Isolation of an Equine Foamy Virus and Sero-Epidemiology of the Viral Infection in Horses in Japan


Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Virus Isolation
2.2. DNA Extraction
2.3. Polymerase Chain Reaction (PCR) for Detection of the Equine Foamy Virus Genome
2.4. Restriction Enzyme Digestion and Southern Blot Hybridization
2.5. Sequence and Phylogenetic Analyses
2.6. Serum Samples
2.7. Indirect Immunofluorescence Assay (IFA) for Detection of Antibodies to EFV
3. Results
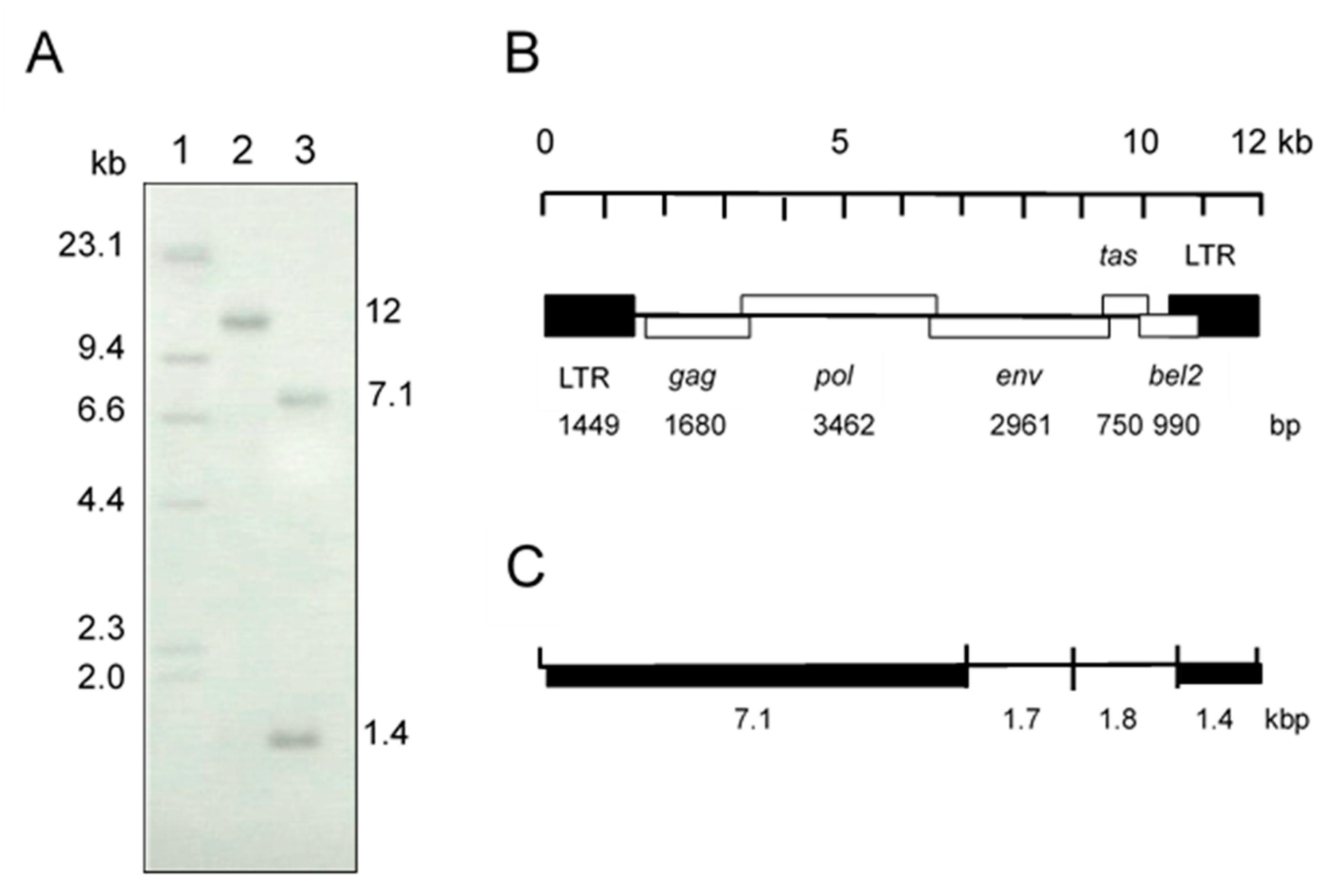
3.1. Virus Isolation and Identification
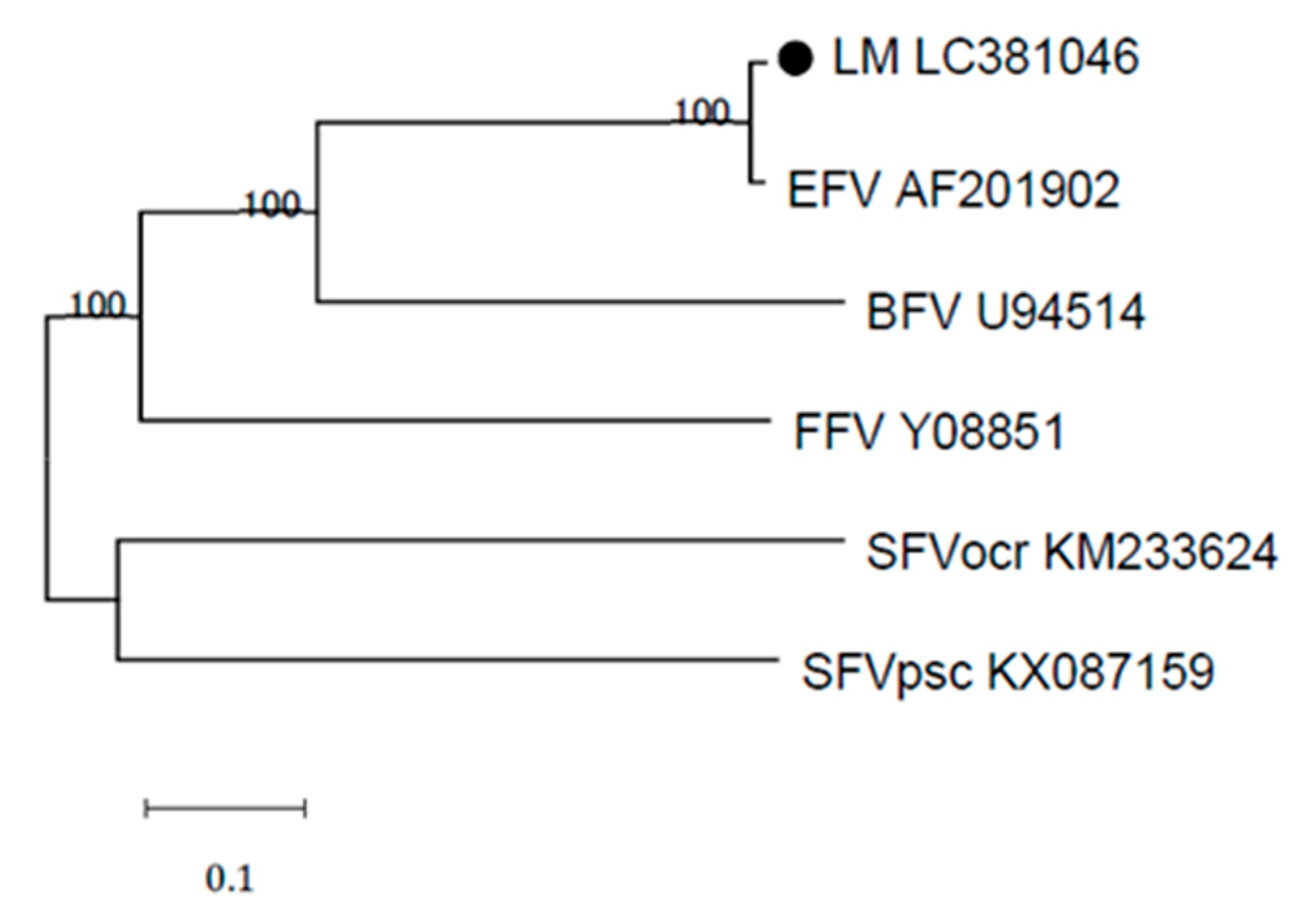
3.2. Sequence Analysis
3.3. Sero-Epizootiology
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Rethwilm, A.; Lindemann, D. Foamy viruses. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P., Eds.; Lippincott, Willilams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1613–1632. [Google Scholar]
- Johnston, P.B. A second immunologic type of simian foamy virus: Monkey throat infections and unmasking by both types. J. Infect. Dis. 1961, 109, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Stiles, G.E.; Bittle, J.L.; Cabasso, V.J. Comparison of simian foamy virus strains including a new serological type. Nature 1964, 201, 1350–1351. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N.G.; Basnight, M.; Gibbs, C.J.; Gajdusek, D.C. Latent viruses in chimpanzees with experimental kuru. Nature 1967, 216, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Achong, B.G.; Mansell, P.W.; Epstein, M.A.; Clifford, P. An unusual virus in cultures from a human nasopharyngeal carcinoma. J. Natl. Cancer Inst. 1971, 46, 299–307. [Google Scholar] [PubMed]
- Riggs, J.L.; Oshirls, L.S.; Taylor, D.O.; Lennette, E.H. Syncytium-forming agent isolated from domestic cats. Nature 1969, 222, 1190–1191. [Google Scholar] [CrossRef] [PubMed]
- Malmquist, W.A.; van der Maaten, M.J.; Boothe, A.D. Isolation, immunodiffusion, immunofluorescence, and electron microscopy of a syncytial virus of lymphosarcomatous and apparently normal cattle. Cancer Res. 1969, 29, 188–200. [Google Scholar] [PubMed]
