Detection of Hepatitis E Virus in Shellfish Harvesting Areas from Galicia (Northwestern Spain)

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
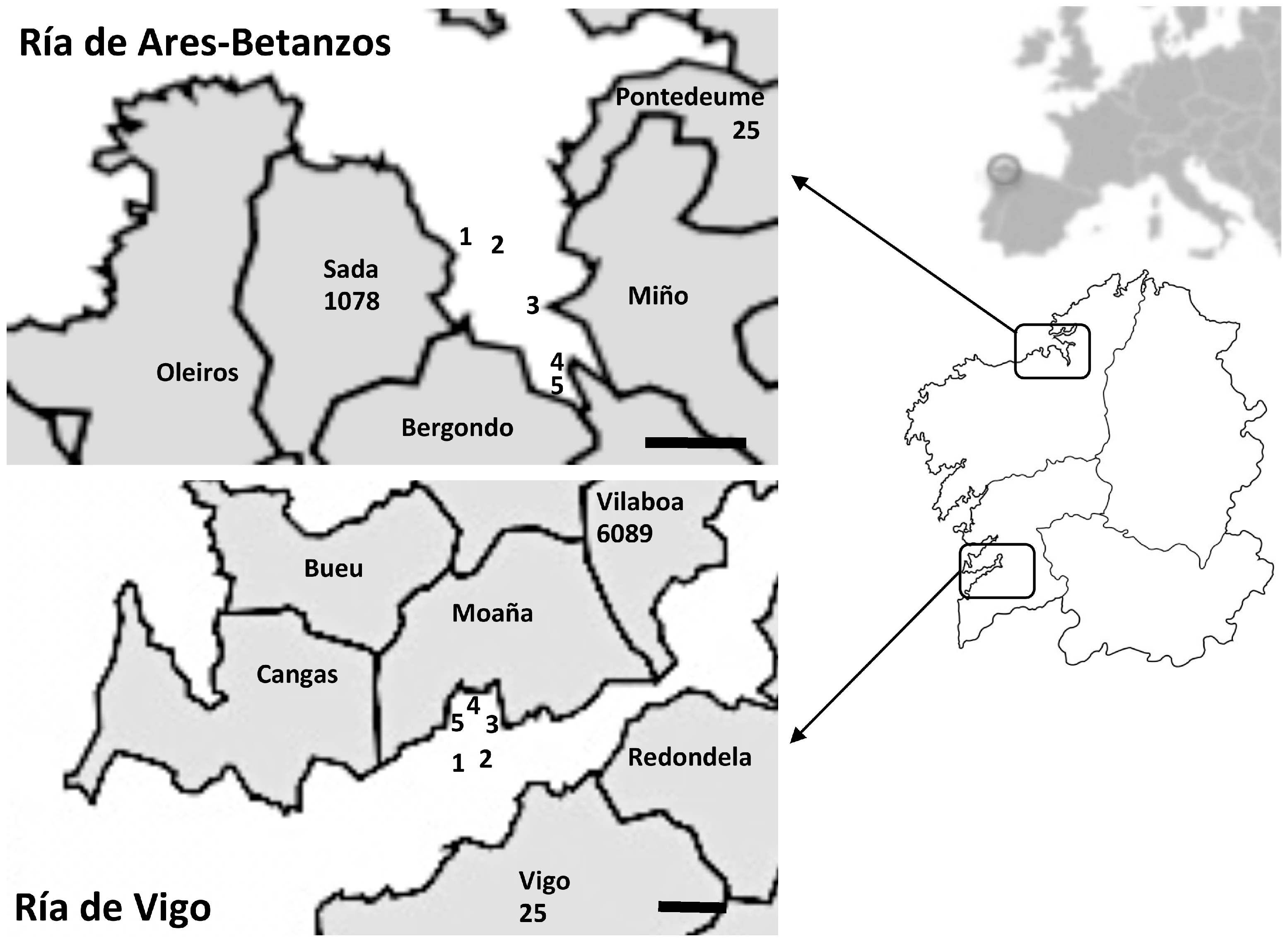
2.1. Shellfish Sampling
2.2. Viral Recovery and RNA Extraction
2.3. RT-qPCR Assay for HEV Screening and Quantification
2.4. Broad-Spectrum Nested RT-PCR Assay
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Zhao, C.; Qi, Y.; Geng, Y. Hepatitis E Virus. In Hepatitis E Virus (Advances in Experimental Medicine and Biology Series 948); Wang, Y., Ed.; Springer: Dordrecht, UK, 2016; pp. 1–16. [Google Scholar]
- World Health Organization (WHO). Hepatitis E. Available online: https://www.who.int/en/news-room/fact-sheets/detail/hepatitis-e (accessed on 21 May 2019).
