Monophosphoryl-Lipid A (MPLA) is an Efficacious Adjuvant for Inactivated Rabies Vaccines



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Antibodies
2.3. Animals
2.4. Virus Titration
2.5. Inactivation of RABV
2.6. Preparation of Bone Marrow-Derived DCs (BMDCs)
2.7. Titration of RABV-Specific VNA
2.8. Measurement of RABV-Specific Total-IgG and Immunoglobulin Isotyping
2.9. Flow Cytometric Analysis
2.10. Statistics
2.11. Ethical Approval
3. Results
3.1. MPLA Facilitates the Maturation of BMDC Via a TLR4-Dependent Pathway
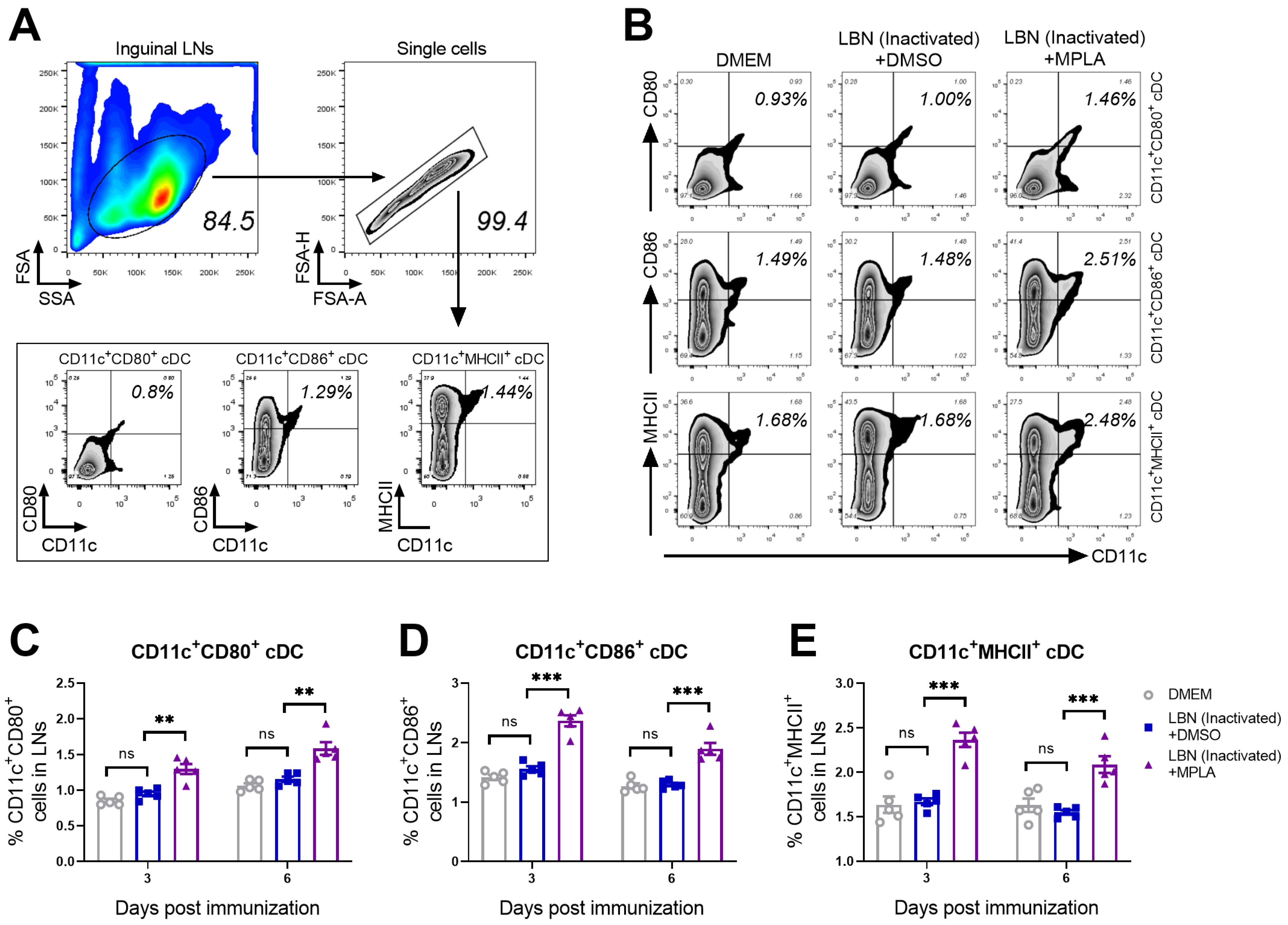
3.2. MPLA Promotes the Maturation of cDCs in LNs Post RABV Immunization
3.3. MPLA Enhances the Recruitment of Tfh Cells in LNs Post RABV Immunization
3.4. MPLA Augments the Proliferation of GC B Cells in LNs Post RABV Immunization
3.5. MPLA Facilitates the Generation of Plasma Cells (PCs) in BMs Post RABV Immunization
3.6. MPLA Improves Antibody Production and Protection Against Virulent RABV Challenge
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Galvez-Romero, G.; Salas-Rojas, M.; Pompa-Mera, E.N.; Chavez-Rueda, K.; Aguilar-Setien, A. Addition of C3d-P28 adjuvant to a rabies DNA vaccine encoding the G5 linear epitope enhances the humoral immune response and confers protection. Vaccine 2018, 36, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Bonnaud, E.M.; Troupin, C.; Dacheux, L.; Holmes, E.C.; Monchatre-Leroy, E.; Tanguy, M.; Bouchier, C.; Cliquet, F.; Barrat, J.; Bourhy, H. Comparison of intra- and inter-host genetic diversity in rabies virus during experimental cross-species transmission. PLoS Pathog. 2019, 15, e1007799. [Google Scholar] [CrossRef] [PubMed]
- Kip, E.; Staal, J.; Tima, H.G.; Verstrepen, L.; Romano, M.; Lemeire, K.; Suin, V.; Hamouda, A.; Baens, M.; Libert, C.; et al. Inhibition of MALT1 Decreases Neuroinflammation and Pathogenicity of Virulent Rabies Virus in Mice. J. Virol. 2018, 92, e00720. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.S.; Selden, D.; Wu, G.; Wright, E.; Horton, D.L.; Fooks, A.R.; Fooks, A.R.; Banyard, A.C. Antigenic site changes in the rabies virus glycoprotein dictates functionality and neutralizing capability against divergent lyssaviruses. J. Gen. Virol. 2018, 99, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, Y.; Huang, Y.; Gnanadurai, C.W.; Zhou, M.; Zhao, L.; Fu, Z.F. The ectodomain of rabies virus glycoprotein determines dendritic cell activation. Antivir. Res. 2017, 141, 1–6. [Google Scholar] [CrossRef]
- Anilionis, A.; Wunner, W.H.; Curtis, P.J. Structure of the glycoprotein gene in rabies virus. Nature 1981, 294, 275–278. [Google Scholar] [CrossRef]
