Association of Genetic Polymorphisms in TLR3, TLR4, TLR7, and TLR8 with the Clinical Forms of Dengue in Patients from Veracruz, Mexico

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Blood Samples
2.3. Hematological Values
2.4. Serotype-Specific Detection of Dengue Viruses
2.5. Anti-Dengue IgM and IgG Capture ELISA
2.6. Anti-Dengue IgG Subclasses EIA
2.7. Genotyping of Polymorphisms in TLR3, TLR4, TLR7, and TLR8
2.8. Calculations and Statistical Analysis
3. Results
3.1. Hematological Values of DHF and DF Patients
3.2. Identification of Viral Serotypes in Patients with DF and DHF
3.3. IgM, Total IgG, and IgG Subclasses in Patients with DF and DHF
3.4. Association of Genetic Polymorphisms in TLR3, TLR7, and TLR8 with the Clinical Forms of Dengue Virus Infection
3.5. Association of Genetic Polymorphisms in the TLR4 Gene with the Clinical Forms of Dengue Virus Infection
3.6. Linkage Disequilibrium Analysis for TLR4 SNPs
3.7. Haplotype Analyses in TLR4 SNPs
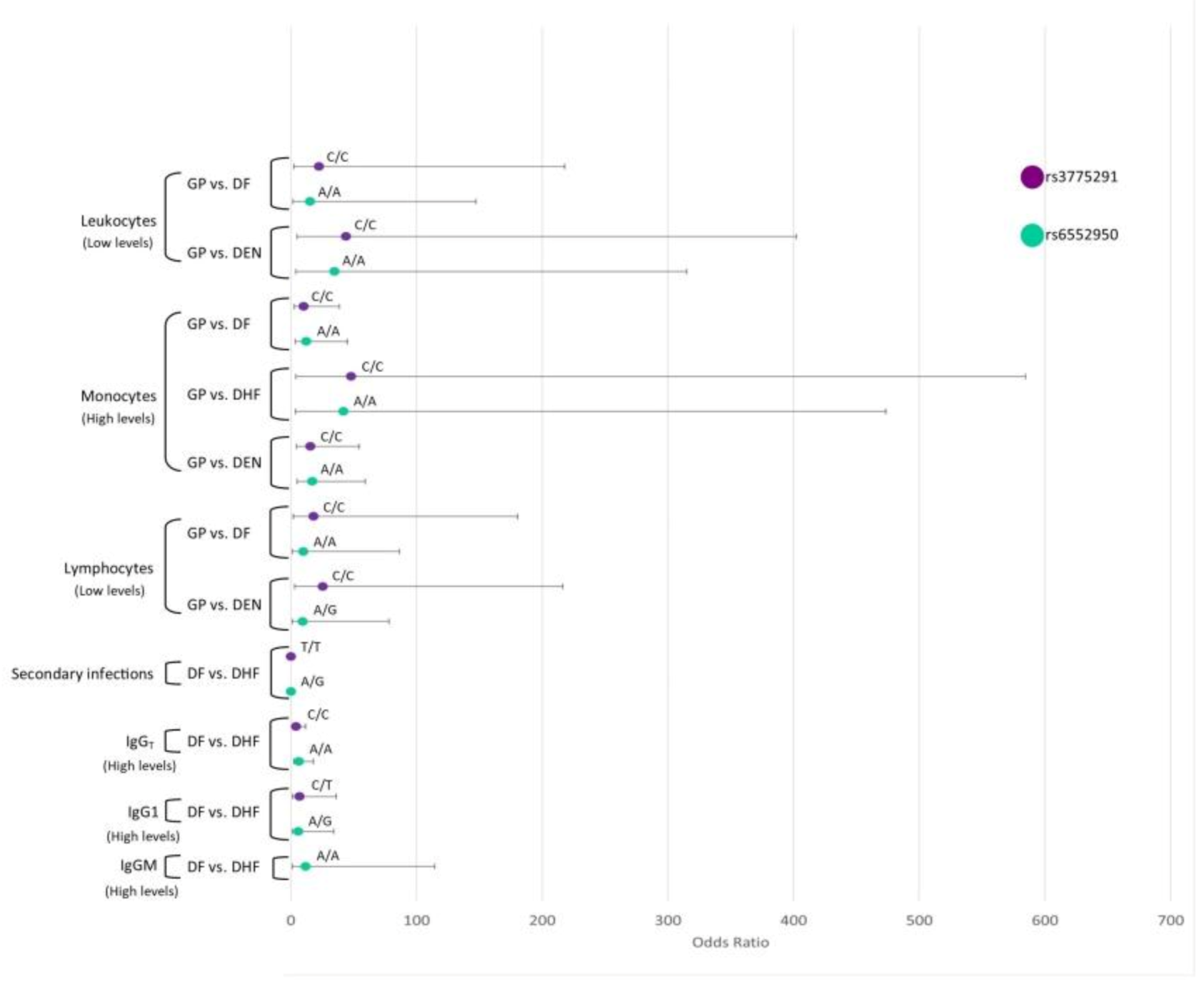
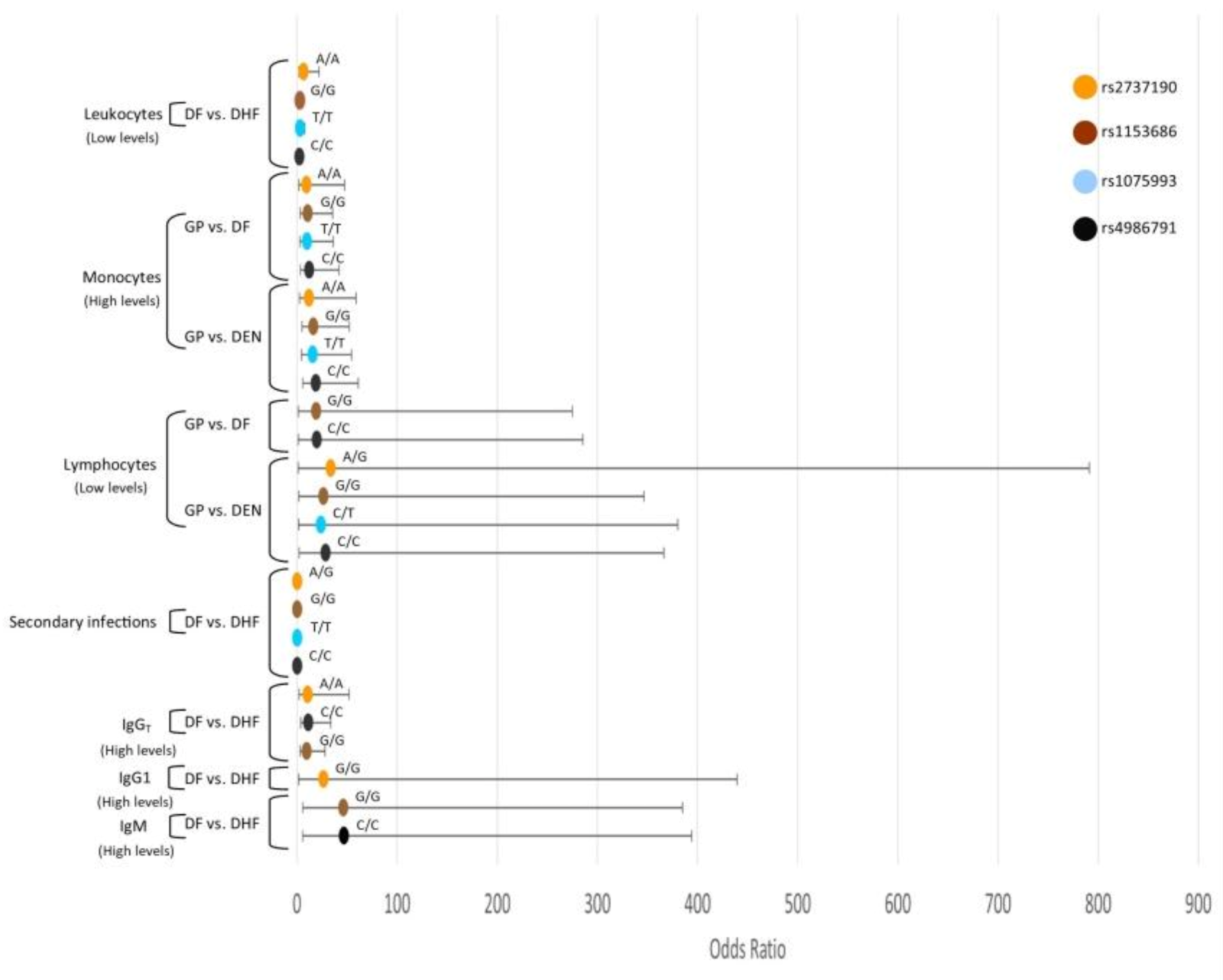
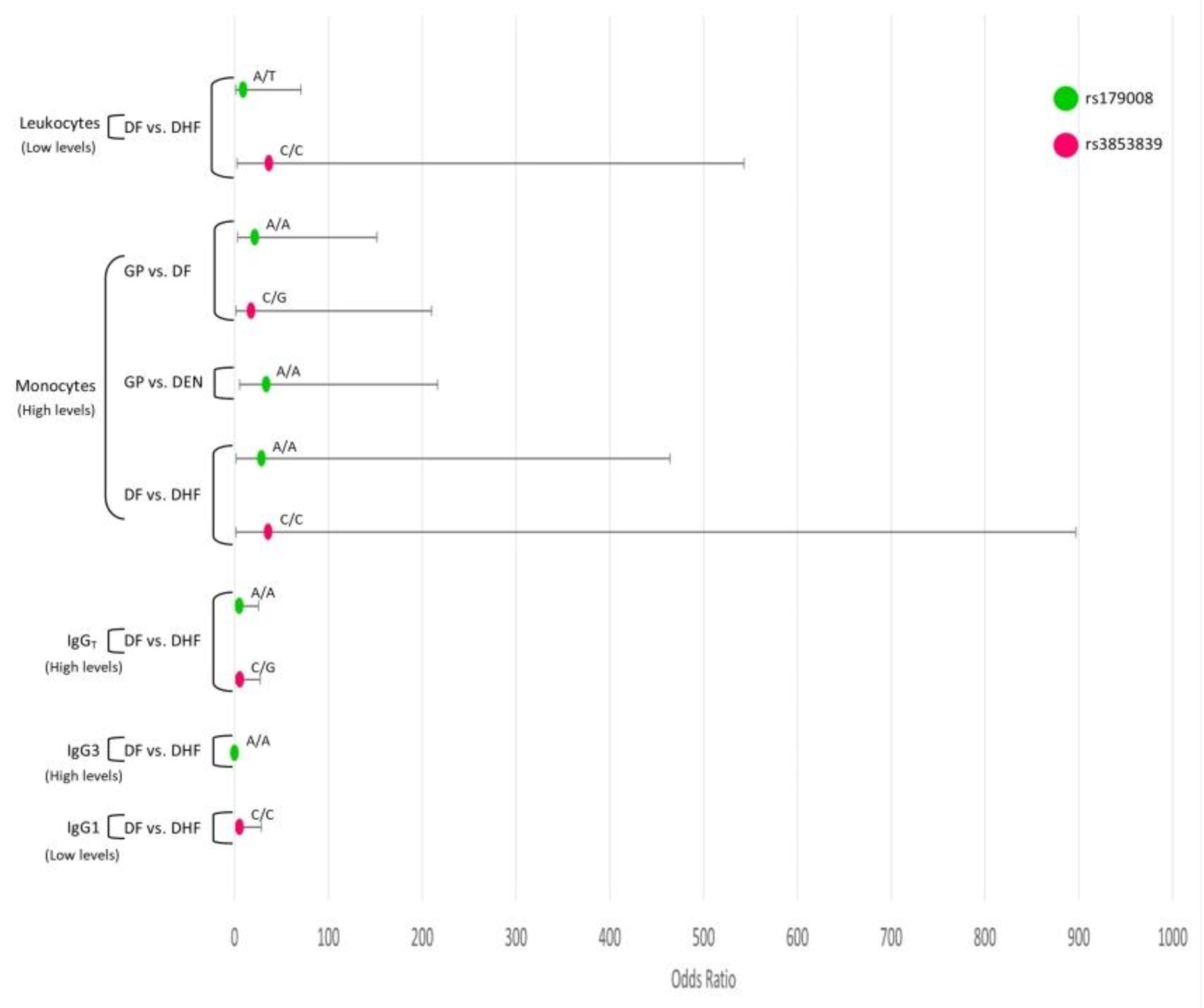
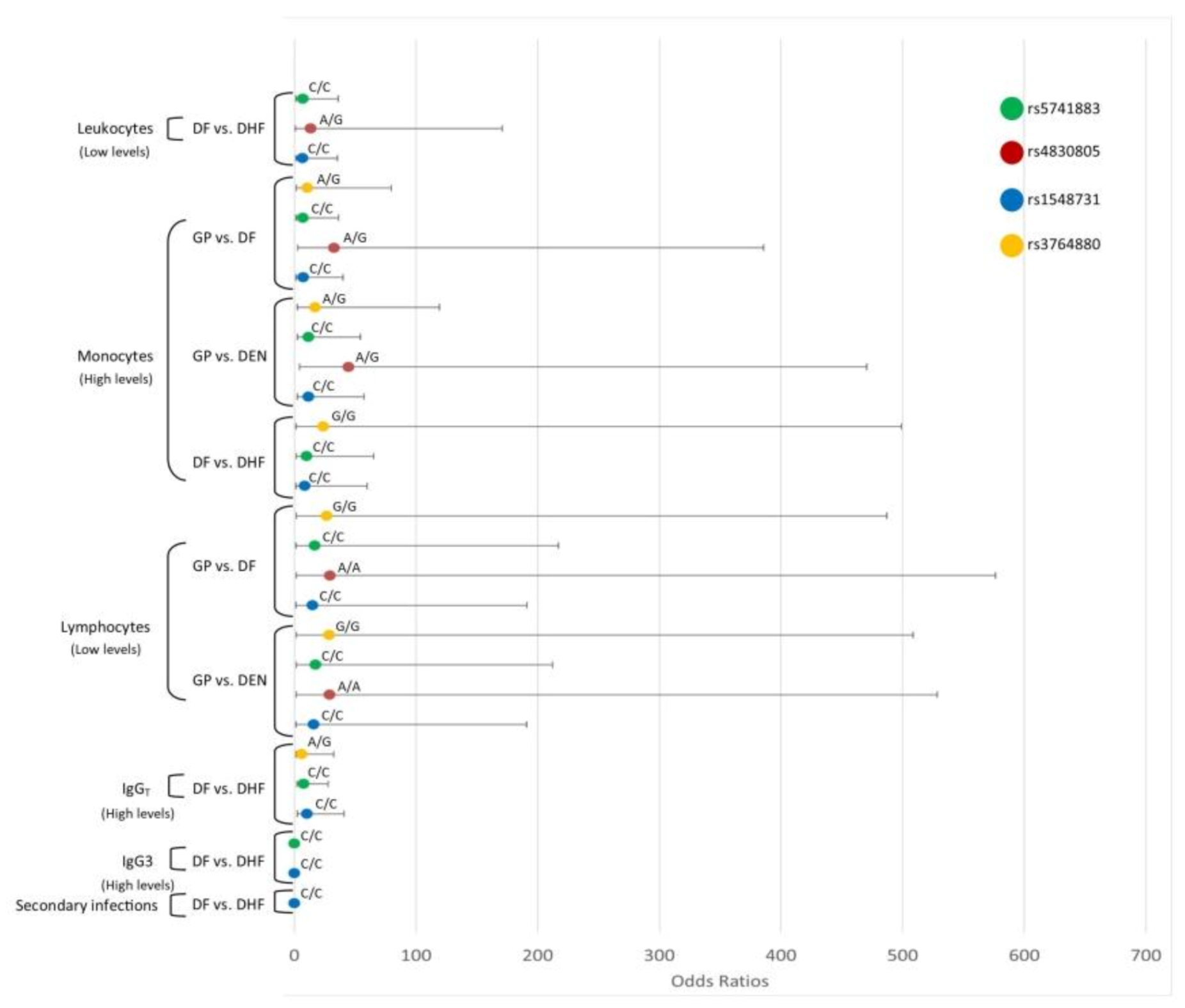
3.8. Interaction Analysis of TLR SNPs and the Hematological Covariates
3.8.1. TLR3
3.8.2. TLR4
3.8.3. TLR7
3.8.4. TLR8
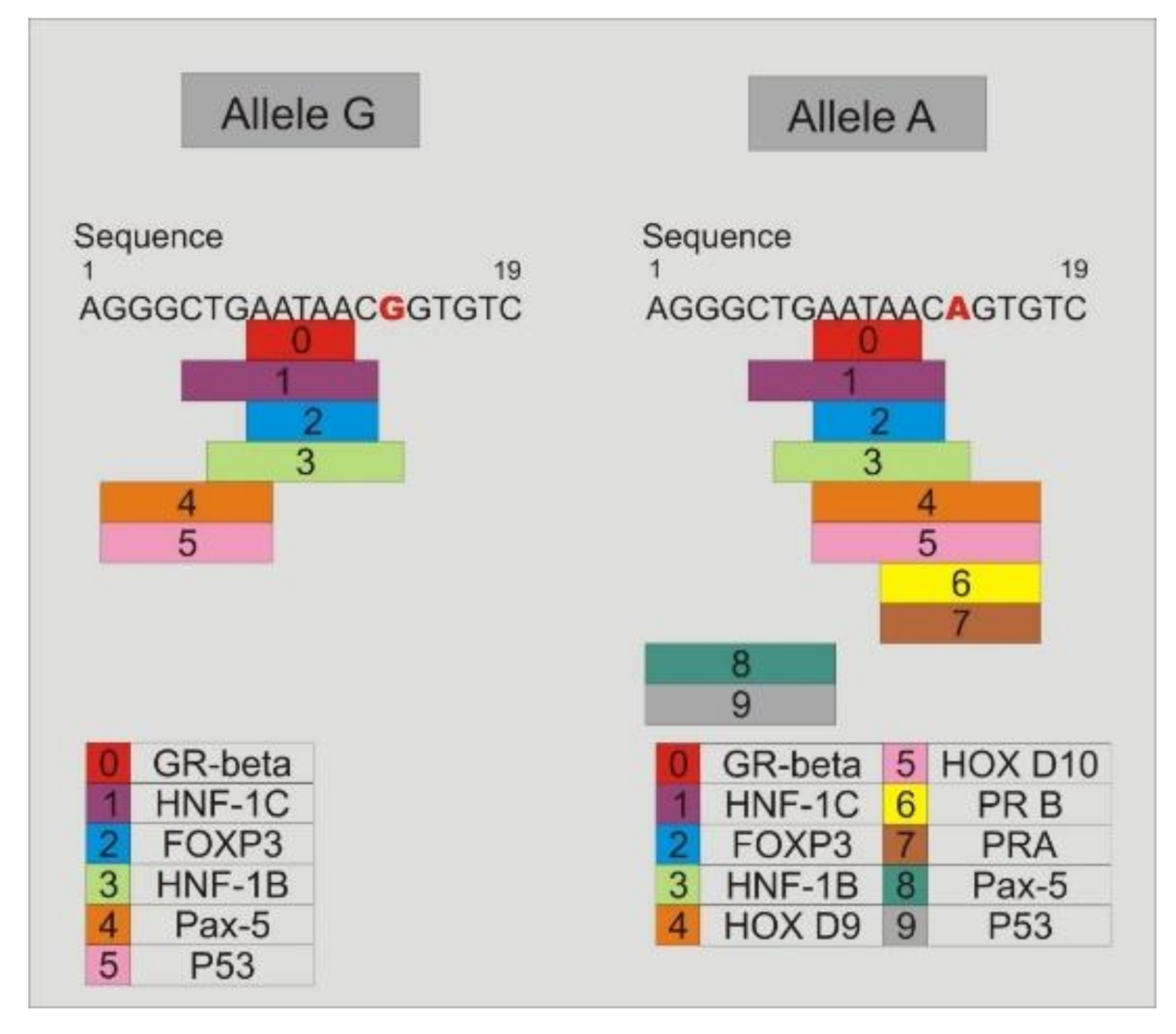
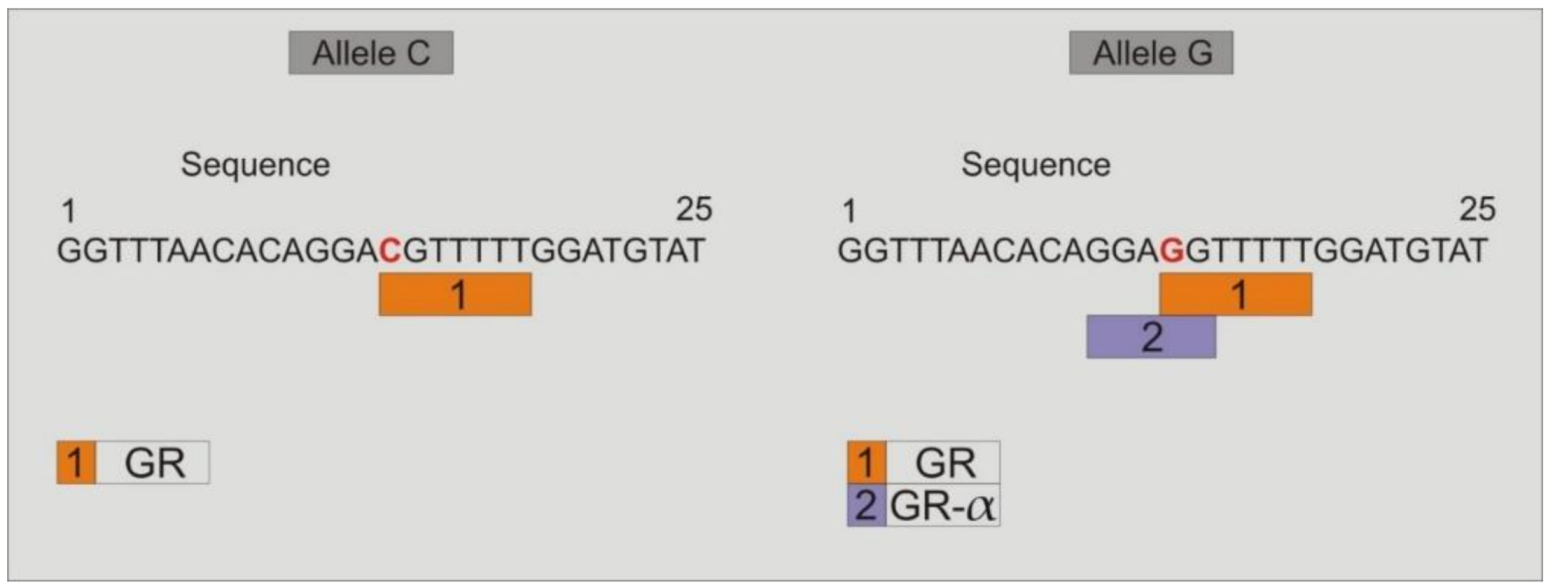
3.9. Study of the Binding of Transcription Factors to the Promoter Region of TLR4-rs2737190 and TLR4-rs11536865
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- De La Cruz, H.S.; Flores, A.H.; González, M.S.; López, M.I.; Alpuche, A.C.; Ludert, J.E.; Del Angel, R.M. Determination of viremia and concentration of circulating nonstructural protein 1 in patients infected with dengue virus in Mexico. Am. J. Trop. Med. Hyg. 2013, 88, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.D.; Mohan, P.M.; Saxena, P.; Kumar, M.; Rai, A.; Tazeen, P.S.; Mukul, J.A. Emergence and Continued Circulation of Dengue-2 (Genotype IV) Virus Strains in Northern India. J. Med. Virol. 2004, 74, 314–322. [Google Scholar]