- Kenney, S.P. The current host range of hepatitis E viruses. Viruses 2019, 11, 452. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Purdy, M.A.; Simmonds, P. Genetic variability and the classification of hepatitis E virus. J. Virol. 2013, 87, 4161–4169. [Google Scholar] [CrossRef] [PubMed]
- Balayan, M.S.; Usmanov, R.K.; Zamyatina, D.I.; Karas, F.R. Brief report: Experimental hepatitis E infection in domestic pigs. J. Med. Virol. 1990, 32, 58–59. [Google Scholar] [CrossRef] [PubMed]
- Reyes, G.R.; Purdy, M.A.; Kim, J.P.; Luk, K.C.; Young, L.M.; Fry, K.E.; Bradley, D.W. Hepatitis E virus (HEV): The novel agent responsible for enterically transmitted non-A, non-B hepatitis. Gastroenterol. Jpn. 1991, 3, 142–147. [Google Scholar] [CrossRef]
- Meng, X.J.; Anderson, D.A.; Arankalle, V.A.; Emerson, S.U.; Harrison, T.J.; Jameel, S.; Okamoto, H. Hepeviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2012; pp. 1021–1028. [Google Scholar]
- Parvez, M.K. The hepatitis E virus nonstructural polyprotein. Future Microbiol. 2017, 12, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Abravanel, F.; Lhomme, S.; Rostaing, L.; Izopet, J. Hepatitis E virus: Chronic infection, extra-hepatic manifestations, and treatment. Clin. Res. Hepatol. Gastroenterol. 2015, 39, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Khuroo, M.S.; Khuroo, M.S.; Khuroo, N.S. Hepatitis E: Discovery, global impact, control and cure. World J. Gastroenterol. 2016, 22, 7030–7045. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, A.R.; Jooma, R.A.; Smego, R.A., Jr. Nosocomial outbreak of hepatitis E infection in Pakistan with possible parenteral transmission. Clin. Infect. Dis. 2005, 40, 908–909. [Google Scholar] [CrossRef]
- Purcell, R.H.; Emerson, S.U. Hepatitis E: An emerging awareness of an old disease. J. Hepatol. 2008, 48, 494–503. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Hepatitis E in the EU/EEA, 2005–2015; ECDC: Stockholm, Sweden, 2017. [CrossRef]
- Lewis, H.C.; Wichmann, O.; Duizer, E. Transmission routes and risk factors for autochthonous hepatitis E virus infection in Europe: A systematic review. Epidemiol. Infect. 2010, 138, 145–166. [Google Scholar] [CrossRef]
- Wyn-Jones, A.P.; Sellwood, J. Enteric viruses in the aquatic environment. J. Appl. Microbiol. 2001, 91, 945–962. [Google Scholar] [CrossRef] [PubMed]
- Barreira, D.M.; Ferreira, M.S.; Fumian, T.M.; Checon, R.; de Sadovsky, A.D.; Leite, J.P.; Miagostovich, M.P.; Spano, L.C. Viral load and genotypes of noroviruses in symptomatic and asymptomatic children in Southeastern Brazil. J. Clin. Virol. 2010, 47, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Loisy, F.; Atmar, R.L.; Le Saux, J.C.; Cohen, J.; Caprais, M.P.; Pommepuy, M.; Le Guyader, F.S. Use of rotavirus virus-like particles as surrogates to evaluate virus persistence in shellfish. Appl. Environ. Microbiol. 2005, 71, 6049–6053. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schwab, K.J.; Neill, F.H.; Estes, M.K.; Metcalf, T.G.; Atmar, R.L. Distribution of Norwalk virus within shellfish following bioaccumulation and subsequent depuration by detection using RT-PCR. J. Food Prot. 1998, 61, 1674–1680. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, T.G.; Melnick, J.L.; Estes, M.K. Environmental virology: From detection of virus in sewage and water by isolation to identification by molecular biology—a trip of over 50 years. Annu. Rev. Microbiol. 1995, 49, 461–487. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.K.; Wong, D.K.; Chung, T.W.; Lim, W.W. Norovirus contamination found in oysters worldwide. J. Med. Virol. 