Bio-Insecticidal Potential of Nucleopolyhedrovirus and Granulovirus Mixtures to Control the Fall Armyworm Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Virus Production, Purification and Quantification
2.3. Evaluation of Mixtures between Granulovirus and Nucleopolyhedrovirus
2.4. Assessment of Enhancer Activity of Proteins Derived from VG008
2.5. Characterization of Selected Mixture Formulated as Wettable Powder
2.5.1. Moisture Content
2.5.2. pH
2.5.3. Contaminants Content
2.5.4. Insecticidal Activity in Laboratory
2.5.5. Photostability
2.6. Greenhouse Trial to Select Application Dose
3. Results
3.1. Mixtures between Granulovirus and Nucleopolyhedrovirus
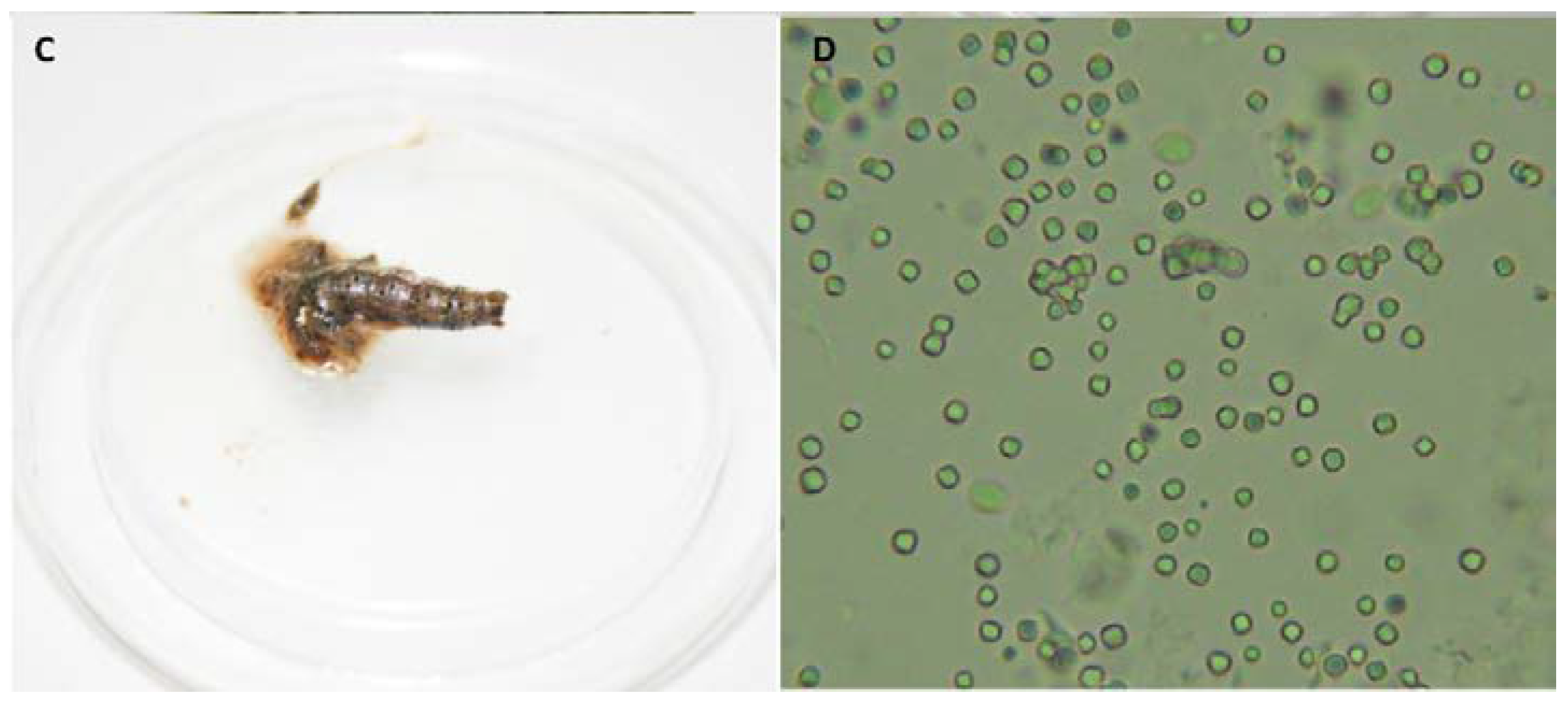
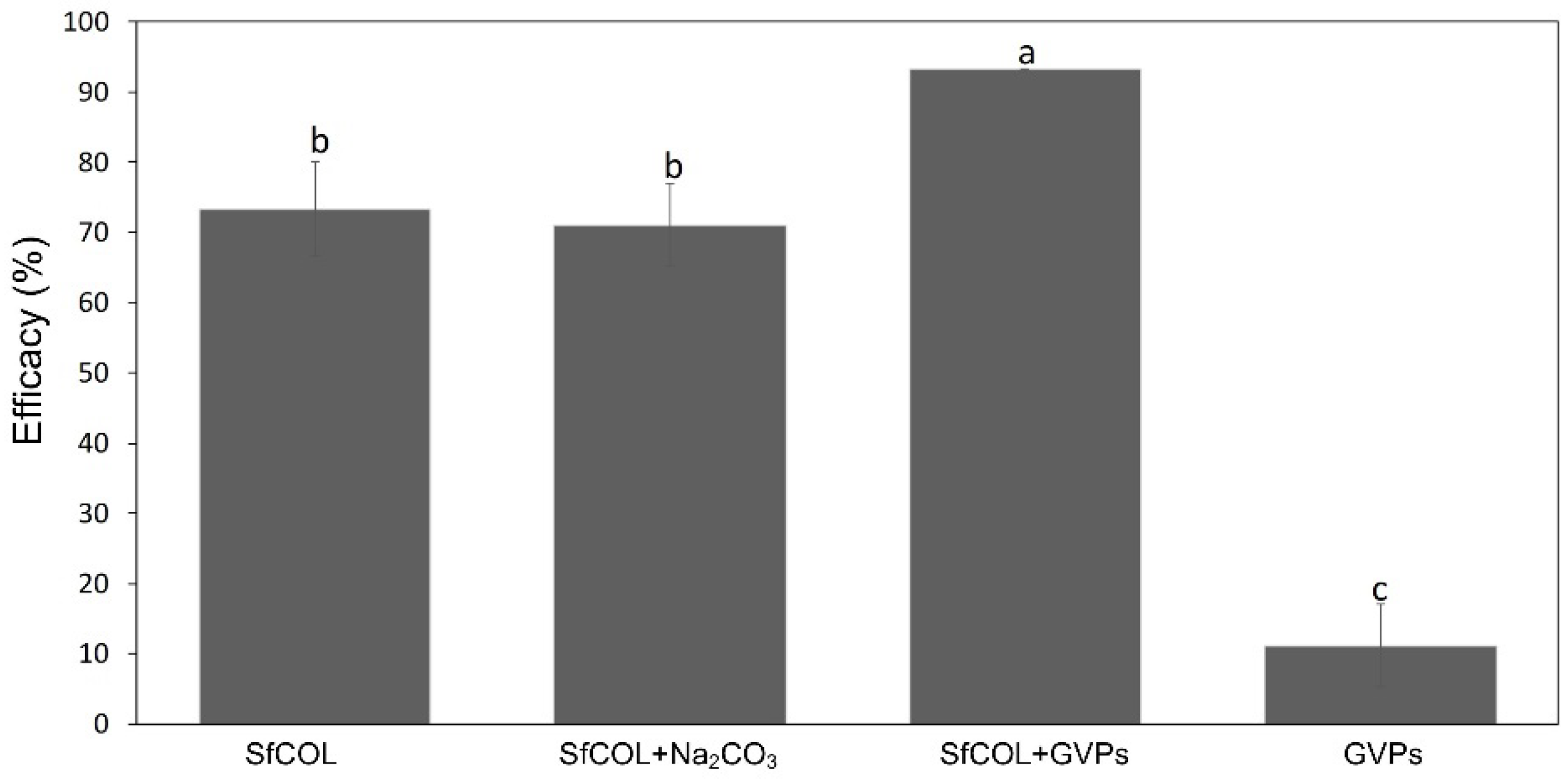
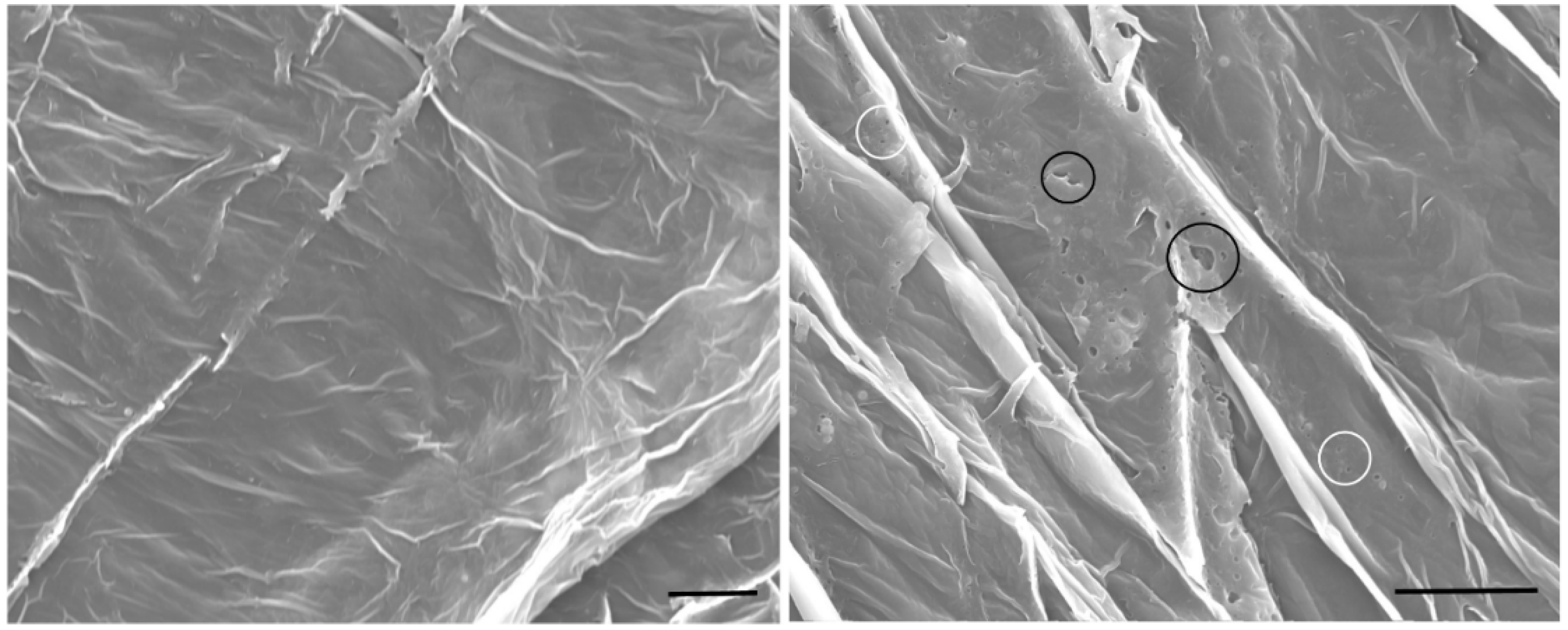
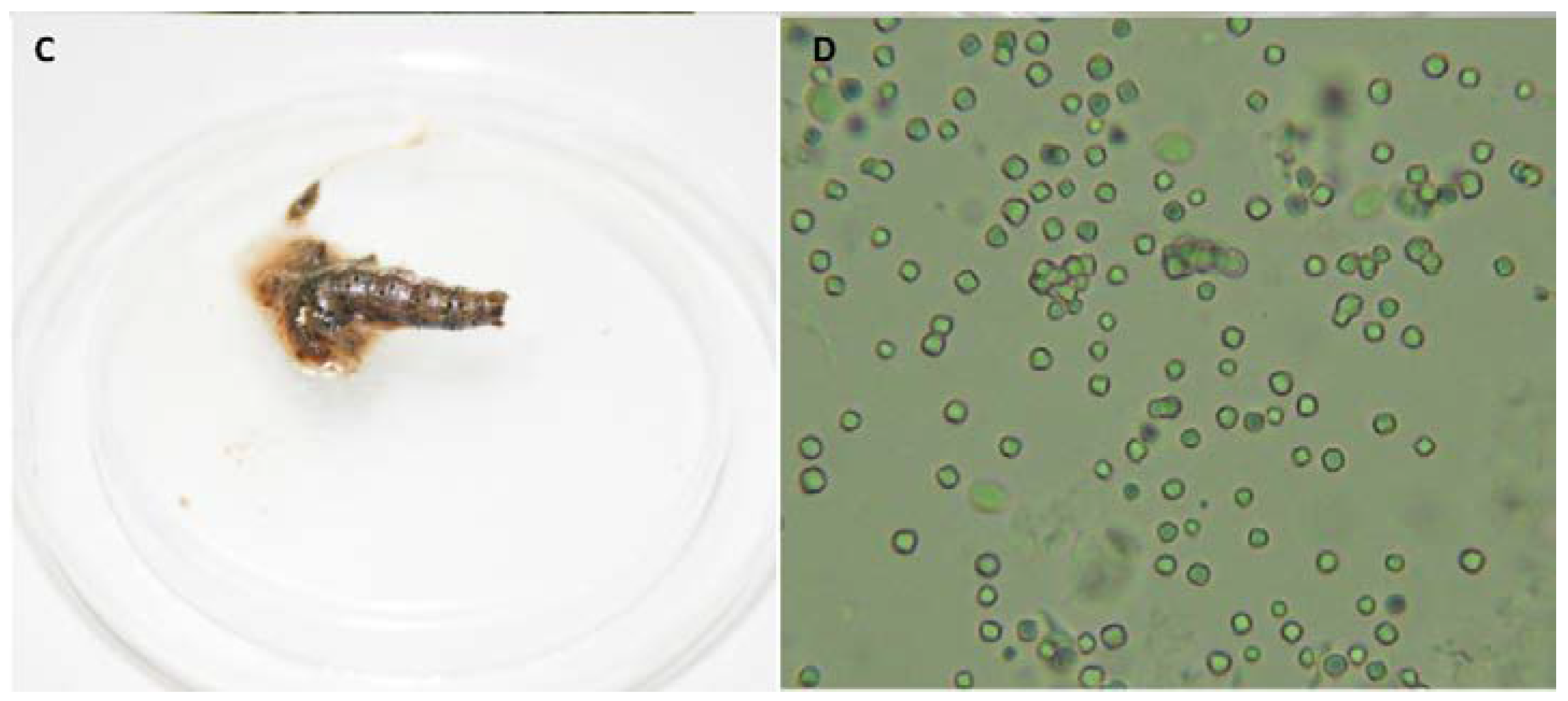
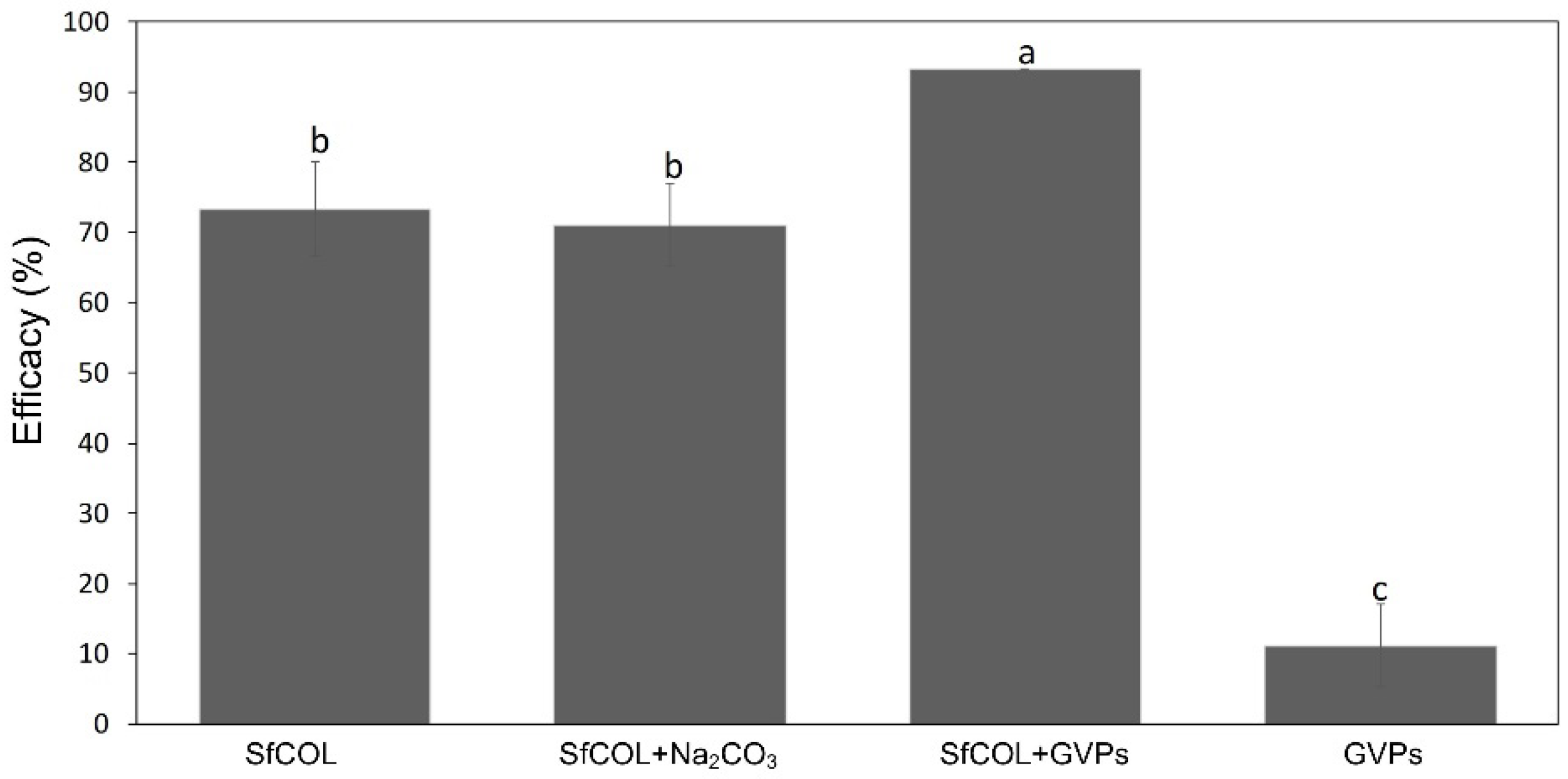
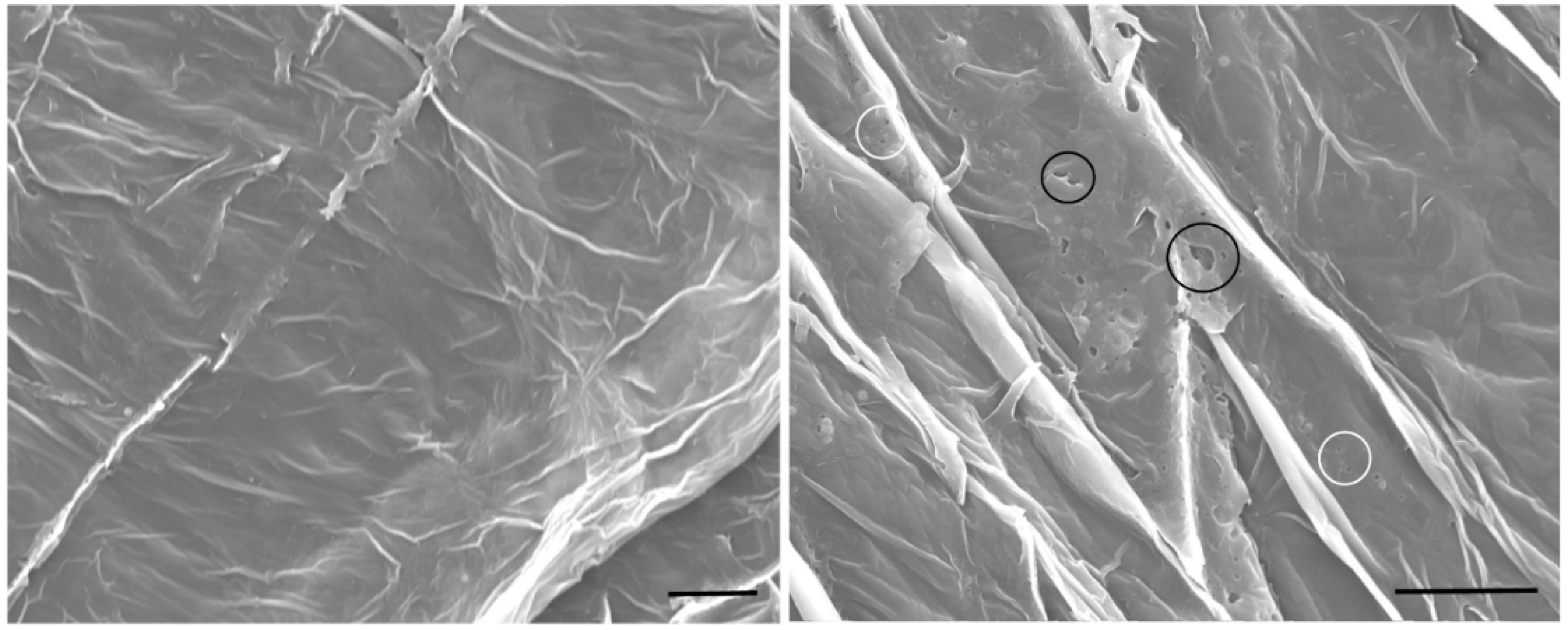
3.2. Enhancer Activity of Proteins Derived from VG008
3.3. Characterization of Selected Mixture Formulated as Wettable Powder
3.4. Greenhouse Trial to Select Application Dose
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- ICA (Instituto Colombiano Agropecuario). Resolución No. 000879 (25 Marzo 2008). Available online: https://www.ica.gov.co/getattachment/29ca37cc-b101-438e-8251-4803a18203cc/2008R0879.aspx (accessed on 28 May 2019).
- Lapied, B.; Pennetier, C.; Apaire-Marchais, V.; Licznar, P.; Corbel, V. Innovative applications for insect viruses: Towards insecticide sensitization. Trends Biotechnol. 2009, 27, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.L.; Herniou, E.A.; Jehle, J.A.; Theilmann, D.A.; Burand, J.P.; Becnel, J.J.; Krell, P.J.; van Oers, M.M.; Mowery, J.D.; Bauchan, G.R.; et al. ICTV Virus Taxonomy Profile: Baculoviridae. J. Gen. Virol. 2018, 99, 1185–1186. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.-M.; Pineda, S.; Figueroa, J.I.; Chavarrieta, J.M.; Williams, T. Los baculovirus como bioinsecticidas: Evaluación de un nucleopoliedrovirus para el combate de Spodoptera frugiperda (Lepidoptera: Noctuidae) en México y Honduras. Cienc. Nicolaita 2012, 56, 35–47. [Google Scholar]