- Fernández, M.M.T.; Gendzekhadze, K.; Rivas, V.P.; Layrisse, Z. TNF-α-308A allele, a possible severity risk factor of hemorrhagic manifestation in dengue fever patients. Tissue Antigens 2004, 64, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Yacoub, S.; Mongkolsapaya, J.; Screaton, G. The pathogenesis of dengue. Curr. Opin. Infect. Dis. 2013, 26, 284–289. [Google Scholar] [CrossRef]
- Rodrigues, C.F.N.; Ribeiro, N.R.M.; Bispo, F.A.M.; Bastos, S.S.J.; Bruycker, N.F.; Rocha, Q.L.M.; Barreto, S.F. Twenty Years of DENV-2 Activity in Brazil: Molecular Characterization and Phylogeny of Strains Isolated from 1990 to 2010. PLoS Negl. Trop. Dis. 2013, 7, e2095. [Google Scholar]
- Satoh, T.; Akira, S. Toll-like receptors: Critical proteins. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Matsumoto, M.; Funami, K.; Oshiumi, H.; Seya, T. Toll-Like Receptor 3: A Link between Toll-Like Receptor, Interferon and Viruses. Microbiol. Immunol. 2004, 48, 147–154. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF- k B by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Takeda, K.; Kaisho, T.; Akira, S. Toll -like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Reis, S.C. Innate Antiviral Responses by Means of TLR7-Mediated Recognition of Single-Stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar] [CrossRef] [PubMed]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-Specific Recognition of Single-Stranded RNA via Toll-like Receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [PubMed]
- Modhiran, N.; Watterson, D.; Muller, D.A.; Panetta, A.K.; Sester, D.P.; Liu, L.; Hume, D.A.; Stacey, K.J.; Young, P.R. Dengue virus NS1 protein activates cells via Toll-like receptor 4 and disrupts endothelial cell monolayer integrity. Sci. Transl. Med. 2015, 7, 304ra142. [Google Scholar] [CrossRef] [PubMed]
- Misch, E.A.; Hawn, T.R. Toll-like receptor polymorphisms and susceptibility to human disease. Clin. Sci. 2008, 114, 347–360. [Google Scholar] [CrossRef]
- Tapping, R.I. Innate immune sensing and activation of cell surface Toll-like receptors. Semin. Immunol. 2009, 21, 175–184. [Google Scholar] [CrossRef]
