Interaction of Structural Glycoprotein E2 of Classical Swine Fever Virus with Protein Phosphatase 1 Catalytic Subunit Beta (PPP1CB)



Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Yeast Two-Hybrid Screening
2.3. Immunoblotting and Antibodies
2.4. Immunoprecipitation
2.5. Proximity Ligation Assay
2.6. PP1 Activating and Inhibitory Experiments
2.7. siRNA Experiments
3. Results
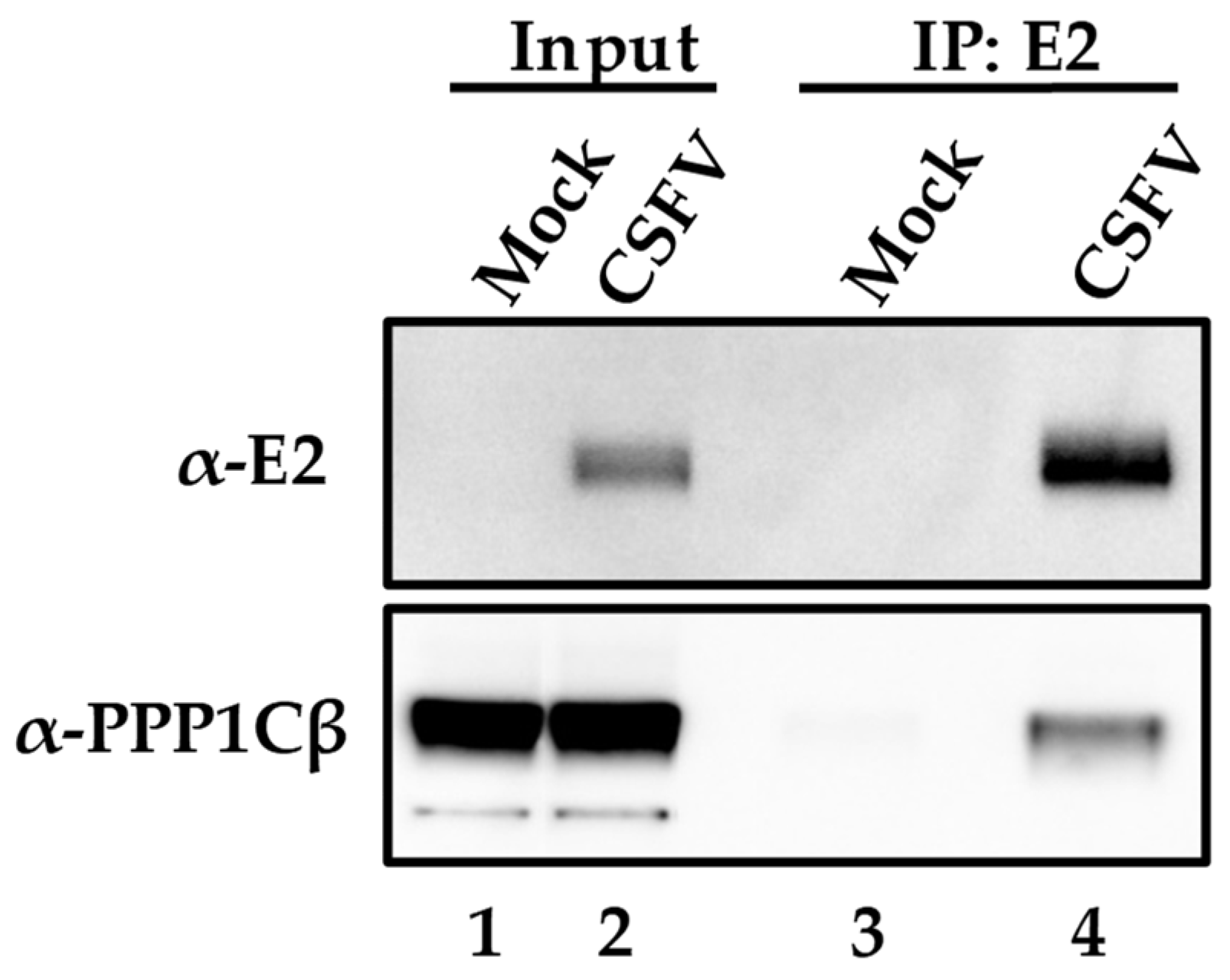
3.1. CSFV Structural E2 Protein Binds Swine PPP1CB
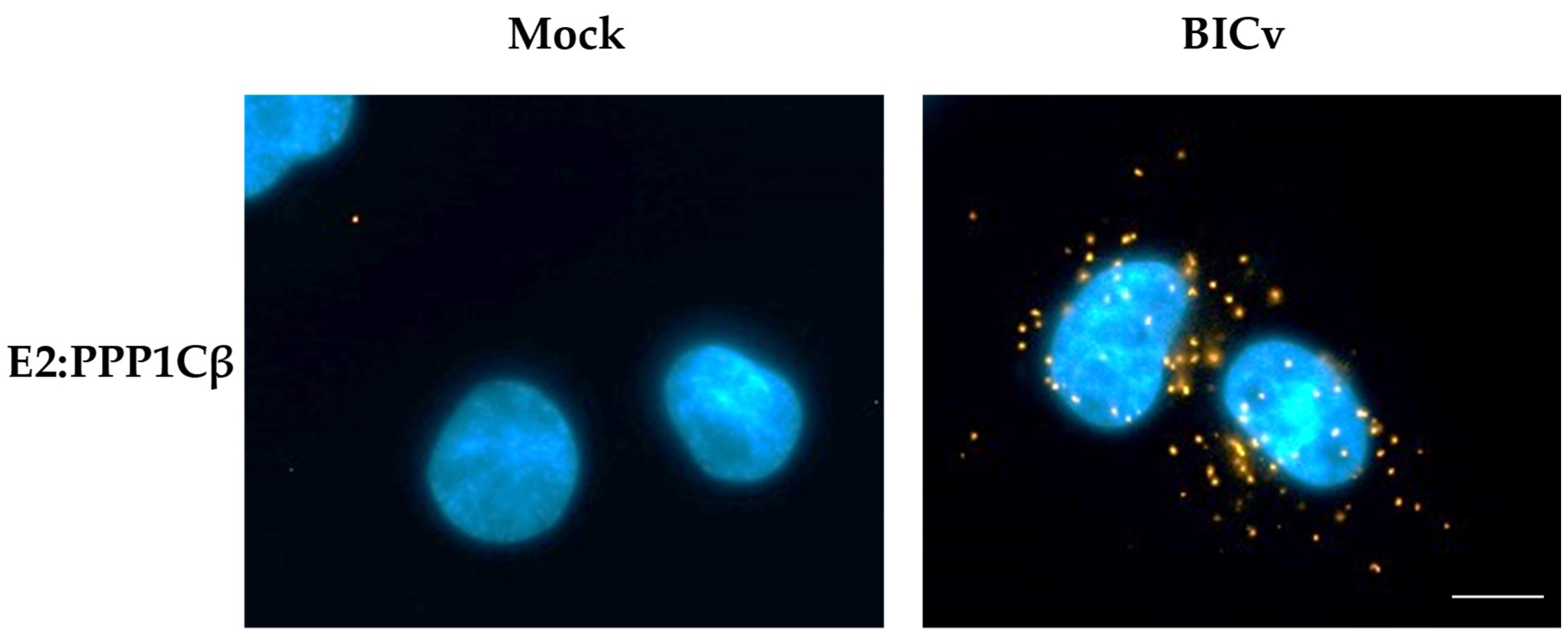
3.2. CSFV E2 Protein Interacts with Host PPP1CB during the Virus Infectious Cycle
3.3. CSFV Is Transiently Phosphorylated during Virus Infection
3.4. Pharmacological Activation of PP1 Inhibits CSFV Replication
3.5. siRNA Knockdown of Cellular PPP1CB
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nishi, T.; Kameyama, K.I.; Kato, T.; Fukai, K. Genome Sequence of a Classical Swine Fever Virus of Subgenotype 2.1, Isolated from a Pig in Japan in 2018. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Fauquet, C.M.; Fargette, D. International Committee on Taxonomy of Viruses and the 3,142 unassigned species. Virol. J. 2005, 2, 64. [Google Scholar] [CrossRef] [PubMed]
- Rice, C.M. Flaviviridae: The Viruses and Their Replication. In Fundamental Virology, 3rd ed.; Fields, B.N., Knipe, D.M., Howley, P.M., Eds.; Lippincott Raven: Philadelphia, PA, USA, 1996; pp. 931–959. [Google Scholar]
- Meyers, G.; Saalmuller, A.; Buttner, M. Mutations abrogating the RNase activity in glycoprotein E(rns) of the pestivirus classical swine fever virus lead to virus attenuation. J. Virol. 1999, 73, 10224–10235. [Google Scholar]
- Risatti, G.R.; Borca, M.V.; Kutish, G.F.; Lu, Z.; Holinka, L.G.; French, R.A.; Tulman, E.R.; Rock, D.L. The E2 glycoprotein of classical swine fever virus is a virulence determinant in swine. J. Virol. 2005, 79, 3787–3796. [Google Scholar] [CrossRef] [PubMed]
