Lettuce Chlorosis Virus Disease: A New Threat to Cannabis Production

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Symptomatic Cannabis Plants
2.2. Small RNA Extraction, Next-Generation Sequencing (NGS)-Illumina MiSeq and Bioinformatics Analysis
2.3. Total RNA Extraction, NGS-Illumina HiSeq and Bioinformatics Analysis
2.4. Genome Assembly of the Cannabis Lettuce Chlorosis Virus (LCV) Isolate
2.5. Phylogenetic Tree Analysis
2.6. Whitefly Transmission Experiments of the Cannabis LCV Isolate
2.7. Partial Host Range of Cannabis LCV Isolate
2.8. LCV Transmission via Cannabis Shoots
2.9. Study of Seed Transmission of the Cannabis LCV
3. Results
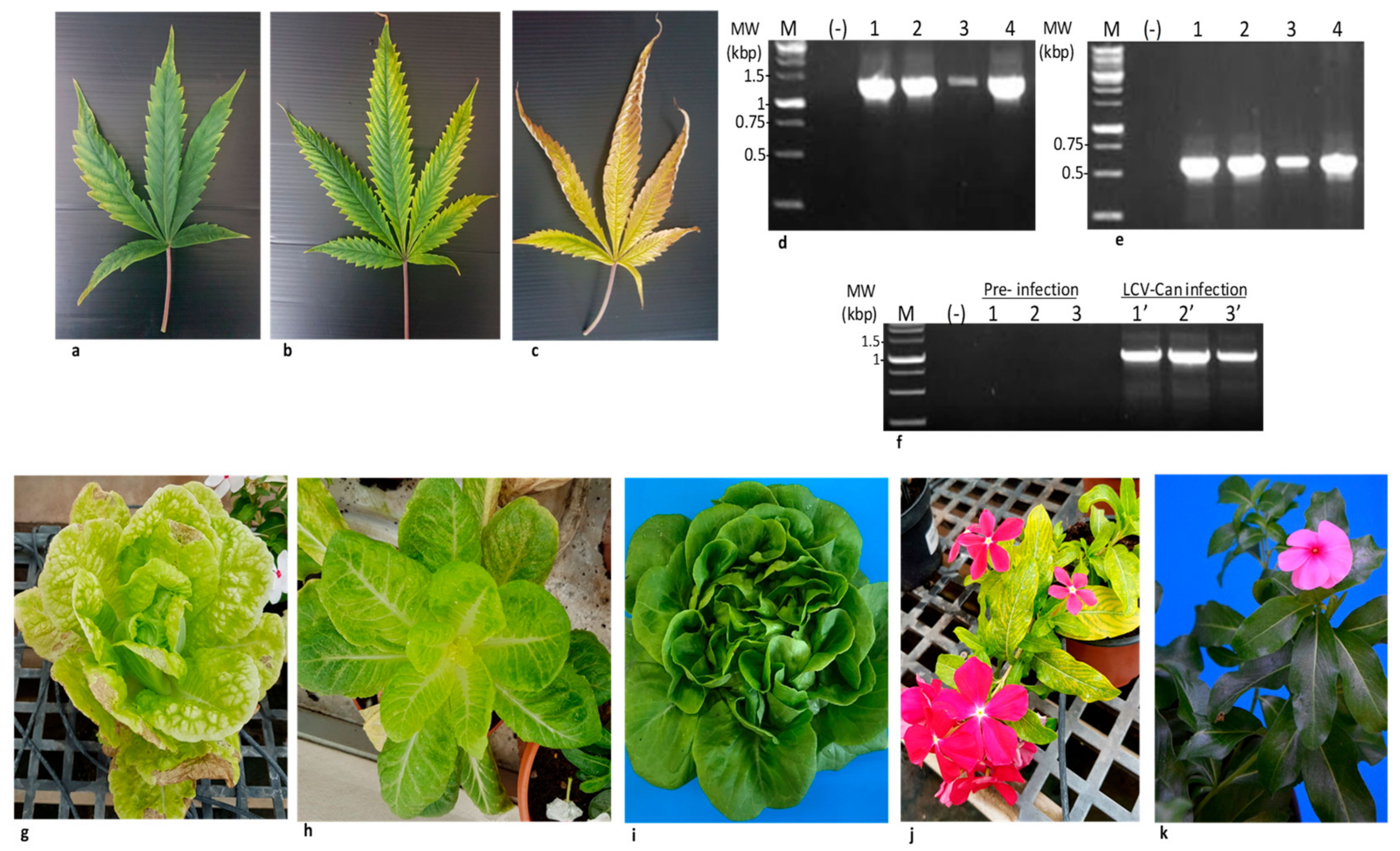
3.1. A Survey
3.2. Next Generation Sequencing Analysis of Small RNA Preparation
3.3. Next Generation Sequencing Analysis of Total RNA Preparations
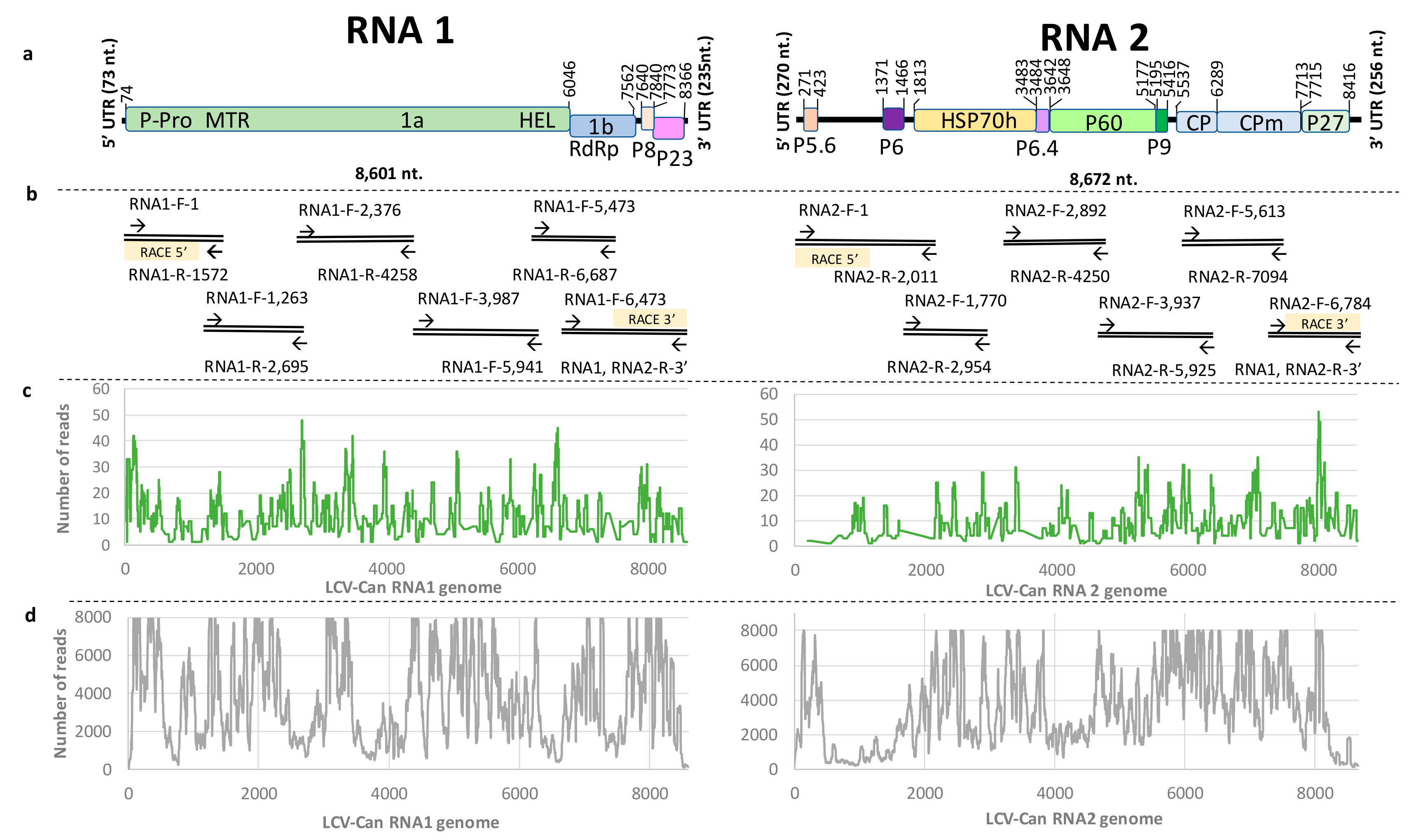
3.4. Revealing the Complete Genome Sequence of Cannabis LCV Isolate by RT-PCR Amplifications and Sanger Sequencing
3.5. Genome Organization of LCV-Can
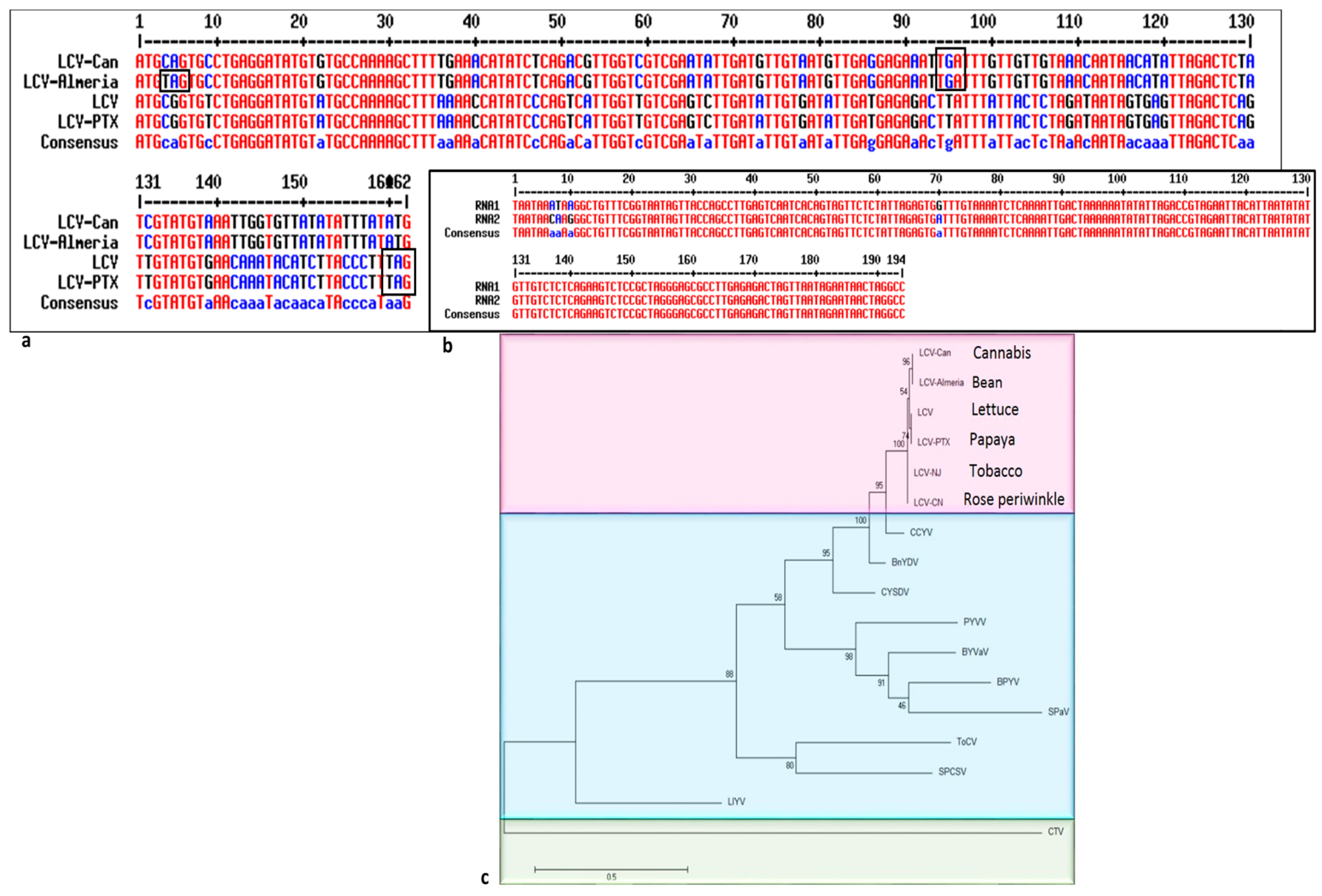
3.6. Consensus Motifs in LCV-Can
3.7. Phylogenetic Tree Analysis
3.8. Whitefly Transmission of LCV-Can in Cannabis Plants and Partial Host Range
3.9. LCV-Can Disease Spread via Shoots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rubin, V. The ”Ganja Vision” in Jamaica. In Cannabis and Culture; De Gruyter: Berlin, Germany, 1975; pp. 257–266. [Google Scholar]
- Berman, P.; Futoran, K.; Lewitus, G.M.; Mukha, D.; Benami, M.; Shlomi, T.; Meiri, D. A new ESI-LC/MS approach for comprehensive metabolic profiling of phytocannabinoids in Cannabis. Sci. Rep. 2018, 8, 14280. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.B.; Hohmann, A.G. Role of cannabinoids in pain management. In Comprehensive Treatment of Chronic Pain by Medical, Interventional, and Integrative Approaches; Springer: Brelin, Germany, 2013; pp. 181–197. [Google Scholar]
