Early Multiphasic HBV Infection Initiation Kinetics Is Not Clone-Specific and Is Not Affected by Hepatitis D Virus (HDV) Infection

 , and
, and 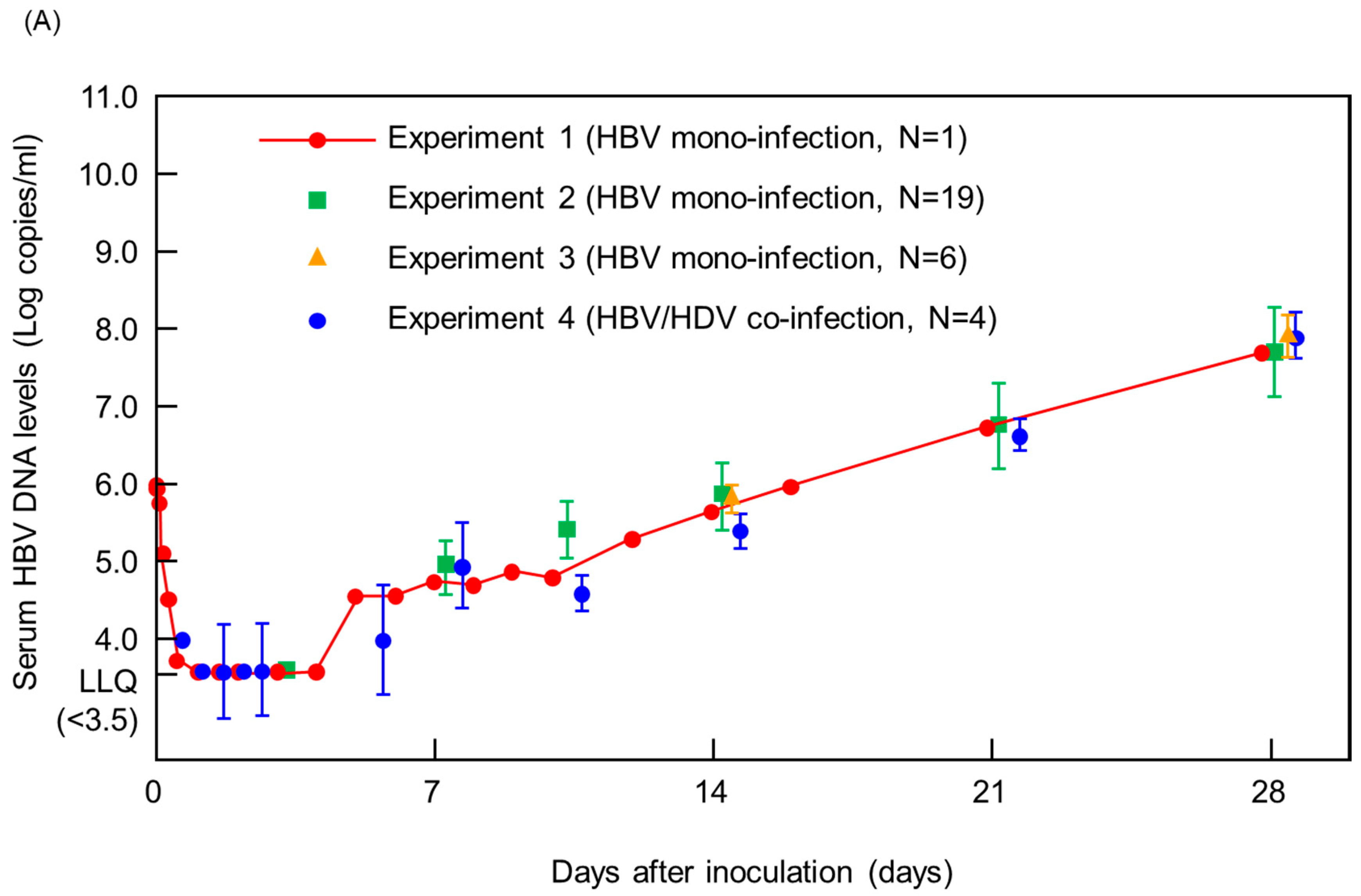{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the Viral Inocula
2.2. Human Hepatocyte Chimeric Mouse Experiments
2.3. Quantification of Serum HBV DNA
2.4. Quantification of serum HDV RNA
2.5. Statistical Analyses
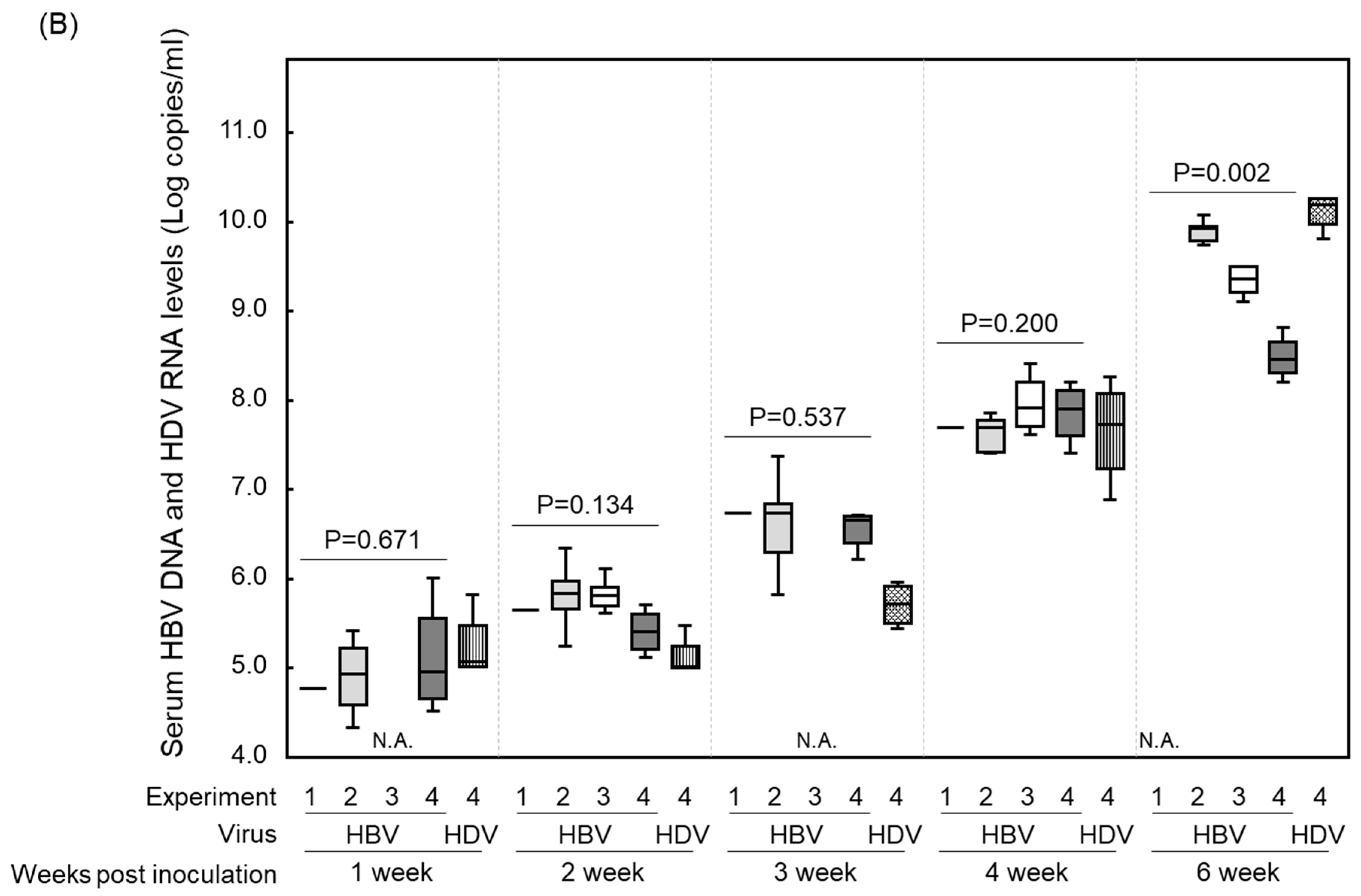
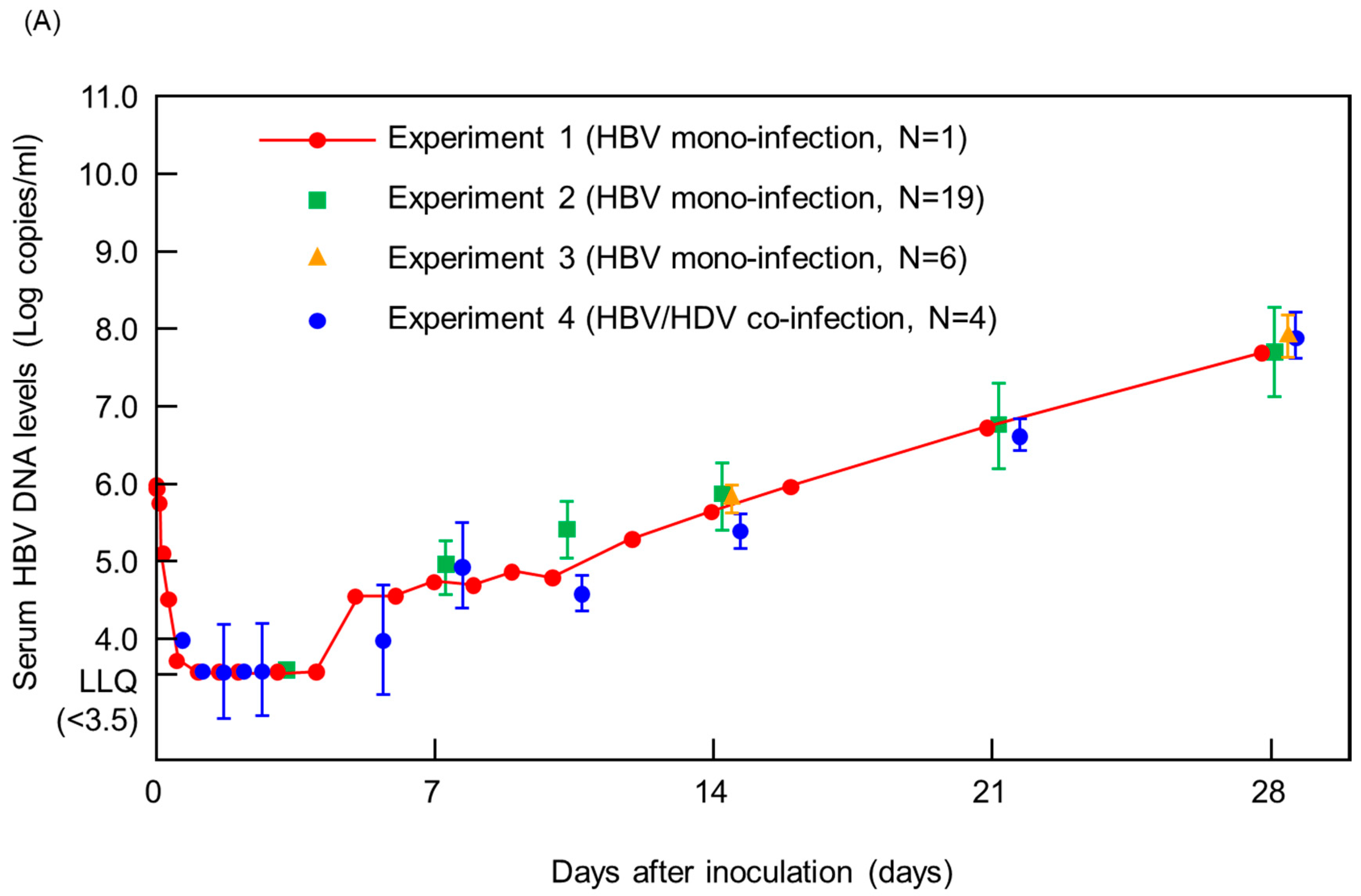
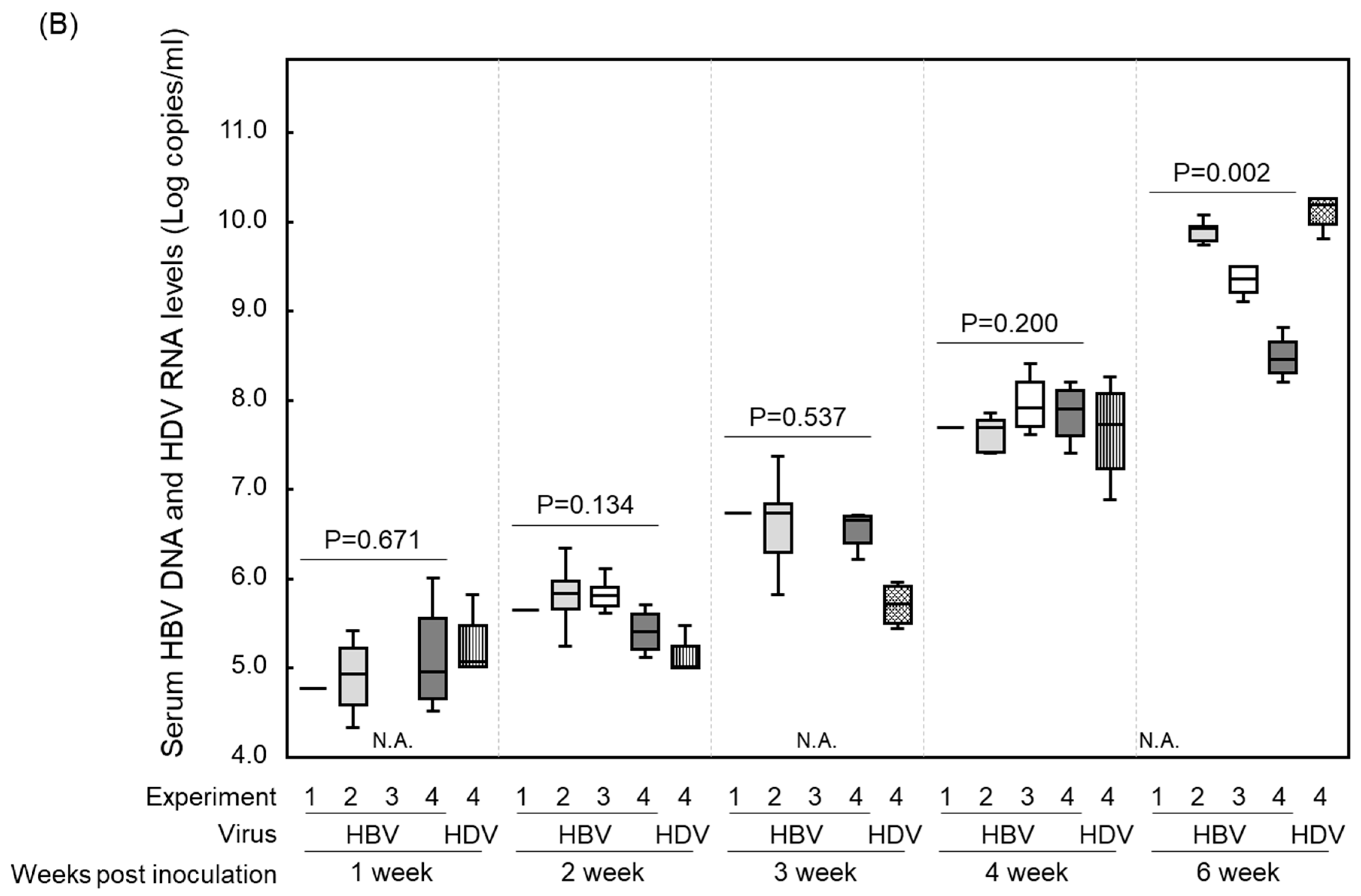
3. Results and Discussions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HBV | hepatitis B virus |
| HDV | hepatitis D virus |
References