- Tobaly-Tapiero, J.; Bittoun, P.; Neves, M.; Guillemin, M.C.; Lecellier, C.H.; Puvion-Dutilleul, F.; Gicquel, B.; Zientara, S.; Giron, M.L.; de Thé, H.; et al. Isolation and characterization of an equine foamy virus. J. Virol. 2000, 74, 4064–4073. [Google Scholar] [CrossRef]
- Wu, Z.; Ren, X.; Yang, L.; Hu, Y.; Yang, J.; He, G.; Zhang, J.; Dong, J.; Sun, L.; Du, J.; et al. Virome analysis for identification of novel mammalian viruses in bat species from Chinese provinces. J. Virol. 2012, 86, 10999–11012. [Google Scholar] [CrossRef]
- Meiering, C.D.; Linial, M.L. Historical perspective of foamy virus epidemiology and infection. Clin. Microbiol. Rev. 2001, 14, 165–176. [Google Scholar] [CrossRef]
- Saïb, A. Non-primate foamy viruses. Curr. Top. Microbiol. Immunol. 2003, 277, 197–211. [Google Scholar]
- Linial, M. Why aren’t foamy viruses pathogenic? Trends Microbiol. 2000, 8, 284–289. [Google Scholar] [CrossRef]
- Rethwilm, A.; Bodem, J. Evolution of foamy viruses: The most ancient of all retroviruses. Viruses 2013, 5, 2349–2374. [Google Scholar] [CrossRef] [PubMed]
- Kehl, T.; Tan, J.; Materniak, M. Non-simian foamy viruses: Molecular virology, tropism and prevalence and zoonotic/interspecies transmission. Viruses 2013, 5, 2169–2209. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Miyazawa, T.; Ikeda, Y.; Sato, E.; Nishimura, Y.; Nguyen, N.T.; Takahashi, E.; Mochizuki, M.; Mikami, T. Contrastive prevalence of feline retrovirus infections between northern and southern Vietnam. J. Vet. Med. Sci. 2000, 62, 921–923. [Google Scholar] [CrossRef]
- Bandecchi, P.; Matteucci, D.; Baldinotti, F.; Guidi, G.; Abramo, F.; Tozzini, F.; Bendinelli, M. Prevalence of feline immunodeficiency virus and other retroviral infections in sick cats in Italy. Vet. Immunol. Immunopathol. 1992, 31, 337–345. [Google Scholar] [CrossRef]
- Lin, J.A.; Cheng, M.C.; Inoshima, Y.; Tomonaga, K.; Miyazawa, T.; Tohya, Y.; Toh, K.; Lu, Y.S.; Mikami, T. Seroepidemiological survey of feline retrovirus infections in cats in Taiwan in 1993 and 1994. J. Vet. Med. Sci. 1995, 57, 161–163. