Identification of Novel Astroviruses in the Gastrointestinal Tract of Domestic Cats


 , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Sample Collection
2.3. Viral Nucleic Acid and Total RNA Isolation and Sequencing
2.4. Genome Assembly and Read Mapping
2.5. Evolutionary Analysis
3. Results
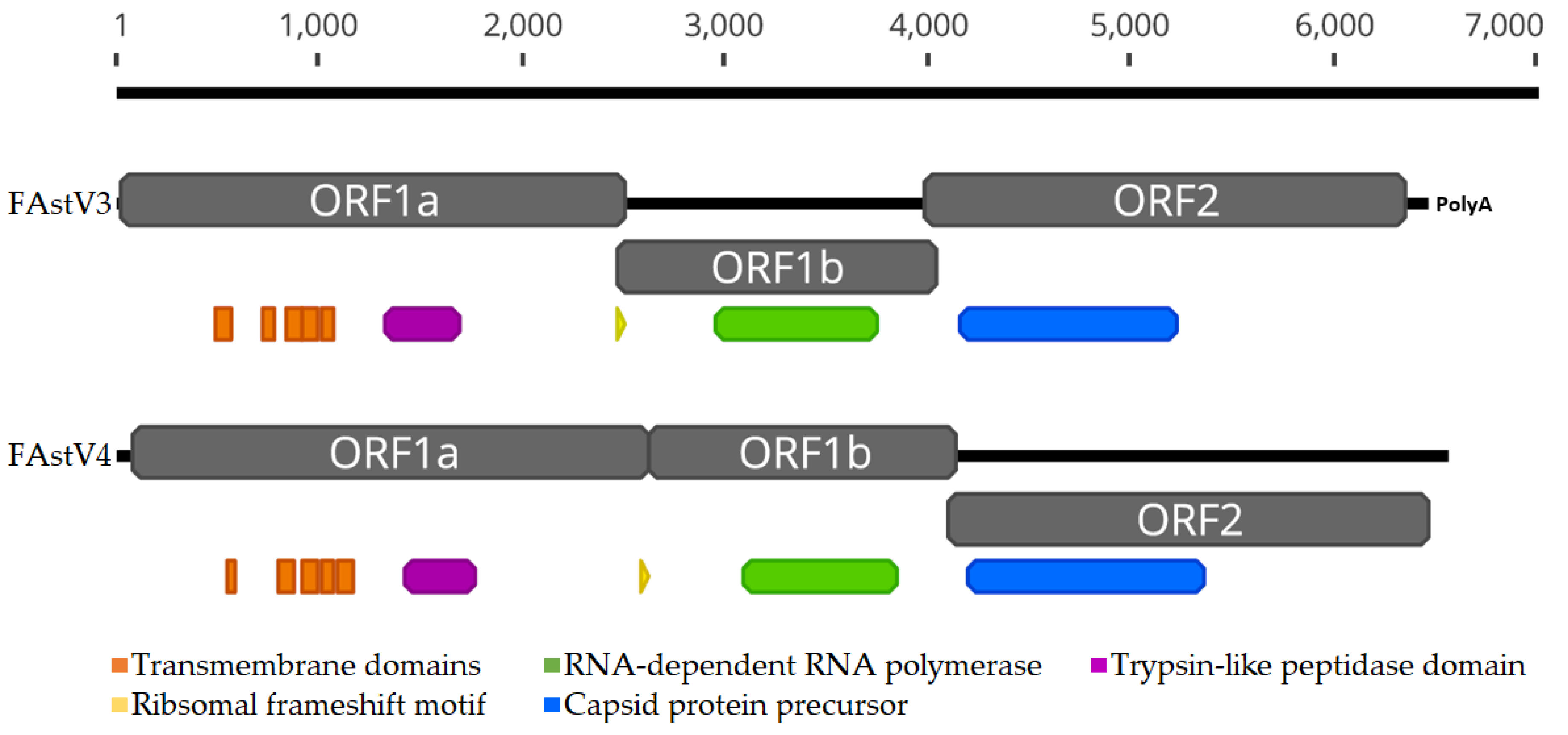
3.1. Genome Features of Novel FAstVs
3.2. Abundance, Sequence Comparison and Other Viruses
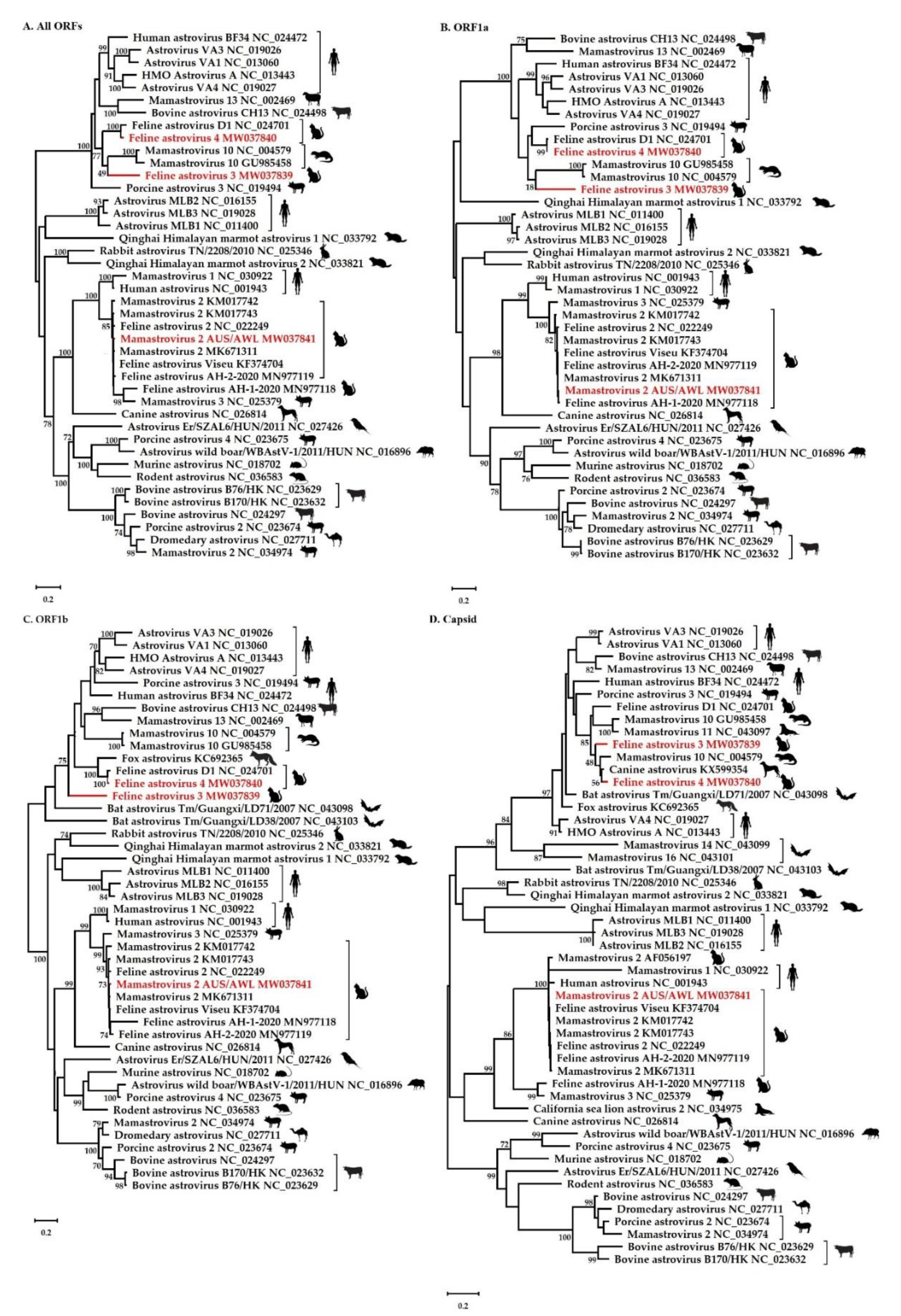
3.3. Phylogenetic and Recombination Analysis
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Guix, S.; Bosch, A.; Pintó, R.M. Astrovirus taxonomy. In Astrovirus Research: Essential Ideas, Everyday Impacts, Future Directions; Schultz-Cherry, S., Ed.; Springer: New York, NY, USA, 2013; pp. 97–118. [Google Scholar]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infection, genetics and evolution. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Pintó, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed]
- Koukou, G.; Niendorf, S.; Hornei, B.; Schlump, J.U.; Jenke, A.C.; Jacobsen, S. Human astrovirus infection associated with encephalitis in an immunocompetent child: A case report. J. Med. Case Rep. 2019, 13, 341. [Google Scholar] [CrossRef] [PubMed]
- Quan, P.-L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Mor, S.; Abin, M.; Costa, G.; Durrani, A.; Jindal, N.; Goyal, S.; Patnayak, D. The role of type-2 turkey astrovirus in poult enteritis syndrome. Poult. Sci. 2011, 90, 2747–2752. [Google Scholar] [CrossRef]
- Gough, R.E.; Borland, E.D.; Keymer, I.F.; Stuart, J.C. An outbreak of duck hepatitis type II in commercial ducks. Avian Pathol. J. WVPA 1985, 14, 227–236. [Google Scholar] [CrossRef]
- Fu, Y.; Pan, M.; Wang, X.; Xu, Y.; Xie, X.; Knowles, N.J.; Yang, H.; Zhang, D. Complete sequence of a duck astrovirus associated with fatal hepatitis in ducklings. J. Gen. Virol. 2009, 90, 1104–1108. [Google Scholar] [CrossRef]
- Imada, T.; Yamaguchi, S.; Mase, M.; Tsukamoto, K.; Kubo, M.; Morooka, A. Avian Nephritis Virus (ANV) as a new member of the family Astroviridae and Construction of infectious ANV cDNA. J. Virol. 2000, 74, 8487–8493. [Google Scholar] [CrossRef]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef]