- Behle, R.W.; Popham, H.J. Laboratory and field evaluations of the efficacy of a fast-killing baculovirus isolate from Spodoptera frugiperda. J. Invertebr. Pathol. 2012, 109, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.; Guevara, J.; Cuartas, P.; Espinel, C.; Villamizar, L. Microencapsulated Spodoptera frugiperda nucleopolyhedrovirus: Insecticidal activity and effect on arthropod populations in maize. Biocontrol Sci. Technol. 2013, 23, 829–846. [Google Scholar] [CrossRef]
- Slavicek, J. Baculovirus enhancins and their role in viral pathogenicity. In Molecular Biology; Adoga, M.P., Ed.; IntechOpen: Rijeka, Croatia, 2012; pp. 147–168. [Google Scholar]
- Gómez Valderrama, J.A.; Guevara Agudelo, E.J.; Barrera Cubillos, G.P.; Cotes Prado, A.M.; Villamizar Rivero, L.F. Aislamiento, identificación y caracterización de nucleopoliedrovirus nativos de Spodoptera frugiperda en Colombia. Rev. Fac. Nac. Agron. Medellín 2010, 63, 5511–5520. [Google Scholar]
- Cuartas, P.; Barrera, G.; Barreto, E.; Villamizar, L. Characterisation of a Colombian granulovirus (Baculoviridae: Betabaculovirus) isolated from Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae. Biocontrol Sci. Technol. 2014, 24, 1265–1285. [Google Scholar] [CrossRef]
- Barrera, G.P.; Belaich, M.N.; Patarroyo, M.A.; Villamizar, L.F.; Ghiringhelli, P.D. Evidence of recent interspecies horizontal gene transfer regarding nucleopolyhedrovirus infection of Spodoptera frugiperda. BMC genomics 2015, 16, 1008. [Google Scholar] [CrossRef]
- Cuartas, P.E.; Barrera, G.P.; Belaich, M.N.; Barreto, E.; Ghiringhelli, P.D.; Villamizar, L.F. The complete sequence of the first Spodoptera frugiperda Betabaculovirus genome: A natural multiple recombinant virus. Viruses 2015, 7, 394–421. [Google Scholar] [CrossRef]