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Daniels, M.J.; Golder, M.C.; Jarrett, O.; MacDonald, D.W. Feline viruses in wildcats from Scotland. J. Wildl. Dis. 1999, 35, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Glaus, T.; Hofmann-Lehmann, R.; Greene, C.; Glaus, B.; Wolfensberger, C.; Lutz, H. Seroprevalence of Bartonella henselae infection and correlation with disease status in cats in Switzerland. J. Clin. Microbiol. 1997, 35, 2883–2885. [Google Scholar]
- Winkler, I.G.; Löchelt, M.; Flower, R.L. Epidemiology of feline foamy virus and feline immunodeficiency virus infections in domestic and feral cats: A seroepidemiological study. J. Clin. Microbiol. 1999, 37, 2848–2851. [Google Scholar]
- Mochizuki, M.; Akuzawa, M.; Nagatomo, H. Serological survey of the Iriomote cat (Felis iriomotensis) in Japan. J. Wildl. Dis. 1990, 26, 236–245. [Google Scholar] [CrossRef]
- Miyazawa, T.; Ikeda, Y.; Maeda, K.; Horimoto, T.; Tohya, Y.; Mochizuki, M.; Vu, D.; Vu, G.D.; Cu, D.X.; Ono, K.; et al. Seroepidemiological survey of feline retrovirus infections in domestic and leopard cats in northern Vietnam in 1997. J. Vet. Med. Sci. 1998, 60, 1273–1275. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, R.M.; Pollari, F.L.; McNab, W.B.; Jefferson, B. A serological survey of bovine syncytial virus in Ontario: Associations with bovine leukemia and immunodeficiency-like viruses, production records, and management practices. Can. J. Vet. Res. 1995, 59, 271–278. [Google Scholar] [PubMed]
- Johnson, R.H.; de la Rosa, J.; Abher, I.; Kertayadnya, I.G.; Entwistle, K.W.; Fordyce, G.; Holroyd, R.G. Epidemiological studies of bovine spumavirus. Vet. Microbiol. 1988, 16, 25–33. [Google Scholar] [CrossRef]
- Romen, F.; Backes, P.; Materniak, M.; Sting, R.; Vahlenkamp, T.W.; Riebe, R.; Pawlita, M.; Kuzmak, J.; Löchelt, M. Serological detection systems for identification of cows shedding bovine foamy virus via milk. Virology 2007, 364, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Materniak-Kornas, M.; Osiński, Z.; Rudzki, M.; Kuźmak, J. Development of a recombinant protein-based ELISA for detection of antibodies against bovine foamy virus. J. Vet. Res. 2017, 61, 247–252. [Google Scholar] [CrossRef]