- Lentz, T.L.; Burrage, T.G.; Smith, A.L.; Crick, J.; Tignor, G.H. Is the Acetylcholine-Receptor a Rabies Virus Receptor. Science 1982, 215, 182–184. [Google Scholar] [CrossRef]
- Thoulouze, M.I.; Lafage, M.; Schachner, M.; Hartmann, U.; Cremer, H.; Lafon, M. The neural cell adhesion molecule is a receptor for rabies virus. J. Virol. 1998, 72, 7181–7190. [Google Scholar]
- Wiktor, T.J.; Macfarlan, R.I.; Reagan, K.J.; Dietzschold, B.; Curtis, P.J.; Wunner, W.H.; Kieny, M.P.; Lathe, R.; Lecocq, J.P.; Mackett, M.; et al. Protection from rabies by a vaccinia virus recombinant containing the rabies virus glycoprotein gene. Proc. Natl. Acad. Sci. USA 1984, 81, 7194–7198. [Google Scholar] [CrossRef]
- Wang, X.; Feng, N.; Ge, J.; Shuai, L.; Peng, L.; Gao, Y.; Yang, S.; Xia, X.; Bu, Z. Recombinant canine distemper virus serves as bivalent live vaccine against rabies and canine distemper. Vaccine 2012, 30, 5067–5072. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y.; Fooks, A.R.; Zhang, F.; Hu, R. Oral vaccination of dogs (Canis familiaris) with baits containing the recombinant rabies-canine adenovirus type-2 vaccine confers long-lasting immunity against rabies. Vaccine 2008, 26, 345–350. [Google Scholar] [CrossRef] [PubMed]
- DiStefano, D.; Antonello, J.M.; Bett, A.J.; Medi, M.B.; Casimiro, D.R.; ter Meulen, J. Immunogenicity of a reduced-dose whole killed rabies vaccine is significantly enhanced by ISCOMATRIX adjuvant, Merck amorphous aluminum hydroxylphosphate sulfate (MAA) or a synthetic TLR9 agonist in rhesus macaques. Vaccine 2013, 31, 4888–4893. [Google Scholar] [CrossRef] [PubMed]
- Kalimuddin, S.; Wijaya, L.; Chan, Y.F.Z.; Wong, A.W.L.; Oh, H.M.L.; Wang, L.F.; Kassim, J.A.; Zhao, J.; Shi, Z.; Low, J.G. A phase II randomized study to determine the safety and immunogenicity of the novel PIKA rabies vaccine containing the PIKA adjuvant using an accelerated regimen. Vaccine 2017, 35, 7127–7132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.F.; Li, W.; Hu, Y.C.; Zhao, J.Y.; Liu, F.; Lin, H.; Liu, Y.; Wang, L.; Xu, S.; et al. A novel rabies vaccine based-on toll-like receptor 3 (TLR3) agonist PIKA adjuvant exhibiting excellent safety and efficacy in animal studies. Virology 2016, 489, 165–172. [Google Scholar] [CrossRef]
- Barr, T.A.; Brown, S.; Ryan, G.; Zhao, J.; Gray, D. TLR-mediated stimulation of APC: Distinct cytokine responses of B cells and dendritic cells. Eur. J. Immunol. 2007, 37, 3040–3053. [Google Scholar] [CrossRef]