- Medvedev, A.E. Toll-Like Receptor Polymorphisms, Inflammatory and Infectious Diseases, Allergies, and Cancer. J. Interferon Cytokine Res. 2013, 33, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Djamiatun, K.; Ferwerda, B.; Netea, M.G.; Ven, A.J.A.M.; Dolmans, W.M.V.; Faradz, S.M.H. Short Report: Toll-Like Receptor 4 Polymorphisms in Dengue Virus—Infected Children. Am. J. Trop. Med. Hyg. 2011, 85, 352–354. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, S.K.; Kakkar, K.; Nyari, N.; Dhole, T.N.; Kashyap, R.; Hasan, S. Analysis of TLR4 (Asp299Gly and Thr399Ile) gene polymorphisms and mRNA level in patients with dengue infection: A case-control study. Infect. Genet. Evol. 2016, 43, 412–417. [Google Scholar] [CrossRef]
- Alagarasu, K.; Bachal, R.V.; Memane, R.S.; Shah, P.S.; Cecilia, D. Immunobiology Polymorphisms in RNA sensing toll like receptor genes and its association with clinical outcomes of dengue virus infection. Immunobiology 2015, 220, 164–168. [Google Scholar] [CrossRef]
- World Health Organization. Dengue Haemorrhagic Fever: Diagnosis, Treatment, Prevention and Control, 2nd ed.; World Health Organization: Geneva, Switzerland, 1997. [Google Scholar]
- Chien, L.J.; Liao, T.L.; Shu, P.Y.; Huang, J.H.; Gubler, D.J.; Chang, G.J. Development of real-time reverse transcriptase PCR assays to detect and serotype dengue viruses. J. Clin. Microbiol. 2006, 44, 1295–1304. [Google Scholar] [CrossRef]
- World Health Organization. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control, new ed.; WHO/HTM/NTD/DEN/2009.1; WHO: Geneva, Switzerland, 2009. [Google Scholar]
- Messeguer, X.; Escudero, R.; Farré, D.; Núñez, O.; Martínez, J.; Albà, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef] [PubMed]
- Farré, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Roselló, L.; Albà, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef] [PubMed]
- Posadas, M.A.; Aguilar, F.J.L.; Chávez, N.A.; Guillén, S.E.; Alcántara, F.V.; Luna, R.L.; Ávila, T.A.M.; Pacheco, Y.J.C. Indices of anti-dengue immunoglobulin G subclasses in adult Mexican patients with febrile and hemorrhagic dengue in the acute phase. Microbiol. Immunol. 2017, 61, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Huik, K.; Avi, R.; Pauskar, M.; Kallas, E.; Jõgeda, E.L.; Karki, T.; Marsh, K.; Des, J.D.; Uusküla, A.; Lutsar, I. Association between TLR3 rs3775291 and resistance to HIV among highly exposed Caucasian intravenous drug users. Infect. Genet. Evol. 2013, 20, 78–82. [Google Scholar] [CrossRef]
