Porcine Circovirus Type 2 Induces Single Immunoglobulin Interleukin-1 Related Receptor (SIGIRR) Downregulation to Promote Interleukin-1β Upregulation in Porcine Alveolar Macrophage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus Preparation
2.2. Infection, RNA Extraction, and cDNA Synthesis
2.3. The Real-Time Quantitative PCR Assay
2.4. Analysis of Cytokine Expression in Protein Level
2.5. Western Blot Analysis
2.6. Inhibition of Signal Transduction Pathway
2.7. SIGIRR Overexpression and Knocked off in 3D4/2 Cells
2.8. Immunofluorescence Assay (IFA)
2.9. ELISA
2.10. Statistic Analysis
2.11. Ethics Statement
3. Results
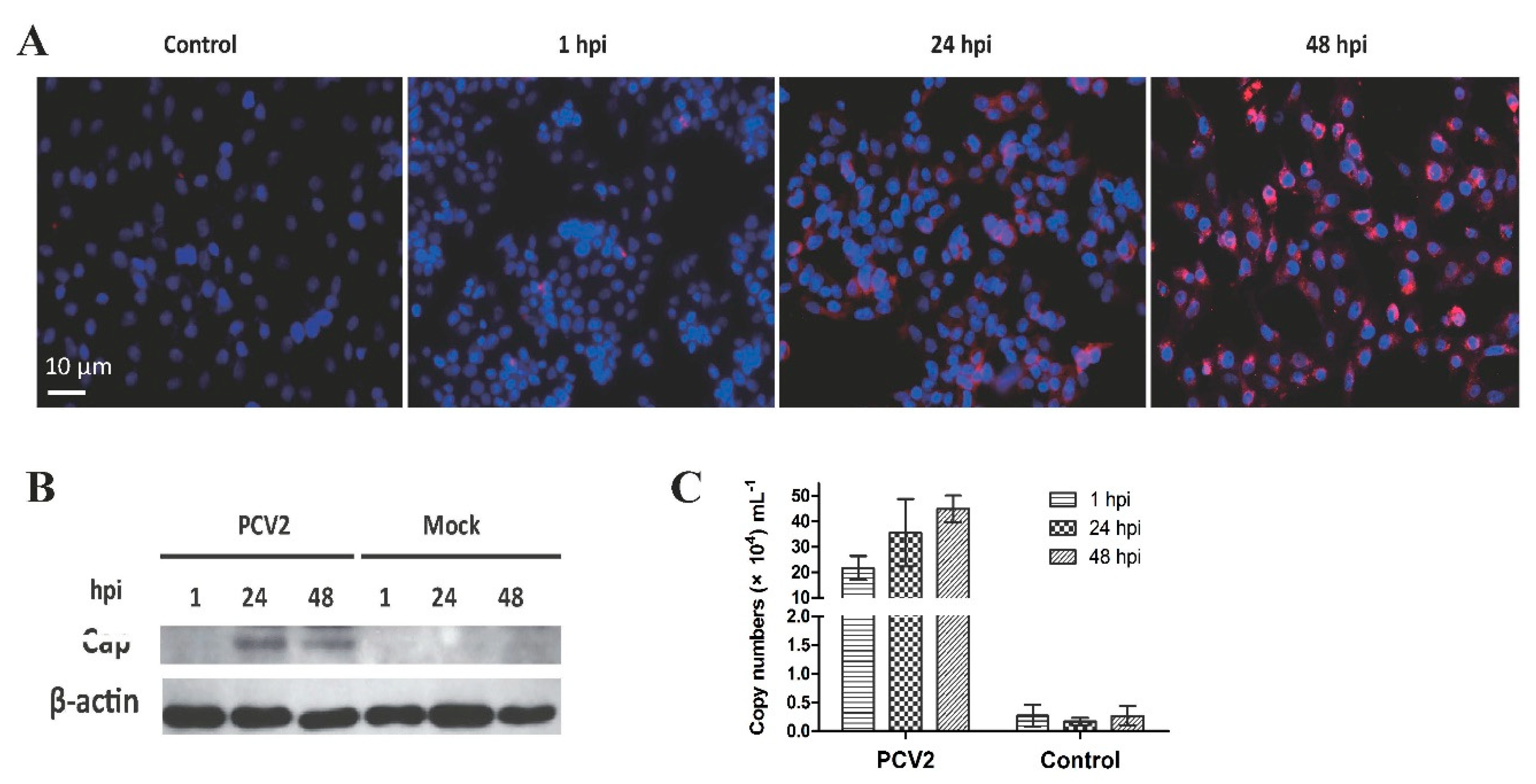