- Grund, J.-P.C.; Breeksema, J.J. Drug policy in the Netherlands. In European Drug Policies: The Ways of Reform; Routledge: Abingdon on Thames, UK, 2017; pp. 128–148. [Google Scholar]
- Zaami, S.; Di Luca, A.; Di Luca, N.; Vergallo, G.M. Medical use of cannabis: Italian and European legislation. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1161–1167. [Google Scholar] [PubMed]
- Room, R. Legalizing a market for cannabis for pleasure: Colorado, Washington, Uruguay and beyond. Addiction 2014, 109, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Hughes, B.; Wiessing, L.; Jarlais, D.D.; Griffiths, P. Could cannabis liberalisation lead to wider changes in drug policies and outcomes? Int. J. Drug Policy 2018, 51, 156–159. [Google Scholar] [CrossRef] [PubMed]
- King, K.C.; Lively, C.M. Does genetic diversity limit disease spread in natural host populations? Heredity 2012, 109, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.; Borja, M. High incidence of Pelargonium line pattern virus infecting asymptomatic Pelargonium spp. in Spain. Eur. J. Plant Pathol. 2005, 112, 95–100. [Google Scholar] [CrossRef]
- Röder, K. Einige Untersuchungen über ein an Hanf (Cannabis sativa L.) auftretendes Virus. Faserforschung 1941, 15, 77. [Google Scholar]
- Ceapoiu, N. Cinepa: Estudiu Monografic. In Institutul de Cercetari Agronomice; CABI: Bucarest, Romania, 1958. [Google Scholar]
- McPartland, J.M.; Clarke, R.C.; Watson, D.P. Hemp Diseases and Pests: Management and Biological Control—An Advanced Treatise; CABI: Wallingford, UK, 2000. [Google Scholar]
- Bektas, A.; Hardwick, K.M.; Waterman, K.; Kristof, J. The Occurrence of Hop Latent Viroid in Cannabis sativa with symptoms of Cannabis Stunting Disease in California. Plant Dis. 2019. [Google Scholar] [CrossRef]
- Warren, J.; Mercado, J.; Grace, D. The occurrence of Hop latent viroid causing disease in Cannabis sativa in California. Plant Dis. 2019. [Google Scholar] [CrossRef]