- Kirisawa, R.; Ohmori, H.; Iwai, H.; Kawakami, Y. The genomic diversity among equine herpesvirus-1 strains isolated in Japan. Arch. Virol. 1993, 129, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Kirisawa, R.; Fukuda, T.; Yamanaka, H.; Hagiwara, K.; Goto, M.; Obata, Y.; Yoshino, T.; Iwai, H. Enzymatic amplification and expression of bovine interleukin-1 receptor antagonist cDNA. Vet. Immunol. Immunopathol. 1998, 62, 197–208. [Google Scholar] [CrossRef]
- Southern, E.M. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 1975, 98, 503–517. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tobaly-Tapiero, J.; Bittoun, P.; Saïb, A. Isolation of foamy viruses from peripheral blood lymphocytes. Methods Mol. Biol. 2005, 304, 125–137. [Google Scholar] [PubMed]
- Liebermann, H.; Riebe, R. Isolation of bovine syncytial virus in East Germany. Arch. Exp. Veterinarmed. 1981, 35, 917–919. [Google Scholar] [PubMed]
- Bao, Q.; Hipp, M.; Hugo, A.; Lei, J.; Liu, Y.; Kehl, T.; Hechler, T.; Löchelt, M. In vitro evolution of bovine foamy virus variants with enhanced cell-free virus titers and transmission. Viruses 2015, 7, 5855–5874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, X.; Liang, Z.; Bing, T.; Qiao, W.; Tan, J. The influence of envelope C-terminus amino acid composition on the ratio of cell-free to cell-cell transmission for bovine foamy virus. Viruses 2019, 11, 130. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, Y.; Kimura, K.; Oguma, K.; Ono, M.; Horikita, T.; Sentsui, H. Isolation of bovine foamy virus in Japan. J. Vet. Med. Sci. 2018, 80, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Goepfert, P.A.; Shaw, K.L.; Ritter, G.D., Jr.; Mulligan, M.J. A sorting motif localizes the foamy virus glycoprotein to the endoplasmic reticulum. J. Virol. 1997, 71, 778–784. [Google Scholar] [PubMed]
- Goepfert, P.A.; Shaw, K.; Wang, G.; Bansal, A.; Edwards, B.H.; Mulligan, M.J. An endoplasmic reticulum retrieval signal partitions human foamy virus maturation to intracytoplasmic membranes. J. Virol. 1999, 73, 7210–7217. [Google Scholar]
- Phung, H.T.; Ikeda, Y.; Miyazawa, T.; Nakamura, K.; Mochizuki, M.; Izumiya, Y.; Sato, E.; Nishimura, Y.; Tohya, Y.; Takahashi, E.; et al. Genetic analyses of feline foamy virus isolates from domestic and wild feline species in geographically distinct areas. Virus Res. 2001, 76, 171–181. [Google Scholar] [CrossRef]