- Toprak, U.; Harris, S.; Baldwin, D.; Theilmann, D.; Gillott, C.; Hegedus, D.D.; Erlandson, M.A. Role of enhancin in Mamestra configurata nucleopolyhedrovirus virulence: Selective degradation of host peritrophic matrix proteins. J. Gen. Virol. 2012, 93, 744–753. [Google Scholar] [CrossRef]
- Salvador, R.; Ferrelli, M.L.; Sciocco-Cap, A.; Romanowski, V. Analysis of a chitinase from EpapGV, a fast killing betabaculovirus. Virus Genes 2014, 48, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Greene, G.L.; Leppla, N.; Dickerson, W.A. Velvetbean Caterpillar: A Rearing Procedure and Artificial Medium. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar] [CrossRef]
- Hughes, P.R.; Wood, H.A. A synchronous peroral technique for the bioassay of insect viruses. J. Invertebr. Pathol. 1981, 37, 154–159. [Google Scholar] [CrossRef]
- Bullock, J.; Crawley, M.J. GLIM for Ecologists; Wiley-Blackwell: Hoboken, NJ, USA, 1994; Volume 31, p. 402. [Google Scholar]
- Zar, J. Biostatistical Analysis, 5th ed.; Prentice-Hall, Inc.: Upper Saddle River, NJ, USA, 2007. [Google Scholar]
- Cuartas, P.E.; Villamizar, L.F.; Barrera, G.P.; Ruiz, J.C.; Campos, J.C.; Leon-Martinez, G.; Gomez-Valderrama, J. Novel biopesticide based on Erinnyis ello betabaculovirus: Characterization and preliminary field evaluation to control Erinnyis ello in rubber plantations. Pest Manag. Sci. 2019, 75, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, I.; Gómez, M.; Villamizar, L. Estabilidad de formulaciones a base de granulovirus para controlar Teci solanivora (Lepidotera: Gelechiidae) en campo. Rev. Colomb. Entomol. 2011, 37, 27–35. [Google Scholar]
- Villamizar, L.; Barrera, G.; Cotes, A.M.; Martínez, F. Eudragit S100 microparticles containing Spodoptera frugiperda nucleopolyehedrovirus: Physicochemical characterization, photostability and in vitro virus release. J. Microencapsul. 2010, 27, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G. SDS-PAGE of GVPs from SpfrGV VG008. Corporación Colombiana de Investigación Agropecuaria-Agrosavia. 2019; (not intended for publication). [Google Scholar]