- Ziegler, A.; Matoušek, J.; Steger, G.; Schubert, J. Complete sequence of a cryptic virus from hemp (Cannabis sativa). Arch. Virol. 2012, 157, 383–385. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging Virus Diseases Transmitted by Whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Tzanetakis, I.E.; Martin, R.R.; Wintermantel, W.M. Epidemiology of criniviruses: An emerging problem in world agriculture. Front. Microbiol. 2013, 4, 119. [Google Scholar] [CrossRef] [PubMed]
- Luria, N.; Smith, E.; Sela, N.; Koren, A.; Lachman, O.; Dombrovsky, A. Insights Into a Watermelon Virome Contribute to Monitoring Distribution of Whitefly-Borne Viruses. Phytobiomes J. 2019, 3, 61–70. [Google Scholar] [CrossRef]
- Sela, N.; Luria, N.; Dombrovsky, A. Genome Assembly of Bell Pepper Endornavirus from Small RNA. J. Virol. 2012, 86, 7721. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Stocks, M.B.; Moxon, S.; Mapleson, D.; Woolfenden, H.C.; Mohorianu, I.; Folkes, L.; Schwach, F.; Dalmay, T.; Moulton, V. The UEA sRNA workbench: A suite of tools for analysing and visualizing next generation sequencing microRNA and small RNA datasets. Bioinformatics 2012, 28, 2059–2061. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Zheng, Y.; Gao, S.; Padmanabhan, C.; Li, R.; Galvez, M.; Gutierrez, D.; Fuentes, S.; Ling, K.-S.; Kreuze, J.; Fei, Z. VirusDetect: An automated pipeline for efficient virus discovery using deep sequencing of small RNAs. Virology 2017, 500, 130–138. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Frohman, M.A. RACE: Rapid amplification of cDNA ends. In PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; pp. 28–38. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Boil. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.M.; Chen, A.Y.; Tzanetakis, I.E.; Mongkolsiriwattana, C.; Ng, J.C. Further complexity of the genus Crinivirus revealed by the complete genome sequence of Lettuce chlorosis virus (LCV) and the similar temporal accumulation of LCV genomic RNAs 1 and 2. Virology 2009, 390, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Simón, A.; García, C.; Velasco, L.; Janssen, D. First natural crossover recombination between two distinct species of the family Closteroviridae leads to the emergence of a new disease. PLoS ONE 2018, 13, e0198228. [Google Scholar] [CrossRef]
- Rozanov, M.N.; Koonin, E.V.; Gorbalenya, A.E. Conservation of the putative methyltransferase domain: A hallmark of the ‘Sindbis-like’ supergroup of positive-strand RNA viruses. J. Gen. Virol. 1992, 73, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Tzanetakis, I.; Reed, J.; Martin, R. Nucleotide sequence, genome organization and phylogenetic analysis of Strawberry pallidosis associated virus, a new member of the genus Crinivirus. Arch. Virol. 2005, 150, 273–286. [Google Scholar] [CrossRef]
- Koonin, E.V. The phylogeny of RNA-dependent RNA polymerases of positive-strand RNA viruses. J. Gen. Virol. 1991, 72, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- Bork, P.; Sander, C.; Valencia, A. An ATPase domain common to prokaryotic cell cycle proteins, sugar kinases, actin, and hsp70 heat shock proteins. Proc. Natl. Acad. Sci. USA 1992, 89, 7290–7294. [Google Scholar] [CrossRef]
- Alzhanova, D.V.; Napuli, A.J.; Creamer, R.; Dolja, V.V. Cell-to-cell movement and assembly of a plant closterovirus: Roles for the capsid proteins and Hsp70 homolog. EMBO J. 2001, 20, 6997–7007. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: San Diego, CA, USA, 2012. [Google Scholar]
- Klaassen, V.A.; Boeshore, M.L.; Koonin, E.V.; Tian, T.; Falk, B.W. Genome Structure and Phylogenetic Analysis of Lettuce Infectious Yellows Virus, a Whitefly-Transmitted, Bipartite Closterovirus. Virology 1995, 208, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Duffus, J.E.; Liu, H.-Y.; Wisler, G.C.; Li, R. Lettuce chlorosis virus—A new whitefly-transmitted closterovirus. Eur. J. Plant Pathol. 1996, 102, 591–596. [Google Scholar] [CrossRef]