- Gupta, A.; Cooper, Z.A.; Tulapurkar, M.E.; Potla, R.; Maity, T.; Hasday, J.D.; Singh, I.S. Toll-like receptor agonists and febrile range hyperthermia synergize to induce heat shock protein 70 expression and extracellular release. J. Biol. Chem. 2013, 288, 2756–2766. [Google Scholar] [CrossRef]
- Goff, P.H.; Hayashi, T.; Martinez-Gil, L.; Corr, M.; Crain, B.; Yao, S.; Cottam, H.B.; Chan, M.; Ramos, I.; Eggink, D.; et al. Synthetic Toll-like receptor 4 (TLR4) and TLR7 ligands as influenza virus vaccine adjuvants induce rapid, sustained, and broadly protective responses. J. Virol. 2015, 89, 3221–3235. [Google Scholar] [CrossRef]
- Boukhvalova, M.S.; Prince, G.A.; Soroush, L.; Harrigan, D.C.; Vogel, S.N.; Blanco, J.C. The TLR4 agonist, monophosphoryl lipid A, attenuates the cytokine storm associated with respiratory syncytial virus vaccine-enhanced disease. Vaccine 2006, 24, 5027–5035. [Google Scholar] [CrossRef]
- Wang, Z.; Liang, Q.; Zhang, Y.J.; Yang, J.; Li, M.M.; Wang, K.L.; Cui, M.; Chen, H.; Fu, Z.F.; Zhao, L.; et al. An optimized HMGB1 expressed by recombinant rabies virus enhances immunogenicity through activation of dendritic cells in mice. Oncotarget 2017, 8, 83539–83554. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, P.A.; Maggi, J.; Schinnerling, K.; Sepulveda-Gutierrez, A.; Soto, L.; Neira, O.; Mehdi, A.M.; Nel, H.J.; Pesce, B.; Aravena, O.; et al. Regulation of Tolerogenic Features on Dexamethasone-Modulated MPLA-Activated Dendritic Cells by MYC. Front. Immunol. 2019, 10, 1171. [Google Scholar] [CrossRef]
- Patil, H.P.; Murugappan, S.; ter Veer, W.; Meijerhof, T.; de Haan, A.; Frijlink, H.W.; Wilschut, J.; Hinrichs, W.L.; Huckriede, A. Evaluation of monophosphoryl lipid A as adjuvant for pulmonary delivered influenza vaccine. J. Control. Release 2014, 174, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Wang, H.; Wu, H.; Yang, F.; Tripp, R.A.; Hogan, R.J.; Fu, Z.F. Rabies virus expressing dendritic cell-activating molecules enhances the innate and adaptive immune response to vaccination. J. Virol. 2011, 85, 1634–1644. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, M.; Luo, Z.; Zhang, Y.; Cui, M.; Chen, H.; Fu, Z.F.; Zhao, L. Overexpression of Interleukin-7 Extends the Humoral Immune Response Induced by Rabies Vaccination. J. Virol. 2017, 91, e02324. [Google Scholar] [CrossRef]
- Yu, F.L.; Zhang, G.Q.; Zhong, X.F.; Han, N.; Song, Y.F.; Zhao, L.; Cui, M.; Rayner, S.; Fu, Z.F. Comparison of complete genome sequences of dog rabies viruses isolated from China and Mexico reveals key amino acid changes that may be associated with virus replication and virulence. Arch. Virol. 2014, 159, 1593–1601. [Google Scholar] [CrossRef]