- Citores, M.J.; Pérez, P.S.; Duca, A.; Crespo, G.; Fuente, S.; Vilches, C.; Navasa, M.; Cuervas, M.V. Rapidity of fibrosis progression in liver transplant recipients with recurrent hepatitis C is influenced by toll-like receptor 3 polymorphism. Clin. Transplant. 2016, 30, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Studzinska, M.; Jabłonska, A.; Wisniewska, L.M.; Nowakowska, D.; Gaj, Z.; Lesnikowski, Z.J.; Wozniakowska, G.T.; Wilczynski, J.; Paradowska, E. Association of TLR3 L412F Polymorphism with Cytomegalovirus Infection in Children. PLoS ONE 2017, 12, e0169420. [Google Scholar] [CrossRef]
- Her, Z.; Teng, T.S.; Tan, J.J.L.; Teo, T.H.; Kam, Y.W.; Lum, F.M.; Lee, W.W.L.; Gabriel, C.; Melchiotti, R.; Andiappan, A.K.; et al. Loss of TLR 3 aggravates CHIKV replication and pathology due to an altered virus-specific neutralizing antibody response. EMBO Mol. Med. 2015, 7, 24–41. [Google Scholar] [CrossRef]
- Mukherjee, S.; Tripathi, A. Contribution of Toll like receptor polymorphisms to dengue susceptibility and clinical outcome among eastern Indian patients. Immunobiology 2019, 224, 774–785. [Google Scholar] [CrossRef]
- Kumar, D.S.; Tripathi, A. Association of toll-like receptor polymorphisms with susceptibility to chikungunya virus infection. Virology 2017, 511, 207–213. [Google Scholar]
- Kurt, J.E.A.; Popova, L.; Kwinn, L.; Haynes, L.M.; Jones, L.P.; Tripp, R.A.; Walsh, E.E.; Freeman, M.W.; Golenbock, D.T.; Anderson, L.J.; et al. Pattern recognition receptors TLR4 and CD14 mediate response to respiratory syncytial virus. Nat. Immunol. 2000, 1, 398–401. [Google Scholar] [CrossRef]
- Haynes, L.M.; Moore, D.D.; Kurt, J.E.A.; Finberg, R.W.; Anderson, L.J.; Tripp, R.A. Involvement of Toll-Like Receptor 4 in Innate Immunity to Respiratory Syncytial Virus. J. Virol. 2001, 75, 10730–10737. [Google Scholar] [CrossRef] [PubMed]