- Hillig, K.W.; Mahlberg, P.G. A chemotaxonomic analysis of cannabinoid variation in Cannabis (Cannabaceae). Am. J. Bot. 2004, 91, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the Insect Supervectors Bemisia tabaci and Frankliniella occidentalis in the Emergence and Global Spread of Plant Viruses. Annu. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.L.; Simon, A.; Garcia, M.C.; Janssen, D. First Report of Lettuce chlorosis virus Infecting Bean in Spain. Plant Dis. 2014, 98, 857. [Google Scholar] [CrossRef]
- Alabi, O.J.; Al Rwahnih, M.; Jifon, J.L.; Sétamou, M.; Brown, J.K.; Gregg, L.; Park, J.-W. A mixed infection of Lettuce chlorosis virus, Papaya ringspot virus, and Tomato yellow leaf curl virus-IL detected in a Texas papaya orchard affected by a virus-like disease outbreak. Plant Dis. 2017, 101, 1094–1102. [Google Scholar] [CrossRef] [PubMed][Green Version]
- De Barro, P.J.; Liu, S.-S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A Statement of Species Status. Annu. Rev. Èntomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Hadjistylli, M.; Roderick, G.K.; Brown, J.K. Global Population Structure of a Worldwide Pest and Virus Vector: Genetic Diversity and Population History of the Bemisia tabaci Sibling Species Group. PLoS ONE 2016, 11, 0165105. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, M. A review of the mechanisms and components that determine the transmission efficiency of Tomato yellow leaf curl virus (Geminiviridae; Begomovirus) by its whitefly vector. Virus Res. 2014, 186, 47–54. [Google Scholar] [CrossRef]
- Loebenstein, G.; Lecoq, H. Viruses and Virus Diseases of Vegetables in the Mediterranean Basin; Elsevier: San Diego, CA, USA; Academic Press: San Diego, CA, USA, 2012; Volume 84. [Google Scholar]