- Risatti, G.R.; Holinka, L.G.; Carrillo, C.; Kutish, G.F.; Lu, Z.; Tulman, E.R.; Sainz, I.F.; Borca, M.V. Identification of a novel virulence determinant within the E2 structural glycoprotein of classical swine fever virus. Virology 2006, 355, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Risatti, G.R.; Holinka, L.G.; Fernandez Sainz, I.; Carrillo, C.; Kutish, G.F.; Lu, Z.; Zhu, J.; Rock, D.L.; Borca, M.V. Mutations in the carboxyl terminal region of E2 glycoprotein of classical swine fever virus are responsible for viral attenuation in swine. Virology 2007, 364, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Risatti, G.R.; Holinka, L.G.; Fernandez Sainz, I.; Carrillo, C.; Lu, Z.; Borca, M.V. N-linked glycosylation status of classical swine fever virus strain Brescia E2 glycoprotein influences virulence in swine. J. Virol. 2007, 81, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Risatti, G.R.; Holinka, L.G.; Lu, Z.; Kutish, G.F.; Tulman, E.R.; French, R.A.; Sur, J.H.; Rock, D.L.; Borca, M.V. Mutation of E1 glycoprotein of classical swine fever virus affects viral virulence in swine. Virology 2005, 343, 116–127. [Google Scholar] [CrossRef] [PubMed]
- van Rijn, P.A.; Miedema, G.K.; Wensvoort, G.; van Gennip, H.G.; Moormann, R.J. Antigenic structure of envelope glycoprotein E1 of hog cholera virus. J. Virol. 1994, 68, 3934–3942. [Google Scholar] [PubMed]
- Borca, M.V.; Holinka, L.G.; Ramirez-Medina, E.; Risatti, G.R.; Vuono, E.A.; Berggren, K.A.; Gladue, D.P. Identification of structural glycoprotein E2 domain critical to mediate replication of Classical Swine Fever Virus in SK6 cells. Virology 2019, 526, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Dong, H.; Li, S.; Munir, M.; Chen, J.; Luo, Y.; Sun, Y.; Liu, L.; Qiu, H.J. Hemoglobin subunit beta interacts with the capsid protein and antagonizes the growth of classical swine fever virus. J. Virol. 2013, 87, 5707–5717. [Google Scholar] [CrossRef] [PubMed]
- Gladue, D.P.; Holinka, L.G.; Fernandez-Sainz, I.J.; Prarat, M.V.; O’Donell, V.; Vepkhvadze, N.; Lu, Z.; Rogers, K.; Risatti, G.R.; Borca, M.V. Effects of the interactions of classical swine fever virus Core protein with proteins of the SUMOylation pathway on virulence in swine. Virology 2010, 407, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Gladue, D.P.; Holinka, L.G.; Fernandez-Sainz, I.J.; Prarat, M.V.; O’Donnell, V.; Vepkhvadze, N.G.; Lu, Z.; Risatti, G.R.; Borca, M.V. Interaction