2005, 76, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Bellou, M.; Kokkinos, P.; Vantarakis, A. Shellfish-borne viral outbreaks: A systematic review. Food Environ. Virol. 2013, 5, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Pommepuy, M.; Le Guyader, F.S.; Le Saux, J.C.; Guilfoyle, F.; Doré, B.; Kershaw, S.; Lees, D.; Lowther, J.A.; Morgan, O.C.; Romalde, J.L.; et al. Reducing microbial risk associated with shellfish in European countries. In Improving Seafood Product for the Consumer; Børresen, T., Ed.; Woodhead Publishing Ltd.: Cambridge, UK, 2008; pp. 212–245. [Google Scholar]
- Mesquita, J.R.; Oliveira, D.; Rivadulla, E.; Abreu-Silva, J.; Varela, M.F.; Romalde, J.L.; Nascimento, M.S. Hepatitis E virus genotype 3 in mussels (Mytilus galloprovinciallis), Spain. Food Microbiol. 2016, 58, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. European Regulation (EC) N° 854/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific rules for the organization of official controls on products of animal origin intended for human consumption. Off. J. Eur. Union 2004, 226, 83–127. [Google Scholar]
- Manso, C.F.; Romalde, J.L. Detection and characterization of hepatitis A virus and norovirus in mussels from Galicia (NW Spain). Food Environ. Virol. 2013, 5, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Polo, D.; Varela, M.F.; Romalde, J.L. Detection and quantification of hepatitis A virus and norovirus in Spanish authorized shellfish harvesting areas. Int J. Food Microbiol 2015, 193, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.F.; Polo, D.; Romalde, J.L. Prevalence and genetic diversity of human Sapovirus in shellfish from commercial production areas in Galicia, Spain. Appl. Environ. Microbiol. 2016, 82, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization (ISO). Microbiology of Food Chain—Horizontal Method for Determination of Hepatitis A Virus and Norovirus Using Real-Time RT-PCR—Part 1: Method for Quantification; ISO 15216-1:2017; International Organization for Standardization: Geneva, Switzerland, 2013. [Google Scholar]
- Rivadulla, E.; Varela, M.F.; Romalde, J.L. Low prevalence of Aichi virus in molluscan shellfish samples from Galicia (NW Spain). J. Appl Microbiol. 2017, 122, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Costafreda, M.I.; Bosch, A.; Pintó, R.M. Development, evaluation, and standardization of a real-time TaqMan reverse transcription-PCR assay for quantification of hepatitis A virus in clinical and shellfish samples. Appl. Environ. Microbiol. 2006, 72, 3846–3855. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Pintó, R.M.; Costafreda, M.I.; Bosch, A. Risk assessment in shellfish-borne outbreaks of hepatitis A. Appl. Environ. Microbiol. 2009, 75, 7350–7355. [Google Scholar] [CrossRef] [PubMed]
- Erker, J.C.; Desai, S.M.; Schlauder, G.G.; Dawson, G.J.; Mushahwar, I.K. A hepatitis E virus variant from the United States: Molecular characterization and transmission in cynomolgus macaques. J. Gen. Virol. 1999, 80, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Gao, S.; Li, D.; Zha, E.; Zhou, T.; Wang, S.; Yue, X. Surveillance of hepatitis E virus contamination in shellfish in China. Int. J. Environ. Res. Public Health 2015, 12, 2026–2036. [Google Scholar] [CrossRef]
- Li, T.; Miyamura, T.; Takeda, N. Short report: Detection of hepatitis E virus RNA from the bivalve yamato-shijimi (Corbicula japonica) in Japan. Am. J. Trop. Med. Hyg. 2007, 76, 170–172. [Google Scholar] [CrossRef]