- Dietzschold, B.; Morimoto, K.; Hooper, D.C.; Smith, J.S.; Rupprecht, C.E.; Koprowski, H. Genotypic and phenotypic diversity of rabies virus variants involved in human rabies: Implications for postexposure prophylaxis. J. Hum. Virol. 2000, 3, 50–57. [Google Scholar] [PubMed]
- Wang, Z.; Li, M.; Zhou, M.; Zhang, Y.; Yang, J.; Cao, Y.; Wang, K.; Cui, M.; Chen, H.; Fu, Z.F.; et al. A Novel Rabies Vaccine Expressing CXCL13 Enhances Humoral Immunity by Recruiting both T Follicular Helper and Germinal Center B Cells. J. Virol. 2017, 91, e01956. [Google Scholar] [CrossRef]
- Tang, H.B.; Lu, Z.L.; Wei, X.K.; Zhong, T.Z.; Zhong, Y.Z.; Ouyang, L.X.; Luo, Y.; Xing, X.W.; Liao, F.; Peng, K.K.; et al. Viperin inhibits rabies virus replication via reduced cholesterol and sphingomyelin and is regulated upstream by TLR4. Sci. Rep. 2016, 6, 30529. [Google Scholar] [CrossRef]
- Greco, F.; Toinon, A.; Moreno, N.; Nicolai, M.C.; Manin, C.; Guinet-Morlot, F.; Ronzon, F. Study of rabies virus by Differential Scanning Calorimetry: Identification of Proteins Involved in Thermal Transitions. Protein Sci. 2015, 24, 97. [Google Scholar]
- Morgeaux, S.; Tordo, N.; Gontier, C.; Perrin, P. Beta-propiolactone treatment impairs the biological activity of residual DNA from BHK-21 cells infected with rabies virus. Vaccine 1993, 11, 82–90. [Google Scholar] [CrossRef]
- Faul, E.J.; Wanjalla, C.N.; Suthar, M.S.; Gale, M.; Wirblich, C.; Schnell, M.J. Rabies virus infection induces type I interferon production in an IPS-1 dependent manner while dendritic cell activation relies on IFNAR signaling. PLoS Pathog. 2010, 6, e1001016. [Google Scholar] [CrossRef]
- Zhao, L.; Toriumi, H.; Wang, H.; Kuang, Y.; Guo, X.; Morimoto, K.; Fu, Z.F. Expression of MIP-1alpha (CCL3) by a recombinant rabies virus enhances its immunogenicity by inducing innate immunity and recruiting dendritic cells and B cells. J. Virol. 2010, 84, 9642–9648. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Perez, F.; Goretzki, A.; Wolfheimer, S.; Schulke, S. The vaccine adjuvant MPLA activates glycolytic metabolism in mouse mDC by a JNK-dependent activation of mTOR-signaling. Mol. Immunol. 2019, 106, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, L.; Zhou, S.; Wang, Z.; Ruan, J.; Tang, L.; Jia, Z.; Cui, M.; Zhao, L.; Fu, Z.F. Recombinant rabies virus expressing dog GM-CSF is an efficacious oral rabies vaccine for dogs. Oncotarget 2015, 6, 38504–38516. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.; Han, J.A.; Koh, H.; Choi, B.; Cho, Y.; Jeong, H.; Ra, J.S.; Sung, P.S.; Shin, E.C.; Ryu, S.; et al. CD8alpha(-) Dendritic Cells Induce Antigen-Specific T Follicular Helper Cells Generating Efficient Humoral Immune Responses. Cell Rep. 2015, 11, 1929–1940. [Google Scholar] [CrossRef]