- Okumura, A.; Pitha, P.M.; Yoshimura, A.; Harty, R.N. Interaction between Ebola Virus Glycoprotein and Host Toll-Like Receptor 4 Leads to Induction of Proinflammatory Cytokines and SOCS1. J. Virol. 2010, 84, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Burzyn, D.; Rassa, J.C.; Kim, D.; Nepomnaschy, I.; Ross, S.R.; Piazzon, I. Toll-Like Receptor 4-Dependent Activation of Dendritic Cells by a Retrovirus. J. Virol. 2004, 78, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Oetting, W.S.; Guan, W.; Schladt, D.P.; Leduc, R.E.; Jacobson, P.A.; Matas, A.J.; Chinnakotla, S.; Schröppel, B.; Murphy, B.T.; Israni, A.K. Donor polymorphisms of toll-like receptor 4 associated with graft failure in liver transplant recipients. Liver Transpl. 2012, 18, 1399–1405. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Cidlowski, J.A. Corticosteroids: Mechanisms of Action in Health and Disease. Rheum. Dis. Clin. N. Am. 2016, 42, 15–31. [Google Scholar] [CrossRef]
- Wang, M.; Xu, G.; Lü, L.; Xu, K.; Chen, Y.; Pan, H.; Burstrom, B.; Wang, J. Genetic polymorphisms of IL-17A, IL-17F, TLR4 and miR-146a in association with the risk of pulmonary tuberculosis. Sci. Rep. 2016, 6, 28586. [Google Scholar] [CrossRef]
- Zhou, J.G.; Zhang, J.P.; Lee, B.H.; Zhang, S.L.; Zhu, L.; Chen, C. Toll-like receptor 4 polymorphisms in gram-negative bacterial infections of han chinese neonates. Am. J. Perinatol. 2015, 32, 363–370. [Google Scholar]
- Ragupathy, V.; Xue, W.; Tan, J.; Devadas, K.; Gao, Y.; Hewlett, I. Progesterone augments cell susceptibility to HIV-1 and HIV-1/HSV-2 co-infections. J. Mol. Endocrinol. 2016, 57, 185–199. [Google Scholar] [CrossRef][Green Version]
- Bose, P.D.; Das, B.C.; Kumar, A.; Gondal, R.; Kumar, D.; Kar, P. High viral load and deregulation of the progesterone receptor signaling pathway: Association with Hepatitis E-related poor pregnancy outcome. J. Hepatol. 2011, 54, 1107–1113. [Google Scholar] [CrossRef]
- Hall, O.J.; Limjunyawong, N.; Vermillion, M.S.; Robinson, D.P.; Wohlgemuth, N.; Pekosz, A.; Mitzner, W.; Klein, S.L. Progesterone-Based Therapy Protects Against Influenza by Promoting Lung Repair and Recovery in Females. PLoS Pathog. 2016, 12, e1005840. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DF (n = 100) | DHF (n = 65) | Dengue (n = 165) | GP (n = 89) | |
|---|---|---|---|---|
| Age | 37.86 ± 13.49 a,b | 42.95 ± 14.50 a | 39.85 ± 14.07 c | 49.52 ± 22.99 b,c |
| Female/male | 49/51 | 28/37 | 77/88 | 59/30 |
| Platelets (×109/L) | 191.1 ± 65.07 a,b | 54.03 ± 30.48 a,c | 137.1 ±86.21 d | 253.7 ± 64.34 b,c,d |