- Ravensberg, W. A Roadmap to the Successful Development and Commercialization of Microbial Pest Control Products for Control of Arthropods; Springer: Berlin, Germany, 2011. [Google Scholar]
- Guo, H.F.; Fang, J.C.; Wang, J.P.; Zhong, W.F.; Liu, B.S. Interaction of Xestia c-nigrum granulovirus with peritrophic matrix and Spodoptera litura nucleopolyhedrovirus in Spodoptera litura. J. Econ. Entomol. 2007, 100, 20–25. [Google Scholar] [CrossRef]
- Hayakawa, T.; Shimojo, E.-I.; Mori, M.; Kaido, M.; Furusawa, I.; Miyata, S.; Sano, Y.; Matsumoto, T.; Hashimoto, Y.; Granados, R.R. Enhancement of baculovirus infection in Spodoptera exigua (Lepidoptera: Noctuidae) larvae with Autographa californica nucleopolyhedrovirus or Nicotiana tabacum engineered with a granulovirus enhancin gene. J. Appl. Entomol. Z. 2000, 35, 163–170. [Google Scholar] [CrossRef]
- Mukawa, S.; Goto, C. Mamestra brassicae nucleopolyhedrovirus infection and enhancing effect of proteins derived from Xestia c-nigrum granulovirus in larvae of Mamestra brassicae and Helicoverpa armigera (Lepidoptera: Noctuidae) on cabbage. J. Econ. Entomol. 2010, 103, 257–264. [Google Scholar] [CrossRef]
- Mukawa, S.; Goto, C. Enhancement of nucleopolyhedrovirus infectivity against Mamestra brassicae (Lepidoptera: Noctuidae) by proteins derived from granulovirus and a fluorescent brightener. J. Econ. Entomol. 2007, 100, 1075–1083. [Google Scholar] [CrossRef]
- Shapiro, M. Effect of two granulosis viruses on the activity of the gypsy moth (Lepidoptera: Lymantriidae) nuclear polyhedrosis virus. J. Econ. Entomol. 2000, 93, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Corsaro, B.G.; Gijzen, M.; Wang, P.; Granados, R. Baculovirus enhancing proteins as determinants of viral pathogenesis. In Parasites and Pathogens of Insects; Academic Press: New York, NY, USA, 1993; pp. 127–145. [Google Scholar]