between Core protein of classical swine fever virus with cellular IQGAP1 protein appears essential for virulence in swine. Virology 2011, 412, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Gladue, D.P.; O’Donnell, V.; Fernandez-Sainz, I.J.; Fletcher, P.; Baker-Branstetter, R.; Holinka, L.G.; Sanford, B.; Carlson, J.; Lu, Z.; Borca, M.V. Interaction of structural core protein of classical swine fever virus with endoplasmic reticulum-associated degradation pathway protein OS9. Virology 2014, 460–461, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; He, W.R.; Shen, L.; Dong, H.; Yu, J.; Wang, X.; Yu, S.; Li, Y.; Li, S.; Luo, Y.; et al. The laminin receptor is a cellular attachment receptor for classical Swine Fever virus. J. Virol. 2015, 89, 4894–4906. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Ling, L.; Liao, Y.; Li, S.; Han, W.; Zhao, B.; Sun, Y.; Qiu, H.J. Beta-actin interacts with the E2 protein and is involved in the early replication of classical swine fever virus. Virus Res. 2014, 179, 161–168. [Google Scholar] [CrossRef]
- Yang, Z.; Shi, Z.; Guo, H.; Qu, H.; Zhang, Y.; Tu, C. Annexin 2 is a host protein binding to classical swine fever virus E2 glycoprotein and promoting viral growth in PK-15 cells. Virus Res. 2015, 201, 16–23. [Google Scholar] [CrossRef]
- Li, S.; Wang, J.; He, W.R.; Feng, S.; Li, Y.; Wang, X.; Liao, Y.; Qin, H.Y.; Li, L.F.; Dong, H.; et al. Thioredoxin 2 Is a Novel E2-Interacting Protein That Inhibits the Replication of Classical Swine Fever Virus. J. Virol. 2015, 89, 8510–8524. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, S.; Liao, Y.; Zhang, E.; Feng, S.; Yu, S.; Li, L.F.; He, W.R.; Li, Y.; Luo, Y.; et al. Mitogen-activated Protein Kinase Kinase 2 (MEK2), a Novel E2-interacting Protein, Promotes the Growth of Classical Swine Fever Virus via Attenuation of the JAK-STAT Signaling Pathway. J. Virol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Gladue, D.P.; Baker-Bransetter, R.; Holinka, L.G.; Fernandez-Sainz, I.J.; O’Donnell, V.; Fletcher, P.; Lu, Z.; Borca, M.V. Interaction of CSFV E2 protein with swine host factors as detected by yeast two-hybrid system. PLoS ONE 2014, 9, e85324. [Google Scholar] [CrossRef] [PubMed]
