >160 |
| #3 | 11 | Negative | >40 | Negative | >40 | Negative | >40 | <20 |
| #4 | 13 | Negative | >40 | Negative | >40 | Negative | >40 | 80 |
| #5 | 13 | Positive | 31 | Positive | 30.4 | Positive | 30.8 | 80 |
| #6 | 9 | Positive | 31 | Positive | 30.8 | Positive | 31 | 80 |
| #7 | 17 | Negative | >40 | Negative | >40 | Positive | 38.1 | >160 |
| #8 | 15 | Negative | >40 | Negative | >40 | Positive | 37.2 | >160 |
| #9 | 12 | Positive | 35.4 | Positive | 37.4 | Positive | 35.8 | >160 |
| #10 | 16 | Positive | 32.8 | Positive | 34.2 | Positive | 33.1 | 40 |
| #11 | 13 | Positive | 34 | Positive | 35.8 | Positive | 34.4 | 40 |
| #12 | 15 | Positive | 37.1 | Positive | 36.4 | Positive | 35.9 | 40 |
| #13 | 13 | Positive | 21.5 | Positive | 22.5 | Positive | 21.9 | >160 |
| #14 | 14 | Positive | 36.2 | Positive | 34.7 | Positive | 34.8 | <20 |
| #15 | 7 | Negative | >40 | Positive | 35.2 | Positive | 36.8 | >160 |
| #16 | 9 | Negative | >40 | Positive | 36.6 | Positive | 38.9 | 80 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pezzi, L.; Charrel, R.N.; Ninove, L.; Nougairede, A.; Molle, G.; Coutard, B.; Durand, G.; Leparc-Goffart, I.; de Lamballerie, X.; Thirion, L. Development and Evaluation of a duo SARS-CoV-2 RT-qPCR Assay Combining Two Assays Approved by the World Health Organization Targeting the Envelope and the RNA-Dependant RNA Polymerase (RdRp) Coding Regions. Viruses 2020, 12, 686. https://doi.org/10.3390/v12060686
Pezzi L, Charrel RN, Ninove L, Nougairede A, Molle G, Coutard B, Durand G, Leparc-Goffart I, de Lamballerie X, Thirion L. Development and Evaluation of a duo SARS-CoV-2 RT-qPCR Assay Combining Two Assays Approved by the World Health Organization Targeting the Envelope and the RNA-Dependant RNA Polymerase (RdRp) Coding Regions. Viruses. 2020; 12(6):686. https://doi.org/10.3390/v12060686
Chicago/Turabian StylePezzi, Laura, Remi N. Charrel, Laetitia Ninove, Antoine Nougairede, Gregory Molle, Bruno Coutard, Guillaume Durand, Isabelle Leparc-Goffart, Xavier de Lamballerie, and Laurence Thirion. 2020. "Development and Evaluation of a duo SARS-CoV-2 RT-qPCR Assay Combining Two Assays Approved by the World Health Organization Targeting the Envelope and the RNA-Dependant RNA Polymerase (RdRp) Coding Regions" Viruses 12, no. 6: 686. https://doi.org/10.3390/v12060686
APA StylePezzi, L., Charrel, R. N., Ninove, L., Nougairede, A., Molle, G., Coutard, B., Durand, G., Leparc-Goffart, I., de Lamballerie, X., & Thirion, L. (2020). Development and Evaluation of a duo SARS-CoV-2 RT-qPCR Assay Combining Two Assays Approved by the World Health Organization Targeting the Envelope and the RNA-Dependant RNA Polymerase (RdRp) Coding Regions. Viruses, 12(6), 686. https://doi.org/10.3390/v12060686