- Ishida, Y.; Chung, T.L.; Imamura, M.; Hiraga, N.; Sen, S.; Yokomichi, H.; Tateno, C.; Canini, L.; Perelson, A.S.; Uprichard, S.L.; et al. Acute hepatitis B virus infection in humanized chimeric mice has multiphasic viral kinetics. Hepatology 2018, 68, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Sureau, C.; Negro, F. The hepatitis delta virus: Replication and pathogenesis. J. Hepatol. 2016, 64, S102–S116. [Google Scholar] [CrossRef] [PubMed]
- Giersch, K.; Allweiss, L.; Volz, T.; Helbig, M.; Bierwolf, J.; Lohse, A.W.; Pollok, J.M.; Petersen, J.; Dandri, M.; Lutgehetmann, M. Hepatitis Delta co-infection in humanized mice leads to pronounced induction of innate immune responses in comparison to HBV mono-infection. J. Hepatol. 2015, 63, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Tateno, C.; Kawase, Y.; Tobita, Y.; Hamamura, S.; Ohshita, H.; Yokomichi, H.; Sanada, H.; Kakuni, M.; Shiota, A.; Kojima, Y.; et al. Generation of Novel Chimeric Mice with Humanized Livers by Using Hemizygous cDNA-uPA/SCID Mice. PLoS ONE 2015, 10, e0142145. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, M.; Hiraga, N.; Zhang, Y.; Yamashita, M.; Sato, O.; Oka, N.; Shiraishi, K.; Izaki, Y.; Makokha, G.N.; Uchida, T.; et al. Endoplasmic reticulum-mediated induction of interleukin-8 occurs by hepatitis B virus infection and contributes to suppression of interferon responsiveness in human hepatocytes. Virology 2018, 525, 48–61. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuge, M.; Uchida, T.; Walsh, K.; Ishida, Y.; Tateno, C.; Kumar, U.; Glenn, J.S.; Koh, C.; Heller, T.; Uprichard, S.L.; et al. Early Multiphasic HBV Infection Initiation Kinetics Is Not Clone-Specific and Is Not Affected by Hepatitis D Virus (HDV) Infection. Viruses 2019, 11, 263. https://doi.org/10.3390/v11030263
Tsuge M, Uchida T, Walsh K, Ishida Y, Tateno C, Kumar U, Glenn JS, Koh C, Heller T, Uprichard SL, et al. Early Multiphasic HBV Infection Initiation Kinetics Is Not Clone-Specific and Is Not Affected by Hepatitis D Virus (HDV) Infection. Viruses. 2019; 11(3):263. https://doi.org/10.3390/v11030263
Chicago/Turabian StyleTsuge, Masataka, Takuro Uchida, Kevin Walsh, Yuji Ishida, Chise Tateno, Upendra Kumar, Jeffrey S. Glenn, Christopher Koh, Theo Heller, Susan L. Uprichard, and et al. 2019. "Early Multiphasic HBV Infection Initiation Kinetics Is Not Clone-Specific and Is Not Affected by Hepatitis D Virus (HDV) Infection" Viruses 11, no. 3: 263. https://doi.org/10.3390/v11030263
APA StyleTsuge, M., Uchida, T., Walsh, K., Ishida, Y., Tateno, C., Kumar, U., Glenn, J. S., Koh, C., Heller, T., Uprichard, S. L., Dahari, H., & Chayama, K. (2019). Early Multiphasic HBV Infection Initiation Kinetics Is Not Clone-Specific and Is Not Affected by Hepatitis D Virus (HDV) Infection. Viruses, 11(3), 263. https://doi.org/10.3390/v11030263