- Hoover, K.; Humphries, M.A.; Gendron, A.R.; Slavicek, J.M. Impact of viral enhancin genes on potency of Lymantria dispar multiple nucleopolyhedrovirus in L. dispar following disruption of the peritrophic matrix. J. Invertebr. Pathol. 2010, 104, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Goto, C.; Mukawa, S.; Mitsunaga, T. Two year field study to evaluate the efficacy of Mamestra brassicae nucleopolyhedrovirus combined with proteins derived from Xestia c-nigrum granulovirus. Viruses 2015, 7, 1062–1078. [Google Scholar] [CrossRef]
- Biedma, M.E.; Salvador, R.; Ferrelli, M.L.; Sciocco-Cap, A.; Romanowski, V. Effect of the interaction between Anticarsia gemmatalis multiple nucleopolyhedrovirus and Epinotia aporema granulovirus, on A. gemmatalis (Lepidoptera: Noctuidae) larvae. Biol. Control 2015, 91, 17–21. [Google Scholar] [CrossRef]
- Li, Z.; Li, C.; Yang, K.; Wang, L.; Yin, C.; Gong, Y.; Pang, Y. Characterization of a chitin-binding protein GP37 of Spodoptera litura multicapsid nucleopolyhedrovirus. Virus Res. 2003, 96, 113–122. [Google Scholar] [CrossRef]
- Martemyanov, V.V.; Kabilov, M.R.; Tupikin, A.E.; Baturina, O.A.; Belousova, I.A.; Podgwaite, J.D.; Ilynykh, A.V.; Vlassov, V.V. The enhancin gene: One of the genetic determinants of population variation in baculoviral virulence. Dokl. Biochem. Biophys. 2015, 465, 351–353. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, M.; Preisler, H.K.; Robertson, J.L. Enhancement of baculovirus activity on gypsy moth (Lepidoptera: Lymantriidae) by chitinase. J. Econ. Entomol. 1987, 80, 1113–1116. [Google Scholar] [CrossRef]
- Rao, R.; Fiandra, L.; Giordana, B.; de Eguileor, M.; Congiu, T.; Burlini, N.; Arciello, S.; Corrado, G.; Pennacchio, F. AcMNPV ChiA protein disrupts the peritrophic membrane and alters midgut physiology of Bombyx mori larvae. Insect Biochem. Mol. Biol. 2004, 34, 1205–1213. [Google Scholar] [CrossRef]
- Hackett, K.; Boore, A.; Deming, C.; Buckley, E.; Camp, M.; Shapiro, M. Helicoverpa armigera granulovirus interference with progression of H. zea nucleopolyhedrovirus disease in H. zea larvae. J. Invertebr. Pathol. 2000, 75, 99–106. [Google Scholar] [CrossRef]