- Blomström, A.-L.; Widén, F.; Hammer, A.-S.; Belák, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef]
- Gavier-Widén, D.; Bröjer, C.; Dietz, H.H.; Englund, L.; Hammer, A.S.; Hedlund, K.-O.; Hård af Segerstad, C.; Nilsson, K.; Nowotny, N.; Puurula, V.; et al. Investigations into shaking mink syndrome: An encephalomyelitis of unknown cause in farmed mink (Mustela Vison) kits in Scandinavia. J. Vet. Diagn. Investig. 2004, 16, 305–312. [Google Scholar]
- Hoshino, Y.; Zimmer, J.F.; Moise, N.S.; Scott, F.W. Detection of astroviruses in feces of a cat with diarrhea. Arch. Virol. 1981, 70, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.F.F.; Mesquita, J.R.; Nascimento, M.S.J.; Kondov, N.O.; Wong, W.; Reuter, G.; Knowles, N.J.; Vega, E.; Esona, M.D.; Deng, X.; et al. Feline fecal virome reveals novel and prevalent enteric viruses. Vet. Microbiol. 2014, 171, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Woo, P.C.Y.; Yip, C.C.Y.; Bai, R.; Wu, Y.; Tse, H.; Yuen, K.-Y. Complete genome sequence of a novel feline astrovirus from a domestic cat in Hong Kong. Genome Announc. 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, L.; Deng, X.; Kapusinszky, B.; Pesavento, P.A.; Delwart, E. Faecal virome of cats in an animal shelter. J. Gen. Virol. 2014, 95, 2553–2564. [Google Scholar] [CrossRef]
- Moschidou, P.; Martella, V.; Lorusso, E.; Desario, C.; Pinto, P.; Losurdo, M.; Catella, C.; Parisi, A.; Bányai, K.; Buonavoglia, C. Mixed infection by Feline astrovirus and Feline panleukopenia virus in a domestic cat with gastroenteritis and panleukopenia. J. Vet. Diagn. Investig. 2011, 23, 581–584. [Google Scholar] [CrossRef]
- Marshall, J.A.; Kennett, M.L.; Rodger, S.M.; Studdert, M.J.; Thompson, W.L.; Gust, I.D. Virus and virus-like particles in the faeces of cats with and without diarrhoea. Aust. Vet. J. 1987, 64, 100–105. [Google Scholar] [CrossRef]
- Lawler, P.E.; Cook, K.A.; Williams, H.G.; Archer, L.L.; Schaedel, K.E.; Isaza, N.M.; Wellehan, J.J.F.X. Determination of the diversity of astroviruses in feces from cats in Florida. J. Vet. Diagn. Investig. 2018, 30, 275–279. [Google Scholar] [CrossRef]