| Leukocytes (×109/L) | 6.279 ± 3.618 a,b | 4.459 ± 1.964 a,c | 5.562 ± 3.194 d | 7.926 ± 2.050b c,d |
| Neutrophils (%) | 61.79 ± 16.84 a,b | 46.49 ± 16.62 a,c | 55.76 ± 18.31 | 56.76 ± 10.51 b,c |
| Lymphocytes (%) | 24.77 ± 14.38 a,b | 35.23 ± 14.88 a | 28.89 ± 15.41 | 31.24 ± 9.044 b |
| Monocytes (%) | 10.33 ± 4.705 a,b | 14.44 ± 7.11 a, c | 11.95 ± 6.095 d | 7.517 ± 2.646 b,c,d |
| Antibody | Clinical Dengue Forms | Indices | ||
|---|---|---|---|---|
| Low n (%) | High n (%) | Negative n (%) | ||
| IgM | DF | 12 (12) | 14 (14) | 74 (74) |
| DHF | 4 (6.15) | 38 (58.46) | 23 (35.38) | |
| IgG | DF | 77 (77) | 16 (16) | 7 (7) |
| DHF | 27 (41.53) | 36 (55.38) | 2 (3.077) | |
| IgG1 | DF | 56 (71.8) | 22 (28.2) | 0 |
| DHF | 36 (61.02) | 23 (38.98) | 0 | |
| IgG2 | DF | 0 | 7 (8.97) | 71 (91.03) |
| DHF | 0 | 9 (15.25) | 50 (84.75) | |
| IgG3 | DF | 21 (26.92) | 30 (38.46) | 27 (34.62) |
| DHF | 23 (38.98) | 15 (25.42) | 21 (35.59) | |
| IgG4 | DF | 54 (69.23) | 24 (30.77) | 0 |
| DHF | 35 (59.32) | 21 (35.59) | 3 (5.08) | |
| TLR4 SNP Genotype/Allele | GP (n = 89) N (%) | DF (n = 100) N (%) | DHF (n = 65) N (%) | DEN (DF + DHF) (n = 165) N (%) | DF vs. DHF | GP vs. DF | GP vs. DHF | GP vs. DEN |
|---|---|---|---|---|---|---|---|---|
| rs2737190 | ||||||||
| G Mi,An | 72 (40) | 64 (32) | 40 (31) | 104 (32) | 0.94(0.58−1.52) | 0.69 (0.45−1.05) | 0.65 (0.40−1.05) | 0.67 (0.46–0.98) |
| A | 106 (60) | 136 (68) | 90 (69) | 226 (68) | ||||
| p-value | 0.81 | 0.0875 | 0.08 | 0.043 | ||||
| A/A | 33 (37) | 42 (42) | 31 (48) | 73 (44) | 1.0 | 1.0 | 1.0 | 1.0 |
| A/G | 40 (45) | 52 (52) | 28 (43) | 80 (48) | 0.73 (0.38−1.40) | 1.02 (0.55−1.89) | 0.75 (0.37−1.48) | 0.9 (0.52−1.58) |
| G/G | 16 (18) | 6 (6) | 6 (9) | 12 (8) | 1.35 (0.40−4.60) | 0.29 (0.10−0.84) | 0.40 (0.14−1.15) | 0.34 (0.14−0.8) |
| p-value | 0.47 | 0.035 | 0.21 | 0.038 | ||||
| HWE (p-value) | 0.52 | 0.067 | 1 | 0.15 | ||||
| rs11536865 | ||||||||
| G An | 170 (96) | 198 (99) | 129 (99) | 327 (99) | 1.3 (0.12−14.53) | 4.65 (0.975−22.25) | 6.07 (0.75–49.1) | 5.12 (1.34–19.5) |
| C Mi | 8 (4) | 2 (1) | 1 (1) | 3 (1) | ||||
| p-value | 0.83 | 0.034 | 0.055 | 0.0081 | ||||