- Ishimwe, E.; Hodgson, J.J.; Passarelli, A.L. Expression of the Cydia pomonella granulovirus matrix metalloprotease enhances Autographa californica multiple nucleopolyhedrovirus virulence and can partially substitute for viral cathepsin. Virology 2015, 481, 166–178. [Google Scholar] [CrossRef]
- Tellam, R.L.; Wijffels, G.; Willadsen, P. Peritrophic matrix proteins. Insect Biochem. Mol. Biol. 1999, 29, 87–101. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Martin, M.M. Peritrophic envelope permeability in herbivorous insects. J. Insect Physiol. 1995, 41, 303–311. [Google Scholar] [CrossRef]
- Jenkins, N.E.; Grzywacz, D. Quality control of fungal and viral biocontrol agents - assurance of product performance. Biocontrol Sci. Techn. 2000, 10, 753–777. [Google Scholar] [CrossRef]
- Slack, J.; Arif, B. The baculoviruses occlusion-derived virus: Virion structure and function. Adv. Virus Res. 2007, 69, 99–165. [Google Scholar] [PubMed]
- Lasa, R.; Williams, T.; Caballero, P. Insecticidal properties and microbial contaminants in a Spodoptera exigua multiple nucleopolyhedrovirus (Baculoviridae) formulation stored at different temperatures. J. Econ. Entomol. 2008, 101, 42–49. [Google Scholar] [CrossRef]
- Yang, M.M.; Li, M.L.; Zhang, Y.; Wang, Y.Z.; Qu, L.J.; Wang, Q.H.; Ding, J.Y. Baculoviruses and insect pests control in China. Afr. J. Microbiol. Res. 2012, 6, 214–218. [Google Scholar]
- Grzywacz, D.; Moore, S. Chapter 7—Production, formulation, and bioassay of baculoviruses for pest control. In Microbial Control of Insect and Mite Pests; Lacey, L.A., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 109–124. [Google Scholar]
- Martı́nez, A.M.; Goulson, D.; Chapman, J.W.; Caballero, P.; Cave, R.D.; Williams, T. Is it feasible to use optical brightener technology with a baculovirus bioinsecticide for resource-poor maize farmers in Mesoamerica? Biol. Control 2000, 17, 174–181. [Google Scholar] [CrossRef]
- Castillejos, V.; Trujillo, J.; Ortega, L.D.; Santizo, J.A.; Cisneros, J.; Penagos, D.I.; Valle Mora, J.; Williams, T. Granular phagostimulant nucleopolyhedrovirus formulations for control of Spodoptera frugiperda in maize. Biol. Control. 2002, 24, 300–310. [Google Scholar] [CrossRef]