- Inagaki, Y.; Oshiro, Y.; Hasegawa, N.; Fukuda, K.; Abei, M.; Nishi, M.; Okamoto, H.; Ohkohchi, N. Clinical features of hepatitis E virus infection in Ibaraki, Japan: Autochthonous hepatitis E and acute-on-chronic liver failure. J. Exp. Med. 2015, 235, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.J.; Jeong, H.J.; Kim, Y.J.; Lee, S.W.; Lee, J.B.; Park, S.Y.; Song, C.S.; Park, H.M.; Choi, I.S. Analysis of complete genome sequences of swine hepatitis E virus and possible risk factors for transmission of HEV to humans in Korea. J. Med. Virol. 2010, 82, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Namsai, A.; Louisirirotchanakul, S.; Wongchinda, N.; Siripanyaphinyo, U.; Virulhakul, P.; Puthavathana, P.; Myint, K.S.; Gannarong, M.; Ittapong, R. Surveillance of hepatitis A and E viruses contamination in shellfish in Thailand. Lett. Appl. Microbiol. 2011, 53, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, Y.; Isoda, N.; Sato, Y.; Iwaki, T.; Ono, K.; Ido, K.; Sugano, K.; Takahashi, M.; Nishizawa, T.; Okamoto, H. Infection of a Japanese patient by genotype 4 hepatitis E virus while traveling in Vietnam. J. Clin. Microbiol. 2004, 42, 3883–3885. [Google Scholar] [CrossRef] [PubMed]
- Grodzki, M.; Schaeffer, J.; Piquet, J.C.; Le Saux, J.C.; Chevé, J.; Ollivier, J.; Le Pendu, J.; Le Guyader, F.S. Bioaccumulation efficiency, tissue distribution and environmental occurrence of hepatitis E virus in bivalve shellfish from France. Appl. Environ. Microbiol. 2014, 80, 4269–4276. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, G.; Fratini, M.; Spuri Vennarucci, V.; Guercio, A.; Purpari, G.; Muscillo, M. GIV noroviruses and other enteric viruses in bivalves: A preliminary study. New Microbiol. 2012, 35, 27–34. [Google Scholar] [PubMed]
- Krog, J.S.; Larsen, L.E.; Schultz, A.C. Enteric porcine viruses in farmed shellfish in Denmark. Int. J. Food Microbiol. 2014, 186, 105–109. [Google Scholar] [CrossRef]
- La Rosa, G.; Proroga, Y.T.R.; De Medici, D.; Capuano, F.; Iaconelli, M.; Della Libera, S.; Suffredini, E. First detection of hepatitis E virus in shellfish and in seawater from production areas in Southern Italy. Food Environ. Virol. 2018, 10, 127–131. [Google Scholar] [CrossRef]
- Purpari, G.; Macaluso, G.; Di Bella, S.; Gucciardi, F.; Mira, F.; Di Marco, P.; Lastra, A.; Petersen, E.; La Rosa, G.; Guercio, A. Molecular characterization of human enteric viruses in food, water samples, and surface swabs in Sicily. Int. J. Infect. Dis. 2019, 80, 66–72. [Google Scholar] [CrossRef]
- Pol-Hofstad, I.E.; Rutjes, S.A.; Gerssen, A.; Poelman, M.; van der Linden, A.D. Evaluation of the sanitary status of the Dutch shellfish production waters over a 7 year period. In Proceedings of the 9th International Conference on Molluscan Shellfish Safety, Sydney, Australia, 7–22 March 2013; Organising Committee International Conference on Molluscan Shellfish Society: Galway, Ireland, 2014; pp. 52–54. [Google Scholar]
- Diez-Valcarce, M.; Kokkinos, P.; Söderberg, K.; Bouwknegt, M.; Willems, K.; de Roda-Husman, A.M.; von Bonsdorff, C.H.; Bellou, M.; Hernández, M.; Maunula, L.; et al. Occurrence of human enteric viruses in commercial mussels at retail level in three European countries. Food Environ. Virol. 2012, 4, 73–80. [Google Scholar] [CrossRef]
- Crossan, C.; Baker, P.J.; Craft, J.; Takeuchi, Y.; Dalton, H.R.; Scobie, L. Hepatitis E virus genotype 3 in shellfish, United Kingdom. Emerg. Infect. Dis. 2012, 18, 2085–2087. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, Z.; Crossan, C.; Craft, J.; Scobie, L. First report of the presence of hepatitis E virus in Scottish-harvested shellfish purchased at retail level. Food Environ. Virol. 2018, 10, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Said, B.; Ijaz, S.; Kafatos, G.; Booth, L.; Thomas, H.L.; Walsh, A.; Ramsay, M.; Morgan, D. Hepatitis E outbreak on cruise ship. Emerg. Infect. Dis. 2009, 15, 1738–1744. [Google Scholar] [CrossRef] [PubMed]