- Kubota, K.; Ng, J. Lettuce chlorosis virus P23 Suppresses RNA Silencing and Induces Local Necrosis with Increased Severity at Raised Temperatures. Phytopathology 2016, 106, 653–662. [Google Scholar] [CrossRef]
- Cantino, P.D.; Doyle, J.A.; Graham, S.W.; Judd, W.S.; Olmstead, R.G.; Soltis, D.E.; Soltis, P.S.; Donoghue, M.J.; Wang, W.; Chen, Z.-D. Towards a phylogenetic nomenclature of Tracheophyta. TAXON 2007, 56, 822–846. [Google Scholar] [CrossRef]
- Kiss, Z.A.; Medina, V.; Falk, B.W. Crinivirus replication and host interactions. Front. Microbiol. 2013, 4, 99. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Pendon, J.A.; Ding, S.-W. Direct and Indirect Roles of Viral Suppressors of RNA Silencing in Pathogenesis. Annu. Rev. Phytopathol. 2008, 46, 303–326. [Google Scholar] [CrossRef] [PubMed]
- Alzhanova, D.V.; Hagiwara, Y.; Peremyslov, V.V.; Dolja, V.V. Genetic Analysis of the Cell-to-Cell Movement of Beet Yellows Closterovirus. Virology 2000, 268, 192–200. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhao, X.; Zhu, M.; Wu, Q.; Zhang, J.; Xu, Y.; Tao, X. Complete genome sequence of a lettuce chlorosis virus isolate from China and genome recombination/rearrangement analysis. Arch. Virol. 2018, 163, 751–754. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Small RNA | Total RNA | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Reference Genome a | Complete Genome (nt) | No. Contigs b | Coverage c (%) | Depth d | Identity e | No. Contigs | Coverage (%) | Depth | Identity |
| Lettuce chlorosis virus (LCV) RNA1 | 8591 | 22 | 1054 (12.3%) | 5.7 | 97.25 | 39 | 7103 (82.7%) | 2671.7 | 89.78 |
| Lettuce chlorosis virus (LCV) RNA2 | 8556 | 38 | 2091 (24.4%) | 5.3 | 97.32 | 52 | 6722 (78.6%) | 2499.3 | 92.61 |
| Viruses a | LCV-Can | LCV-Almeria | LCV (California) | LCV-PTX | LCV-CN | LCV-NJ |