- Yi, S.; Niu, J.; Wang, H.; Dong, G.; Guo, Y.; Dong, H.; Wang, K.; Hu, G. Molecular characterization of feline astrovirus in domestic cats from Northeast China. PLoS ONE 2018, 13, e0205441. [Google Scholar] [CrossRef]
- Van Brussel, K.; Carrai, M.; Lin, C.; Kelman, M.; Setyo, L.; Aberdein, D.; Brailey, J.; Lawler, M.; Maher, S.; Plaganyi, I.; et al. Distinct lineages of Feline Parvovirus associated with Epizootic outbreaks in Australia, New Zealand and the United Arab Emirates. Viruses 2019, 11, 1155. [Google Scholar] [CrossRef]
- Conceição-Neto, N.; Zeller, M.; Lefrère, H.; De Bruyn, P.; Beller, L.; Deboutte, W.; Yinda, C.K.; Lavigne, R.; Maes, P.; Van Ranst, M.; et al. Modular approach to customise sample preparation procedures for viral metagenomics: A reproducible protocol for virome analysis. Sci. Rep. 2015, 5, 16532. [Google Scholar] [CrossRef] [PubMed]
- Chong, R.; Shi, M.; Grueber, C.E.; Holmes, E.C.; Hogg, C.J.; Belov, K.; Barrs, V.R. Fecal viral diversity of captive and wild Tasmanian Devils characterized using Virion-Enriched metagenomics and metatranscriptomics. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Kopylova, E.; Noé, L.; Touzet, H. SortMeRNA: Fast and accurate filtering of ribosomal RNAs in metatranscriptomic data. Bioinformatics 2012, 28, 3211–3217. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Rettedal, E.A.; van der Helm, E.; Ellabaan, M.; Panagiotou, G.; Sommer, M.O.A. Antibiotic treatment drives the diversification of the human gut resistome. Genom. Proteom. Bioinform. 2019, 17, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Li, J.; Ni, Y.; Kang, K.; Misiakou, M.-A.; Imamovic, L.; Chow, B.K.C.; Rode, A.A.; Bytzer, P.; Sommer, M.; et al. Mining, analyzing, and integrating viral signals from metagenomic data. Microbiome 2019, 7, 42. [Google Scholar] [CrossRef]
- Peng, Y.; Leung, H.C.; You, S.M.; Chin, F.Y. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Hordijk, W.; Gascuel, O. Improving the efficiency of SPR moves in phylogenetic tree search methods based on maximum likelihood. Bioinformatics 2005, 21, 4338–4347. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1. [Google Scholar] [CrossRef]