- Johne, R.; Trojnar, E.; Filter, M.; Hofmann, J. Thermal stability of hepatitis E virus estimated by a cell culture method. Appl. Environ. Microbiol. 2016, 82, 4225–4231. [Google Scholar] [CrossRef] [PubMed]
- Adlhoch, C.; Wolf, A.; Meisel, H.; Kaiser, M.; Ellerbrok, H.; Pauli, G. High HEV presence in four different wild boar populations in East and West Germany. Vet. Microbiol. 2009, 139, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Inoue, J.; Takahashi, M.; Ito, K.; Shimosegawa, T.; Okamoto, H. Analysis of human and swine hepatitis E virus (HEV) isolates of genotype 3 in Japan that are only 81–83 % similar to reported HEV isolates of the same genotype over the entire genome. J. Gen. Virol. 2006, 87, 2363–2369. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Casares, P.; Pina, S.; Buti, M.; Jardi, R.; Martín, M.; Bofill-Mas, S.; Girones, R. Hepatitis E virus epidemiology in industrialized countries. Emerg. Infect. Dis. 2003, 9, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Polo, D.; Feal, X.; Romalde, J.L. Mathematical model for viral depuration kinetics in shellfish: A useful tool to estimate the risk for the consumers. Food Microbiol. 2015, 49, 220–225. [Google Scholar] [CrossRef]
- Hewitt, P.E.; Ijaz, S.; Brailsford, S.R.; Brett, R.; Dicks, S.; Haywood, B.; Kennedy, I.T.; Kitchen, A.; Patel, P.; Poh, J.; et al. Hepatitis E virus in blood components: A prevalence and transmission study in southeast England. Lancet 2014, 384, 1766–1773. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Zone | Sample 1 | 2011 | 2012 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | Jan | Feb | Mar | Apr | May | Jun | |||
| Northe | RC mussels (1) | ◎◼ | ◎❁ | ◎❁ | ◎ | |||||||||||||||
| RC mussels (2) | ◎◆ | ◎◼ | ◎◆ | ◎* | ◎◆★ | |||||||||||||||
| W mussels (3) | ◎ | ◎◼ | ◎◼ | ◎◆ | ◎◆❁ | ◎◆* | ||||||||||||||
| Clams (4) | ◎◼ | ◎◆ | ||||||||||||||||||
| Cockles (5) | ◎◼◆* | |||||||||||||||||||
| South | RC mussels (1) | ◎◼ | NT | ◎◼ | ◎◼ | ◎◼◆ | ◎ | ◎◆ | ||||||||||||
| RC mussels (2) | ◎◼ | NT | ◎◆ | ◎◆ | ◎* | ◎◼◆ | ||||||||||||||
| W mussels (3) | NT | ◎ | ◎ | ◎◼ | ◎* | ◎* | ◎◆★ | |||||||||||||
| Clams (4) | NT | NT | ◎* | ◎* | NT | ◎ | NT | NT | ◎◆★ | |||||||||||
| Cockles (5) | NT | ◎ | ◎* | NT | NT | NT | ||||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivadulla, E.; Varela, M.F.; Mesquita, J.R.; Nascimento, M.S.J.; Romalde, J.L. Detection of Hepatitis E Virus in Shellfish Harvesting Areas from Galicia (Northwestern Spain). Viruses 2019, 11, 618. https://doi.org/10.3390/v11070618
Rivadulla E, Varela MF, Mesquita JR, Nascimento MSJ, Romalde JL. Detection of Hepatitis E Virus in Shellfish Harvesting Areas from Galicia (Northwestern Spain). Viruses. 2019; 11(7):618. https://doi.org/10.3390/v11070618
Chicago/Turabian StyleRivadulla, Enrique, Miguel F. Varela, João R. Mesquita, Maria S.J. Nascimento, and Jesús L. Romalde. 2019. "Detection of Hepatitis E Virus in Shellfish Harvesting Areas from Galicia (Northwestern Spain)" Viruses 11, no. 7: 618. https://doi.org/10.3390/v11070618
APA StyleRivadulla, E., Varela, M. F., Mesquita, J. R., Nascimento, M. S. J., & Romalde, J. L. (2019). Detection of Hepatitis E Virus in Shellfish Harvesting Areas from Galicia (Northwestern Spain). Viruses, 11(7), 618. https://doi.org/10.3390/v11070618