- Bollen, M. Combinatorial control of protein phosphatase-1. Trends Biochem. Sci. 2001, 26, 426–431. [Google Scholar] [CrossRef]
- Wakula, P.; Beullens, M.; Ceulemans, H.; Stalmans, W.; Bollen, M. Degeneracy and function of the ubiquitous RVXF motif that mediates binding to protein phosphatase-1. J. Biol. Chem. 2003, 278, 18817–18823. [Google Scholar] [CrossRef]
- Cougot, D.; Allemand, E.; Riviere, L.; Benhenda, S.; Duroure, K.; Levillayer, F.; Muchardt, C.; Buendia, M.A.; Neuveut, C. Inhibition of PP1 phosphatase activity by HBx: a mechanism for the activation of hepatitis B virus transcription. Sci. Signal 2012, 5, ra1. [Google Scholar] [CrossRef]
- Mesman, A.W.; Zijlstra-Willems, E.M.; Kaptein, T.M.; de Swart, R.L.; Davis, M.E.; Ludlow, M.; Duprex, W.P.; Gack, M.U.; Gringhuis, S.I.; Geijtenbeek, T.B. Measles virus suppresses RIG-I-like receptor activation in dendritic cells via DC-SIGN-mediated inhibition of PP1 phosphatases. Cell Host Microbe 2014, 16, 31–42. [Google Scholar] [CrossRef]
- Davis, M.E.; Wang, M.K.; Rennick, L.J.; Full, F.; Gableske, S.; Mesman, A.W.; Gringhuis, S.I.; Geijtenbeek, T.B.; Duprex, W.P.; Gack, M.U. Antagonism of the phosphatase PP1 by the measles virus V protein is required for innate immune escape of MDA5. Cell Host Microbe 2014, 16, 19–30. [Google Scholar] [CrossRef]
- Richard, C.A.; Rincheval, V.; Lassoued, S.; Fix, J.; Cardone, C.; Esneau, C.; Nekhai, S.; Galloux, M.; Rameix-Welti, M.A.; Sizun, C.; et al. RSV hijacks cellular protein phosphatase 1 to regulate M2-1 phosphorylation and viral transcription. PLoS Pathog. 2018, 14, e1006920. [Google Scholar] [CrossRef]
- Ilinykh, P.A.; Tigabu, B.; Ivanov, A.; Ammosova, T.; Obukhov, Y.; Garron, T.; Kumari, N.; Kovalskyy, D.; Platonov, M.O.; Naumchik, V.S.; et al. Role of protein phosphatase 1 in dephosphorylation of Ebola virus VP30 protein and its targeting for the inhibition of viral transcription. J. Biol. Chem. 2014, 289, 22723–22738. [Google Scholar] [CrossRef]