- Virus based biopesticide: Invention patent requested for United States. Available online: https://patentimages.storage.googleapis.com/a9/9f/3f/d8411d6cce024b/US20170172154A1.pdf (accessed on 28 May 2019).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Total Dose OBs/ha | Viral Mixture Composition | |
|---|---|---|---|
| NPV OBs (97.5%) | GV OBs (2.5%) | ||
| M3 Dose 1 | 5.0 × 1010 | 4.9 × 1010 | 1.2 × 109 |
| M3 Dose 2 | 1.0 × 1011 | 9.8 × 1011 | 2.4 × 109 |
| M3 Dose 3 | 2.0 × 1011 | 2.0 × 1011 | 4.9 × 109 |
| M3 Dose 4 | 4.0 × 1011 | 3.9 × 1011 | 9.8 × 109 |
| M3 Dose 5 | 8.0 × 1011 | 7.8 × 1011 | 2.0 × 1010 |
| SfCOL-F | 1.5 × 1012 | 1.5 × 1012 | - |
| Treatment | LC50 (OBs/mL) | Fiducial Limits (95%) | RP | P Value | MTD (Days) | Fiducial Limits (95%) | ||
|---|---|---|---|---|---|---|---|---|
| Low | High | Low | High | |||||
| SfCOL | 2.05 × 105 | 1.08 × 105 | 3.81 × 105 | - | - | 7 | 7 | 8 |
| VG008 | 4.65 × 105 | 1.91 × 105 | 1.13 × 106 | 0.44 | 0.110 | 29 | 25 | 33 |
| M1 | 4.30 × 104 | 2.51 × 104 | 7.17 × 104 | 4.76 | <0.001 | 4 | 4 | 4 |
| M2 | 5.87 × 104 | 3.49 × 104 | 9.77 × 104 | 3.49 | <0.001 | 4 | 4 | 4 |
| M3 | 1.80 × 104 | 1.01 × 104 | 3.09 × 104 | 11.40 | <0.001 | 3 | 3 | 4 |
| Parameter | Mean | SD | Acceptance Limits | Reference |
|---|---|---|---|---|
| Moisture content (%) | 2.30 | 0.16 | <5 | [6] |
| pH | 7.67 | 0.04 | <8 | |
| Bacteria content (CFU/g) | 2.43 × 106 | 0.07 | <5 × 108 | |
| Mold content (CFU/g) | 7.16 × 105 | 0.12 | ||
| Yeast content (CFU/g) | <1 × 104 | 0 | ||
| Efficacy (%) | 96.7 | 5.8 | ≥80% |
| Parameter | Irradiation Time (h) | Treatments | |||
|---|---|---|---|---|---|
| M3 F | M3 UF | SfCOL F | SfCOL UF | ||
| Efficacy (%) | 0 | 96.7 a | 96.7 a | 100.0 a | 100.0 a |
| 2 | 100.0 a | 6.7 b | 100.0 a | 10.0 b | |
| 4 | 96.7 a | 3.3 b | 100.0 a | 6.7 b | |
| 6 | 100.0 a | 6.7 b | 100.0 a | 13.3 b | |
| OAR (%) | 2 | 100.0 a | 6.9 b | 100.0 a | 10.0 a |