| G/C | 8 (9) | 2 (2) | 1 (2) | 3 (2) | 0.77 (0.07−8.62) | 0.21 (0.04−1.0) | 0.16 (0.02−1.30) | 0.19 (0.05−0.73) |
| G/G | 81 (91) | 98 (98) | 64 (98) | 162 (98) | 1.0 | 1.0 | 1.0 | 1.0 |
| p-value | 0.83 | 0.028 | 0.035 | 0.0092 | ||||
| HWE (p-value) | 1 | 1 | 1 | 1 |
| Model | Genotype | GP | DF | OR (95% IC) | p-Value | AIC |
|---|---|---|---|---|---|---|
| Codominant | A/A | 33 (37.1%) | 42 (42%) | 1.0 | 0.035 | 260.6 |
| A/G | 40 (44.9%) | 52 (52%) | 1.02 (0.55–1.89) | |||
| G/G | 16 (18%) | 6 (6%) | 0.29 (0.10–0.84) | |||
| Recessive | A/A-A/G | 73 (82%) | 94 (94%) | 1.00 | 0.0095 | 258.6 |
| G/G | 16 (18%) | 6 (6%) | 0.29 (0.11–0.78) |
| Model | Genotype | GP | DEN | OR (95% IC) | p-Value | AIC |
|---|---|---|---|---|---|---|
| Codominant | A/A | 33 (37.1%) | 73 (44.2%) | 1.0 | 0.038 | 328.5 |
| A/G | 40 (44.9%) | 80 (48.5%) | 0.9 (0.52–1.58) | |||
| G/G | 16 (18%) | 12 (7.3%) | 0.34 (0.14–0.8) | |||
| Recessive | A/A-A/G | 73 (82%) | 153 (92.7%) | 1.00 | 0.011 | 326.6 |
| G/G | 16 (18%) | 12 (7.3%) | 0.36 (0.16–0.8) |
| rs2737190 | rs4986791 | rs11536865 | |
|---|---|---|---|
| rs10759932 | 0.0642 | −0.0013 | 0.0041 |
| 0.999 | 0.842 | 0.212 | |
| 0.4533 | −0.0352 | 0.0953 | |
| 104.38 | 0.629 | 4.61 | |
| <2 × 10−16 | 0.4278 | 0.0318 | |
| 254 | 254 | 254 | |
| rs2737190 | 0.0102 | 0.006 | |
| 0.992 | 0.426 | ||
| 0.1724 | 0.0870 | ||
| 15.097 | 3.843 | ||
| 0.0001 | 0.0499 | ||
| 254 | 254 | ||
| rs4986791 | D | −0.0002 | |
| D’ | 0.768 | ||
| r | −0.0145 | ||
| χ2 | 0.106 | ||
| p-Value | 0.7446 | ||
| N | 254 |
| rs10759932 | rs2737190 | rs4986791 | rs11536865 | GP (n = 89) | DF (n = 100) | DHF (n = 65) | DEN (n = 165) | |
|---|---|---|---|---|---|---|---|---|
| 1 | T | A | C | G | 0.56 | 0.68 | 0.6939 | 0.6848 |
| 2 | T | G | C | G | 0.2999 | 0.19 | 0.2 | 0.1939 |
| 3 | C | G | C | G | 0.084 | 0.1 | 0.0846 | 0.0939 |
| 4 | T | G | T | G | 0.0056 | 0.02 | 0.154 | 0.0182 |
| 5 | T | A | C | C | 0.0355 | NA | NA | NA |
| 6 | T | G | C | C | 0.0091 | NA | NA | NA |
| 7 | C | G | C | C | 3e-04 | 0.01 | 0.0077 | 0.0091 |
| 8 | C | G | T | G | 0.0056 | NA | NA | NA |
| Groups | rs10759932 | rs2737190 | rs4986791 | rs11536865 | OR (95% CI), p-Value | Global Haplotype Association p-Value |