- Flower, R.L.; Wilcox, G.E.; Cook, R.D.; Ellis, T.M. Detection and prevalence of serotypes of feline syncytial spumaviruses. Arch. Virol. 1985, 83, 53–63. [Google Scholar] [CrossRef]
- Winkler, I.G.; Flügel, R.M.; Löchelt, M.; Flower, R.L. Detection and molecular characterisation of feline foamy virus serotypes in naturally infected cats. Virology 1998, 247, 144–151. [Google Scholar] [CrossRef]
- Hechler, T.; Materniak, M.; Kehl, T.; Kuzmak, J.; Löchelt, M. Complete genome sequences of two novel European clade bovine foamy viruses from Germany and Poland. J. Virol. 2012, 86, 10905–10906. [Google Scholar] [CrossRef] [PubMed]
- Materniak, M.; Kuzmak, J. Occurrence of equine foamy virus infection in horses from Poland. In Proceedings of the 9th International Foamy Conference, Bethesda, MD, USA, 29–30 May 2012. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5’–3’) | Location 1 |
|---|---|---|
| LTR-F | TGTCATGGAATGAGGATCCAG | 1–21 |
| 10587–10607 | ||
| LTR-R | ATTGTCGCGGTATCTCCTTAA | 1449–1429 |
| 12035–12015 | ||
| Gag-F | AGATACCGCGACAATTGGCG | 1435–1454 |
| Gag-R3 | CCATTGTCCCGAGGTAAATC | 1637–1618 |
| Gag-F3 | AAGAAGAGGCCCTGGAAGAA | 3071–3090 |
| Gag-R | AGGGACACAAGTTATTTCAGCTC | 3246–3224 |
| Pol-F | GGCGTTATTGAAGGCATTTG | 4670–4690 |
| Pol-R | CCCATACCTGCTGAATGTTG | 4997–4978 |
| Env-F | ATGACACCTCCAATGACTCTAC | 6536–6557 |
| Env-R3 | CAGGTATGGCCTCTCTGAT | 6617–6599 |
| Env-R | TTATTCTCCTTTGTCCTCTC | 9496–9477 |
| Env-F2 | TTTGGGTAAAGTACCAGCCTC | 8113–8133 |
| Env-R2 | GGATAAGTCCACTTCCCAGAG | 9528–9508 |
| TAS-F | AGGATATTATCATGGCTAGCA | 9431–9451 |
| TAS-R | ATGGTTCTCGAATAAAGCGGT | 11885–11865 |
| 1299–1279 |
| Primer Pair | Amplified Region 1 | PCR Product Size (bp) |
|---|---|---|
| LTR-F & LTR-R | 1–1429, 10587–12035 | 1429 |
| LTR-F & Gag-R3 | 1–1637 | 1637 |
| Gag-F & Gag-R | 1435–3246 | 1812 |
| Gag-F3 & Pol-R | 3071–4997 | 1927 |
| Pol-F & Env-R3 | 4670–6617 | 1948 |
| Env-F & Env-R | 6536–9496 | 2961 |
| Env-F2 & Env-R2 | 8113–9528 | 1416 |
| TAS-F & TAS-R | 9431–11885 | 2454 |