| 4 | 100.0 a | 3.4 b | 100.0 a | 6.7 a | |
| 6 | 100.0 a | 6.9 b | 100.0 a | 13.3 a | |
| Inactivation (%) | 2 | 0 b | 93.1 a | 0 b | 90.0 a |
| 4 | 0 b | 96.6 a | 0 b | 93.3 a | |
| 6 | 0 b | 93.1 a | 0 b | 86.7 a | |
| Treatment | Total Dose OBs/ha | Mortality (%) | Efficacy (%) |
|---|---|---|---|
| Control | - | 9.4 | - |
| M3 Dose 1 | 5.0 × 1010 | 28.6 | 21.2 c |
| M3 Dose 2 | 1.0 × 1011 | 37.8 | 31.3 c |
| M3 Dose 3 | 2.0 × 1011 | 62.6 | 58.7 b |
| M3 Dose 4 | 4.0 × 1011 | 67.4 | 64.0 b |
| M3 Dose 5 | 8.0 × 1011 | 91.5 | 90.6 a |
| SfCOL-F | 1.5 × 1012 | 91.7 | 90.8 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuartas-Otálora, P.E.; Gómez-Valderrama, J.A.; Ramos, A.E.; Barrera-Cubillos, G.P.; Villamizar-Rivero, L.F. Bio-Insecticidal Potential of Nucleopolyhedrovirus and Granulovirus Mixtures to Control the Fall Armyworm Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae). Viruses 2019, 11, 684. https://doi.org/10.3390/v11080684
Cuartas-Otálora PE, Gómez-Valderrama JA, Ramos AE, Barrera-Cubillos GP, Villamizar-Rivero LF. Bio-Insecticidal Potential of Nucleopolyhedrovirus and Granulovirus Mixtures to Control the Fall Armyworm Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae). Viruses. 2019; 11(8):684. https://doi.org/10.3390/v11080684
Chicago/Turabian StyleCuartas-Otálora, Paola E., Juliana A. Gómez-Valderrama, Andrea E. Ramos, Gloria P. Barrera-Cubillos, and Laura F. Villamizar-Rivero. 2019. "Bio-Insecticidal Potential of Nucleopolyhedrovirus and Granulovirus Mixtures to Control the Fall Armyworm Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae)" Viruses 11, no. 8: 684. https://doi.org/10.3390/v11080684
APA StyleCuartas-Otálora, P. E., Gómez-Valderrama, J. A., Ramos, A. E., Barrera-Cubillos, G. P., & Villamizar-Rivero, L. F. (2019). Bio-Insecticidal Potential of Nucleopolyhedrovirus and Granulovirus Mixtures to Control the Fall Armyworm Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae). Viruses, 11(8), 684. https://doi.org/10.3390/v11080684