- Ammosova, T.; Jerebtsova, M.; Beullens, M.; Lesage, B.; Jackson, A.; Kashanchi, F.; Southerland, W.; Gordeuk, V.R.; Bollen, M.; Nekhai, S. Nuclear targeting of protein phosphatase-1 by HIV-1 Tat protein. J. Biol. Chem. 2005, 280, 36364–36371. [Google Scholar] [CrossRef]
- Zsak, L.; Lu, Z.; Kutish, G.F.; Neilan, J.G.; Rock, D.L. An African swine fever virus virulence-associated gene NL-S with similarity to the herpes simplex virus ICP34.5 gene. J. Virol. 1996, 70, 8865–8871. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Edwards, S.; Moennig, V.; Wensvoort, G. The development of an international reference panel of monoclonal antibodies for the differentiation of hog cholera virus from other pestiviruses. Vet. Microbiol. 1991, 29, 101–108. [Google Scholar] [CrossRef]
- Soderberg, O.; Gullberg, M.; Jarvius, M.; Ridderstrale, K.; Leuchowius, K.J.; Jarvius, J.; Wester, K.; Hydbring, P.; Bahram, F.; Larsson, L.G.; et al. Direct observation of individual endogenous protein complexes in situ by proximity ligation. Nat. Methods 2006, 3, 995–1000. [Google Scholar] [CrossRef]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef]
- Gullberg, M.; Andersson, A.-C. Highly specific detection of phosphorylated proteins by Duolink. Nat. Methods 2009, 6, 686. [Google Scholar] [CrossRef]
- Chalfant, C.E.; Kishikawa, K.; Mumby, M.C.; Kamibayashi, C.; Bielawska, A.; Hannun, Y.A. Long chain ceramides activate protein phosphatase-1 and protein phosphatase-2A. Activation is stereospecific and regulated by phosphatidic acid. J. Biol. Chem. 1999, 274, 20313–20317. [Google Scholar] [CrossRef]
- Zhang, S.; Sun, Y.; Chen, H.; Dai, Y.; Zhan, Y.; Yu, S.; Qiu, X.; Tan, L.; Song, C.; Ding, C. Activation of the PKR/eIF2alpha signaling cascade inhibits replication of Newcastle disease virus. Virol. J. 2014, 11, 62. [Google Scholar] [CrossRef] [PubMed]
- Bollen, M.; Peti, W.; Ragusa, M.J.; Beullens, M. The extended PP1 toolkit: designed to create specificity. Trends Biochem. Sci. 2010, 35, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, D.; Feng, J.; Liu, Z.; Hu, Y.; Liu, C.; Kong, X. HIV-1 Protein Tat1-72 Impairs Neuronal Dendrites via Activation of PP1 and Regulation of the CREB/BDNF Pathway. Virol. Sin. 2018, 33, 261–269. [Google Scholar] [CrossRef]