| Serum Collection Date | ||||
|---|---|---|---|---|
| 26 Jan, 2000 | 11 Oct, 2000 | 14 Feb, 2001 | 1 Oct, 2001 1 | |
| IFA titer | 160 | 160 | 160 | 80 |
| Identity (%) | ||
|---|---|---|
| Region | Nucleotide Sequence | Amino Acid Sequence |
| LTR | 98.2 (1449) 1 | |
| gag | 98.6 (1680) | 99.1 (559) 2 |
| pol | 98.6 (3462) | 99.1 (1153) |
| env | 98.3 (2961) | 98.5 (986) |
| tas | 99.1 (750) | 100.0 (249) |
| bel2 | 97.2 (990) | 97.3 (329) |
| In 2001 to 2002 | In 2014 to 2016 | ||||
|---|---|---|---|---|---|
| Farm | Category | Collection Date | Positive/Negative/Tested Sera | Collection Date | Positive/Negative/Tested Sera |
| A | Stallion | 7 Jun, 2002 | 3 (8.0y) 1/ 25 (9.6y)/ | 30 Jun, 2015 | 18 (13.7y)/ 26 (9.7y)/ |
| 28 (9.1y) 2 (10.7%) 3 | 44 (11.3y) (40.9%) | ||||
| B | Stallion | 15 May, 2015 | 2 (10.0y)/ 13 (10.8y)/ | ||
| 15 (10.7y) (13.3%) | |||||
| C | Broodmare | 13 Jun, 2002 | 18 (9.6y)/ 54 (9.4y)/ | 30 Jun, 2015 | 38 (12.0y)/ 69 (9.9y)/ |
| 72 (9.4y) (25.0%) | 107 (10.6y) (35.5%) | ||||
| D | Broodmare | 17 Apr, 2001 | 7 (11.0y)/ 18 (8.6y)/ | 26 Jan, 2015 | 7 (11.7y)/ 18 (9.4y)/ |
| 25 (9.2y) (28.0%) | 25 (10.0y) (28.0%) | ||||
| E | Broodmare | 30 Jun, 2002 | 6 (12.7y)/ 23 (10.7y)/ | ||
| 29 (11.1y) (20.7%) | |||||
| F | Broodmare | 30 Jun, 2002 | 3 (8.7y)/ 9 (9.7y)/ | ||
| 12 (9.4y) (25.0%) | |||||
| G | Broodmare | 11 Mar, 2015 | 5 (14.0y)/ 25 (11.0y)/ | ||
| 30 (11.5y) (16.7%) | |||||
| H | Broodmare | 17 Jan, 2014 | 5 (11.4y)/ 34 (10.8y)/ | ||
| 39 (10.9y) (12.8%) | |||||
| I | Broodmare | 28 Jan, 2016 | 3 (11.3y)/ 19 (9.3y)/ | ||
| 22 (9.6y) (13.6%) | |||||
| J | Broodmare | 31 Oct, 2016 | 2 (11.5y)/ 9 (12.2y)/ | ||
| 11 (12.1y) (18.2%) | |||||
| Subtotal | Stallion | 3 (8.0y)/ 25 (9.6y) | 20 (13.3y)/ 39 (10.1y)/ | ||
| 28 (9.1y) (10.7%) | 59 (11.2y) (33.9%) | ||||
| Broodmare | 34 (10.3y)/ 104 (9.6y) | 60 (12.0y)/174 (10.2y)/ | |||
| 138 (9.8y) (24.6%) | 234 (10.7y) (25.6%) | ||||
| Total | 37/166 4 (22.3%) | 80/293 (27.3%) | |||
| Age (years) | Farm A | Farm C | ||
|---|---|---|---|---|
| 2002 | 2015 | 2002 | 2015 | |
| Positive/Tested | Positive/Tested | Positive/Tested | Positive/Tested | |