- Mildner, A.; Jung, S. Development and function of dendritic cell subsets. Immunity 2014, 40, 642–656. [Google Scholar] [CrossRef]
- Kawamura, K.; Iyonaga, K.; Ichiyasu, H.; Nagano, J.; Suga, M.; Sasaki, Y. Differentiation, maturation, and survival of dendritic cells by osteopontin regulation. Clin. Diagn. Lab. Immunol. 2005, 12, 206–212. [Google Scholar] [CrossRef]
- Fang, H.; Wu, Y.; Huang, X.; Wang, W.; Ang, B.; Cao, X.; Wan, T. Toll-like receptor 4 (TLR4) is essential for Hsp70-like protein 1 (HSP70L1) to activate dendritic cells and induce Th1 response. J. Biol. Chem. 2011, 286, 30393–30400. [Google Scholar] [CrossRef]
- Haberman, A.M.; Gonzalez, D.G.; Wong, P.; Zhang, T.T.; Kerfoot, S.M. Germinal center B cell initiation, GC maturation, and the coevolution of its stromal cell niches. Immunol. Rev. 2019, 288, 10–27. [Google Scholar] [CrossRef]
- Hamel, K.M.; Liarski, V.M.; Clark, M.R. Germinal center B-cells. Autoimmunity 2012, 45, 333–347. [Google Scholar] [CrossRef]
- Smith, K.M.; Pottage, L.; Thomas, E.R.; Leishman, A.J.; Doig, T.N.; Xu, D.; Liew, F.Y.; Garside, P. Th1 and Th2 CD4+ T Cells Provide Help for B Cell Clonal Expansion and Antibody Synthesis in a Similar Manner In Vivo. J. Immunol. 2000, 165, 3136–3144. [Google Scholar] [CrossRef]
- Hwang, I.Y.; Park, C.; Harrison, K.; Kehrl, J.H. TLR4 signaling augments B lymphocyte migration and overcomes the restriction that limits access to germinal center dark zones. J. Exp. Med. 2009, 206, 2641–2657. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, J.; Yang, Y.; Khan, I.; Zhu, N. Rabies virus lipopeptide conjugated to a TLR7 agonist improves the magnitude and quality of the Th1-biased humoral immune response in mice. Virology 2016, 497, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.F.; Kurup, D.; Hagen, K.R.; Fisher, C.; Keshwara, R.; Papaneri, A.; Perry, D.L.; Cooper, K.; Jahrling, P.B.; Wang, J.T.; et al. An Inactivated Rabies Virus-Based Ebola Vaccine, FILORAB1, Adjuvanted With Glucopyranosyl Lipid A in Stable Emulsion Confers Complete Protection in Nonhuman Primate Challenge Models. J. Infect. Dis. 2016, 214 (Suppl. 3), S342–S354. [Google Scholar] [CrossRef]
- Rajaiah, R.; Perkins, D.J.; Ireland, D.D.; Vogel, S.N. CD14 dependence of TLR4 endocytosis and TRIF signaling displays ligand specificity and is dissociable in endotoxin tolerance. Proc. Natl. Acad. Sci. USA 2015, 112, 8391–8396. [Google Scholar] [CrossRef]