- Baer, A.; Shafagati, N.; Benedict, A.; Ammosova, T.; Ivanov, A.; Hakami, R.M.; Terasaki, K.; Makino, S.; Nekhai, S.; Kehn-Hall, K. Protein Phosphatase-1 regulates Rift Valley fever virus replication. Antiviral Res. 2016, 127, 79–89. [Google Scholar] [CrossRef]
- Husser, L.; Alves, M.P.; Ruggli, N.; Summerfield, A. Identification of the role of RIG-I, MDA-5 and TLR3 in sensing RNA viruses in porcine epithelial cells using lentivirus-driven RNA interference. Virus Res. 2011, 159, 9–16. [Google Scholar] [CrossRef]
- Li, L.F.; Yu, J.; Zhang, Y.; Yang, Q.; Li, Y.; Zhang, L.; Wang, J.; Li, S.; Luo, Y.; Sun, Y.; et al. Interferon-Inducible Oligoadenylate Synthetase-Like Protein Acts as an Antiviral Effector against Classical Swine Fever Virus via the MDA5-Mediated Type I Interferon-Signaling Pathway. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E2-BD | LAM-BD | BD | |
|---|---|---|---|
| PPP1CB-AD | +++ | - | - |
| AD | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vuono, E.A.; Ramirez-Medina, E.; Holinka, L.G.; Baker-Branstetter, R.; Borca, M.V.; Gladue, D.P. Interaction of Structural Glycoprotein E2 of Classical Swine Fever Virus with Protein Phosphatase 1 Catalytic Subunit Beta (PPP1CB). Viruses 2019, 11, 307. https://doi.org/10.3390/v11040307
Vuono EA, Ramirez-Medina E, Holinka LG, Baker-Branstetter R, Borca MV, Gladue DP. Interaction of Structural Glycoprotein E2 of Classical Swine Fever Virus with Protein Phosphatase 1 Catalytic Subunit Beta (PPP1CB). Viruses. 2019; 11(4):307. https://doi.org/10.3390/v11040307
Chicago/Turabian StyleVuono, Elizabeth A., Elizabeth Ramirez-Medina, Lauren G. Holinka, Ryan Baker-Branstetter, Manuel V. Borca, and Douglas P. Gladue. 2019. "Interaction of Structural Glycoprotein E2 of Classical Swine Fever Virus with Protein Phosphatase 1 Catalytic Subunit Beta (PPP1CB)" Viruses 11, no. 4: 307. https://doi.org/10.3390/v11040307
APA StyleVuono, E. A., Ramirez-Medina, E., Holinka, L. G., Baker-Branstetter, R., Borca, M. V., & Gladue, D. P. (2019). Interaction of Structural Glycoprotein E2 of Classical Swine Fever Virus with Protein Phosphatase 1 Catalytic Subunit Beta (PPP1CB). Viruses, 11(4), 307. https://doi.org/10.3390/v11040307