| 4 | 1/2 (50.0%) 1 | 1/1 (100.0%) | 0/2 | |
| 5 | 0/2 | 0/7 | 0/3 | |
| 6 | 0/1 | 0/7 | 2/7 (28.6%) | 1/5 (20.0%) |
| 7 | 0/5 | 2/5 (40.0%) | 4/9 (44.4%) | 2/11 (18.2%) |
| 8 | 0/3 | 2/6 (33.3%) | 1/4 (25.0%) | 3/13 (23.1%) |
| 9 | 0/4 | 1/3 (33.3%) | 2/5 (40.0%) | 2/9 (22.2%) |
| 10 | 2/3 (66.7%) | 1/3 (33.3%) | 2/10 (20.0%) | 6/19 (31.6%) |
| 11 | 0/2 | 0/1 | 2/11 (18.2%) | 4/10 (40.0%) |
| 12 | 0/1 | 0/1 | 3/6 (50.0%) | 5/9 (55.6%) |
| 13 | 0/1 | 1/2 (50.0%) | 1/2 (50.0%) | 2/4 (50.0%) |
| 14 | 0/1 | 0/3 | 0/4 | 4/8 (50.0%) |
| 15 | 0/1 | 2/2 (100.0%) | 1/2 (50.0%) | 3/7 (42.9%) |
| 16 | 0/1 | 3/3 (100.0%) | 0/1 | 1/2 (50.0%) |
| 17 | 2/3 3 (66.7%) | 0/2 | 4/5 6 (80.0%) | |
| 18 | 1/1 6 (100.0%) | |||
| 19 | 0/1 6 | |||
| 20 | ||||
| 21 | ||||
| 22 | 1/1 4 (100.0%) | |||
| 23 | 0/1 | 2/2 5 (100.0%) | ||
| 24 | 0/1 6 | |||
| Subtotal 4y to 9y | 1/17 (5.9%) | 6/22 (27.3%) | 9/34 (26.5%) | 8/41 (19.5%) |
| 10y to 14y | 2/8 (25.0%) | 2/10 (20.0%) | 8/33 (24.2%) | 21/50 (42.0%) |
| 15y to 24y | 0/3 | 10/12 (83.3%) | 1/5 (20.0%) | 9/16 (56.3%) |
| total | 3/28 (10.7%) | 18/44 (40.9%) | 18/72 (25.0%) | 38/107 (35.5%) |
| Average age | 8.0y/ 9.6y/ 9.1y 2 | 13.7y/ 9.7y/ 11.3y | 9.6y/ 9.4y/ 9.4y | 12.0y/ 9.9y/ 10.6y |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirisawa, R.; Toishi, Y.; Hashimoto, H.; Tsunoda, N. Isolation of an Equine Foamy Virus and Sero-Epidemiology of the Viral Infection in Horses in Japan. Viruses 2019, 11, 613. https://doi.org/10.3390/v11070613
Kirisawa R, Toishi Y, Hashimoto H, Tsunoda N. Isolation of an Equine Foamy Virus and Sero-Epidemiology of the Viral Infection in Horses in Japan. Viruses. 2019; 11(7):613. https://doi.org/10.3390/v11070613
Chicago/Turabian StyleKirisawa, Rikio, Yuko Toishi, Hiromitsu Hashimoto, and Nobuo Tsunoda. 2019. "Isolation of an Equine Foamy Virus and Sero-Epidemiology of the Viral Infection in Horses in Japan" Viruses 11, no. 7: 613. https://doi.org/10.3390/v11070613
APA StyleKirisawa, R., Toishi, Y., Hashimoto, H., & Tsunoda, N. (2019). Isolation of an Equine Foamy Virus and Sero-Epidemiology of the Viral Infection in Horses in Japan. Viruses, 11(7), 613. https://doi.org/10.3390/v11070613