|---|---|---|---|---|---|---|
| RNA1 Putative proteins (aa.) | Whole genome (nt.) | 99 | 89 | 89 | 85 | 85 |
| 1a | 99 | 94 | 94 | 91 | 91 | |
| RdRp | 100 | 99 | 99 | 99 | 99 | |
| P8 | - | 74 | 74 | 55 | 57 | |
| P23 | - | 75 | 75 | 77 | 77 | |
| RNA2 Putative proteins (aa.) | Whole genome (nt.) | 99 | 90 | 90 | 89 | 89 |
| P5.6 | 100 | 92 | 92 | 91 | 91 | |
| P6-like | - | 52 | 52 | - | - | |
| HSP70 | 100 | 98 | 98 | 99 | 98 | |
| P6.4 | 100 | 98 | 98 | 98 | 98 | |
| P60 | 99 | 97 | 97 | 95 | 95 | |
| P9 | 100 | 96 | 96 | 96 | 96 | |
| CP | 100 | 96 | 95 | 98 | 98 | |
| CPm | 99 | 96 | 96 | 92 | 92 | |
| P27 | 99 | 95 | 97 | 97 | 97 |
| * Set No. | ** Orientation | Name-Position | Sequence (5′-3′) |
|---|---|---|---|
| 8 | F | RNA1-F-7170 | TCACAGCCGAGATCAACAGA |
| 8 | C | RNA1-R-8433 | GTTACCAGCCTTGAGTCAATCA |
| 12 | F | RNA2-F-6090 | TCATCTTCAGGCCAAACACGG |
| 12 | C | RNA2-R-7094 | TCCACCTAATCCGATTCCAC |
| 13 | F | RNA2-F-7628 | GCAGGTCATGACGTCAGATTT |
| 13 | C | RNA2-R-8189 | TGAACAATCACTACAGGTTTGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hadad, L.; Luria, N.; Smith, E.; Sela, N.; Lachman, O.; Dombrovsky, A. Lettuce Chlorosis Virus Disease: A New Threat to Cannabis Production. Viruses 2019, 11, 802. https://doi.org/10.3390/v11090802
Hadad L, Luria N, Smith E, Sela N, Lachman O, Dombrovsky A. Lettuce Chlorosis Virus Disease: A New Threat to Cannabis Production. Viruses. 2019; 11(9):802. https://doi.org/10.3390/v11090802
Chicago/Turabian StyleHadad, Lior, Neta Luria, Elisheva Smith, Noa Sela, Oded Lachman, and Aviv Dombrovsky. 2019. "Lettuce Chlorosis Virus Disease: A New Threat to Cannabis Production" Viruses 11, no. 9: 802. https://doi.org/10.3390/v11090802
APA StyleHadad, L., Luria, N., Smith, E., Sela, N., Lachman, O., & Dombrovsky, A. (2019). Lettuce Chlorosis Virus Disease: A New Threat to Cannabis Production. Viruses, 11(9), 802. https://doi.org/10.3390/v11090802