|---|---|---|---|---|---|---|
| GP vs. DF | T | A | C | G | 1.0 | |
| T | G | C | G | 0.55 (0.34–0.91), p-Value 0.021 | 0.0095 | |
| GP vs. DHF | T | A | C | G | 1.0 | |
| T | G | C | G | 0.59 (0.34–1.0), p-Value 0.05 | 0.036 | |
| GP vs. DEN | T | A | C | G | 1.0 | |
| T | G | C | G | 0.55 (0.35–0.86), p-Value 0.0084 | 0.0019 | |
| DF vs. DHF | T | A | C | G | 1.0 | |
| T | G | C | G | 1.02 (0.57–1.82), p-Value 0.95 | 0.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Posadas-Mondragón, A.; Aguilar-Faisal, J.L.; Zuñiga, G.; Magaña, J.J.; Santiago-Cruz, J.A.; Guillén-Salomón, E.; Alcántara-Farfán, V.; Arellano-Flores, M.L.; Salas-Benito, J.S.; Neri-Bazán, R.M.; et al. Association of Genetic Polymorphisms in TLR3, TLR4, TLR7, and TLR8 with the Clinical Forms of Dengue in Patients from Veracruz, Mexico. Viruses 2020, 12, 1230. https://doi.org/10.3390/v12111230
Posadas-Mondragón A, Aguilar-Faisal JL, Zuñiga G, Magaña JJ, Santiago-Cruz JA, Guillén-Salomón E, Alcántara-Farfán V, Arellano-Flores ML, Salas-Benito JS, Neri-Bazán RM, et al. Association of Genetic Polymorphisms in TLR3, TLR4, TLR7, and TLR8 with the Clinical Forms of Dengue in Patients from Veracruz, Mexico. Viruses. 2020; 12(11):1230. https://doi.org/10.3390/v12111230
Chicago/Turabian StylePosadas-Mondragón, Araceli, José Leopoldo Aguilar-Faisal, Gerardo Zuñiga, Jonathan Javier Magaña, José Angel Santiago-Cruz, Edith Guillén-Salomón, Verónica Alcántara-Farfán, María Luisa Arellano-Flores, Juan Santiago Salas-Benito, Rocío M. Neri-Bazán, and et al. 2020. "Association of Genetic Polymorphisms in TLR3, TLR4, TLR7, and TLR8 with the Clinical Forms of Dengue in Patients from Veracruz, Mexico" Viruses 12, no. 11: 1230. https://doi.org/10.3390/v12111230
APA StylePosadas-Mondragón, A., Aguilar-Faisal, J. L., Zuñiga, G., Magaña, J. J., Santiago-Cruz, J. A., Guillén-Salomón, E., Alcántara-Farfán, V., Arellano-Flores, M. L., Salas-Benito, J. S., Neri-Bazán, R. M., Luna-Rojas, L., Avila-Trejo, A. M., & Chávez-Negrete, A. (2020). Association of Genetic Polymorphisms in TLR3, TLR4, TLR7, and TLR8 with the Clinical Forms of Dengue in Patients from Veracruz, Mexico. Viruses, 12(11), 1230. https://doi.org/10.3390/v12111230