- Zhang, Q.; Niu, J.; Yi, S.; Dong, G.; Yu, D.; Guo, Y.; Huang, H.; Hu, G. Development and application of a multiplex PCR method for the simultaneous detection and differentiation of feline panleukopenia virus, feline bocavirus, and feline astrovirus. Arch. Virol. 2019, 164, 2761–2768. [Google Scholar] [CrossRef]
- Harbour, D.A.; Ashley, C.R.; Williams, P.D.; Gruffydd-Jones, T.J. Natural and experimental astrovirus infection of cats. Vet. Rec. 1987, 120, 555–557. [Google Scholar] [CrossRef]
- Aydın, H.; Timurkan, M.O. A pilot study on feline astrovirus and feline panleukopenia virus in shelter cats in Erzurum, Turkey. Rev. Méd. Vét. 2018, 169, 52–57. [Google Scholar]
- Cho, Y.-Y.; Lim, S.-I.; Kim, Y.K.; Song, J.-Y.; Lee, J.-B.; An, D.-J. Molecular characterisation and phylogenetic analysis of feline astrovirus in Korean cats. J. Feline Med. Surg. 2013, 16, 679–683. [Google Scholar] [CrossRef]
- Menon, V.K.; George, S.; Sarkar, R.; Giri, S.; Samuel, P.; Vivek, R.; Saravanabavan, A.; Liakath, F.B.; Ramani, S.; Iturriza-Gomara, M.; et al. Norovirus Gastroenteritis in a Birth Cohort in Southern India. PLoS ONE 2016, 11, e0157007. [Google Scholar] [CrossRef]
- Klein-Richers, U.; Hartmann, K.; Hofmann-Lehmann, R.; Unterer, S.; Bergmann, M.; Rieger, A.; Leutenegger, C.M.; Pantchev, N.; Balzer, J.; Felten, S. Prevalence of Feline Coronavirus shedding in German catteries and associated risk factors. Viruses 2020, 12, 1000. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.E.; Briggs, J.; Guerrero, M.L.; Matson, D.O.; Pickering, L.K.; Ruiz-Palacios, G.; Berke, T.; Mitchell, D.K. Molecular characterization of a novel recombinant strain of human astrovirus associated with gastroenteritis in children. Arch. Virol. 2001, 146, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- Wolfaardt, M.; Kiulia, N.M.; Mwenda, J.M.; Taylor, M.B. Evidence of a recombinant wild-type human astrovirus strain from a Kenyan child with Gastroenteritis. J. Clin. Microbiol. 2011, 49, 728–731. [Google Scholar] [CrossRef]