- Pouliot, K.; Buglione-Corbett, R.; Marty-Roix, R.; Montminy-Paquette, S.; West, K.; Wang, S.; Lu, S.; Lien, E. Contribution of TLR4 and MyD88 for adjuvant monophosphoryl lipid A (MPLA) activity in a DNA prime-protein boost HIV-1 vaccine. Vaccine 2014, 32, 5049–5056. [Google Scholar] [CrossRef]
- Hu, X.; Liu, R.; Zhu, N. Enhancement of humoral and cellular immune responses by monophosphoryl lipid A (MPLA) as an adjuvant to the rabies vaccine in BALB/c mice. Immunobiology 2013, 218, 1524–1528. [Google Scholar] [CrossRef]
- Shin, T.; Kennedy, G.; Gorski, K.; Tsuchiya, H.; Koseki, H.; Azuma, M.; Yagita, H.; Chen, L.; Powell, J.; Pardoll, D.; et al. Cooperative B7-1/2 (CD80/CD86) and B7-DC costimulation of CD4+ T cells independent of the PD-1 receptor. J. Exp. Med. 2003, 198, 31–38. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, Z.; Wang, C. Persistence of rabies antibody 5 years after postexposure prophylaxis with vero cell antirabies vaccine and antibody response to a single booster dose. Clin. Vaccine Immunol. CVI 2011, 18, 1477–1479. [Google Scholar] [CrossRef]
- Anderson, D.C. WHO guidelines dealing with immunoglobulin use impede rabies prevention. Asian Biomed 2007, 1, 103–107. [Google Scholar]
- Yanagisawa, N.; Takayama, N.; Nakayama, E.; Mannen, K.; Suganuma, A. Pre-exposure immunization against rabies using Japanese rabies vaccine following the WHO recommended schedule. J. Infect. Chemother. 2010, 16, 38–41. [Google Scholar] [CrossRef]
- Lebrun, A.; Garcia, S.; Li, J.; Kean, R.B.; Hooper, D.C. Protection Against CNS-Targeted Rabies Virus Infection is Dependent upon Type-1 Immune Mechanisms Induced by Live-Attenuated Rabies Vaccines. Trop. Med. Infect. Dis. 2017, 2, 22. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Zhang, C.; Li, R.; Wang, Z.; Yuan, Y.; Li, H.; Fu, Z.; Zhou, M.; Zhao, L. Monophosphoryl-Lipid A (MPLA) is an Efficacious Adjuvant for Inactivated Rabies Vaccines. Viruses 2019, 11, 1118. https://doi.org/10.3390/v11121118
Chen C, Zhang C, Li R, Wang Z, Yuan Y, Li H, Fu Z, Zhou M, Zhao L. Monophosphoryl-Lipid A (MPLA) is an Efficacious Adjuvant for Inactivated Rabies Vaccines. Viruses. 2019; 11(12):1118. https://doi.org/10.3390/v11121118
Chicago/Turabian StyleChen, Chen, Chengguang Zhang, Ruiming Li, Zongmei Wang, Yueming Yuan, Haoqi Li, Zhenfang Fu, Ming Zhou, and Ling Zhao. 2019. "Monophosphoryl-Lipid A (MPLA) is an Efficacious Adjuvant for Inactivated Rabies Vaccines" Viruses 11, no. 12: 1118. https://doi.org/10.3390/v11121118
APA StyleChen, C., Zhang, C., Li, R., Wang, Z., Yuan, Y., Li, H., Fu, Z., Zhou, M., & Zhao, L. (2019). Monophosphoryl-Lipid A (MPLA) is an Efficacious Adjuvant for Inactivated Rabies Vaccines. Viruses, 11(12), 1118. https://doi.org/10.3390/v11121118