- Rivera, R.; Nollens, H.H.; Venn-Watson, S.; Gulland, F.M.; Wellehan, J.F., Jr. Characterization of phylogenetically diverse astroviruses of marine mammals. J. Gen. Virol. 2010, 91, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Kuroda, M.; Masuda, T.; Akagami, M.; Haga, K.; Tsuchiaka, S.; Kishimoto, M.; Naoi, Y.; Sano, K.; Omatsu, T.; et al. Whole genome analysis of porcine astroviruses detected in Japanese pigs reveals genetic diversity and possible intra-genotypic recombination. Infect. Genet. Evol. 2017, 50, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Pérot, P.; Lecuit, M.; Eloit, M. Astrovirus diagnostics. Viruses 2017, 9, 10. [Google Scholar]



{kind=link}
{kind=link}
| Virus | Accession Number | Assembler | Library Virus Was Identified | Library Read Count after Processing (Paired End) | Virus/Contig Read Count | Transcripts per Million |
|---|---|---|---|---|---|---|
| Feline astrovirus 3 | MW037839 | IDBA-UD | AWL4 | 7,030,294 | 62,663 | 1623 |
| AWL6 | 10,116,484 | 240,905 | 3213 | |||
| AWL8 | 10,578,534 | 222 | 32 | |||
| 279 | 42 | |||||
| Feline astrovirus 4 | MW037840 | Trinity | 159 | 77,205,956 | 32,003 | 16 |
| Mamastrovirus 2 AUS/AWL | MW037841 | IDBA-UD | AWL4 | 7,030,294 | 1636 | 70 |
| 1897 | 114 | |||||
| AWL6 | 10,116,484 | 4098 | 53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Brussel, K.; Wang, X.; Shi, M.; Carrai, M.; Li, J.; Martella, V.; Beatty, J.A.; Holmes, E.C.; Barrs, V.R. Identification of Novel Astroviruses in the Gastrointestinal Tract of Domestic Cats. Viruses 2020, 12, 1301. https://doi.org/10.3390/v12111301
Van Brussel K, Wang X, Shi M, Carrai M, Li J, Martella V, Beatty JA, Holmes EC, Barrs VR. Identification of Novel Astroviruses in the Gastrointestinal Tract of Domestic Cats. Viruses. 2020; 12(11):1301. https://doi.org/10.3390/v12111301
Chicago/Turabian StyleVan Brussel, Kate, Xiuwan Wang, Mang Shi, Maura Carrai, Jun Li, Vito Martella, Julia A. Beatty, Edward C. Holmes, and Vanessa R. Barrs. 2020. "Identification of Novel Astroviruses in the Gastrointestinal Tract of Domestic Cats" Viruses 12, no. 11: 1301. https://doi.org/10.3390/v12111301
APA StyleVan Brussel, K., Wang, X., Shi, M., Carrai, M., Li, J., Martella, V., Beatty, J. A., Holmes, E. C., & Barrs, V. R. (2020). Identification of Novel Astroviruses in the Gastrointestinal Tract of Domestic Cats. Viruses, 12(11), 1301. https://doi.org/10.3390/v12111301