3.1. Confirmation of PCV2b Infection in PAMs
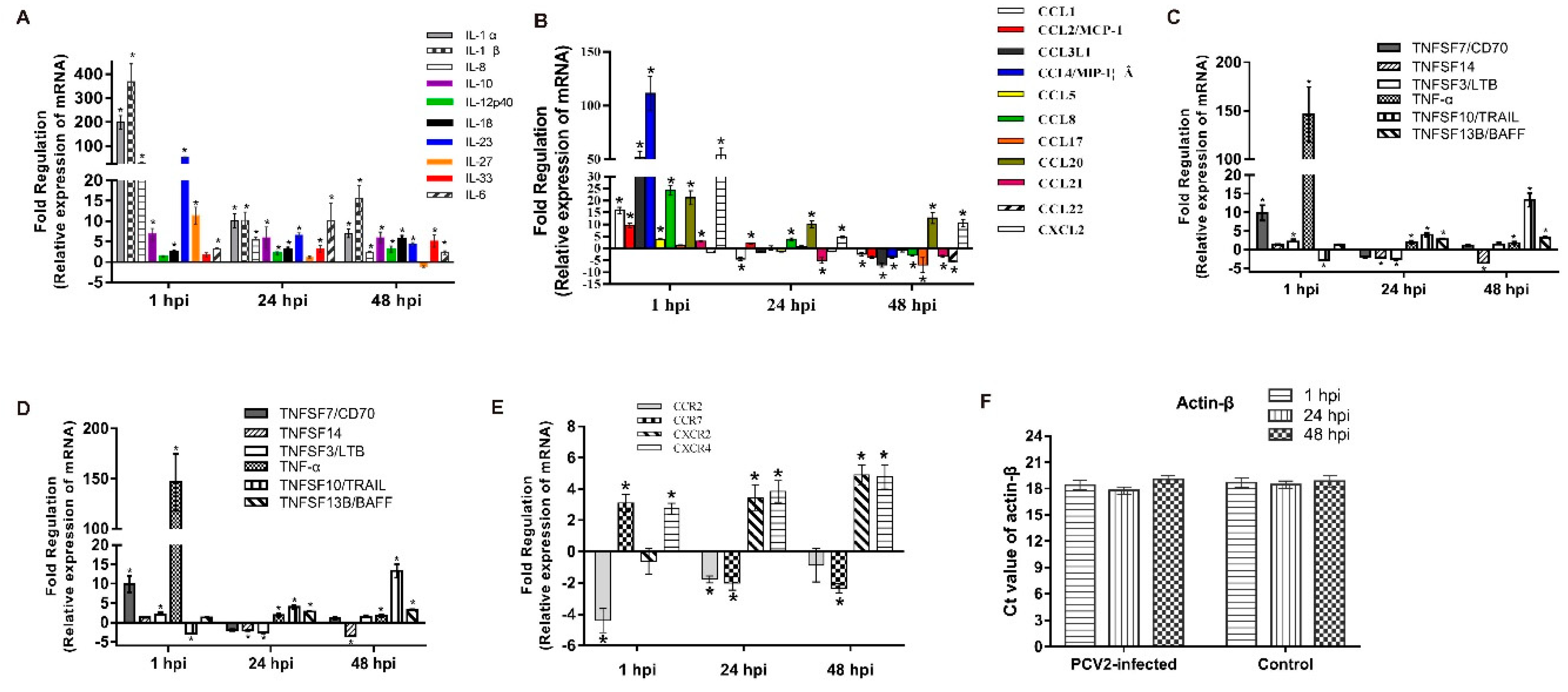
3.2. mRNA Expression Profiles
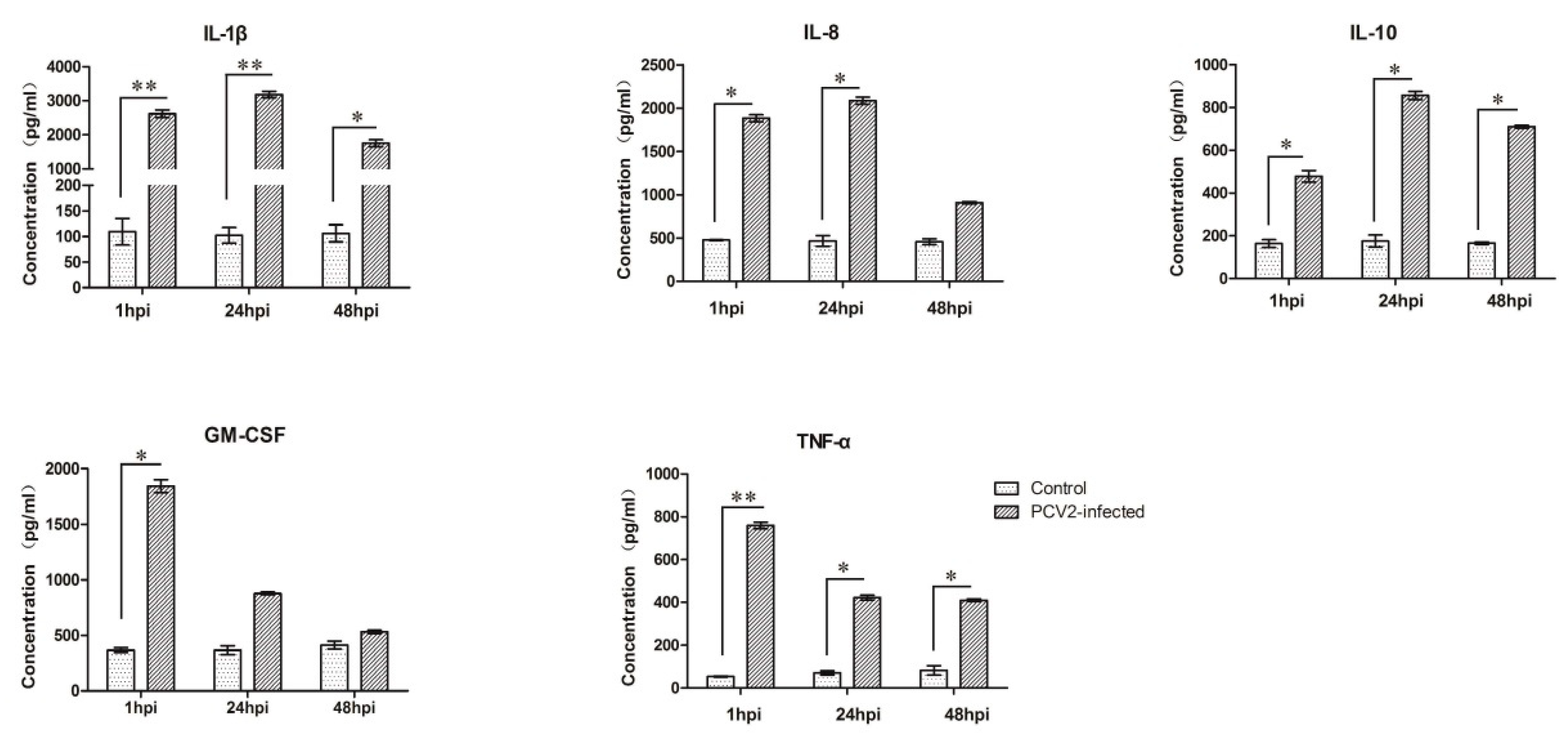
3.3. Expression Analyses of Several Cytokines in PCV2b-Infected PAMs
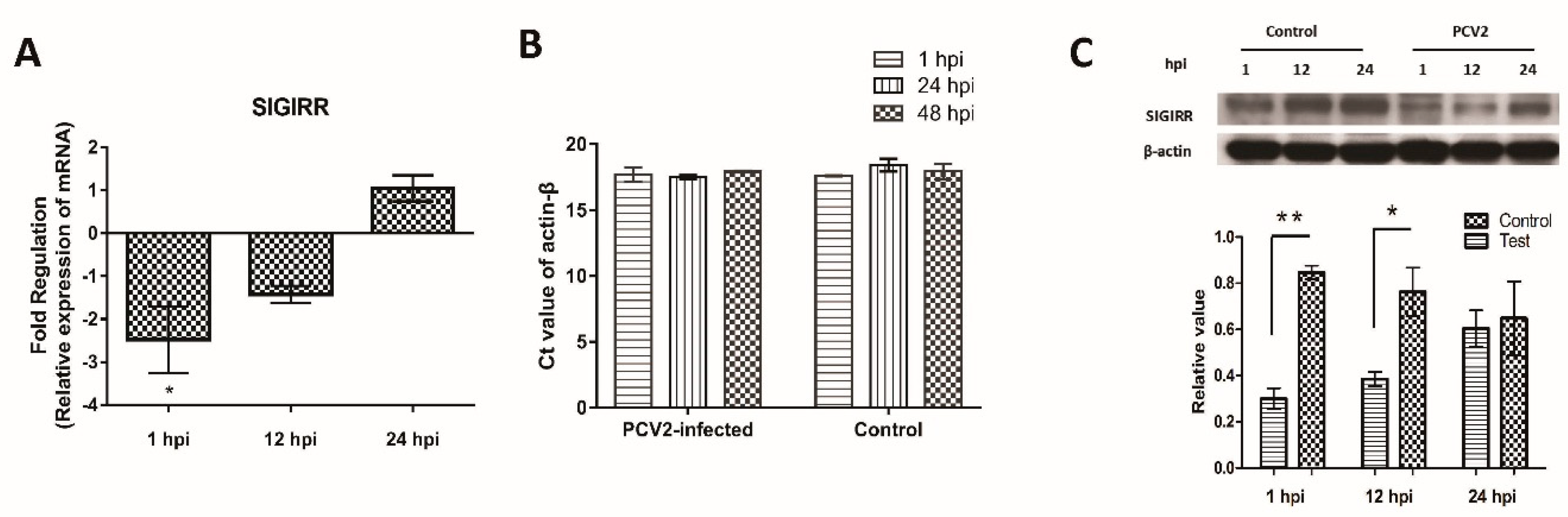
3.4. Downregulation of SIGIRR Expression in PCV2b-Infected PAMs
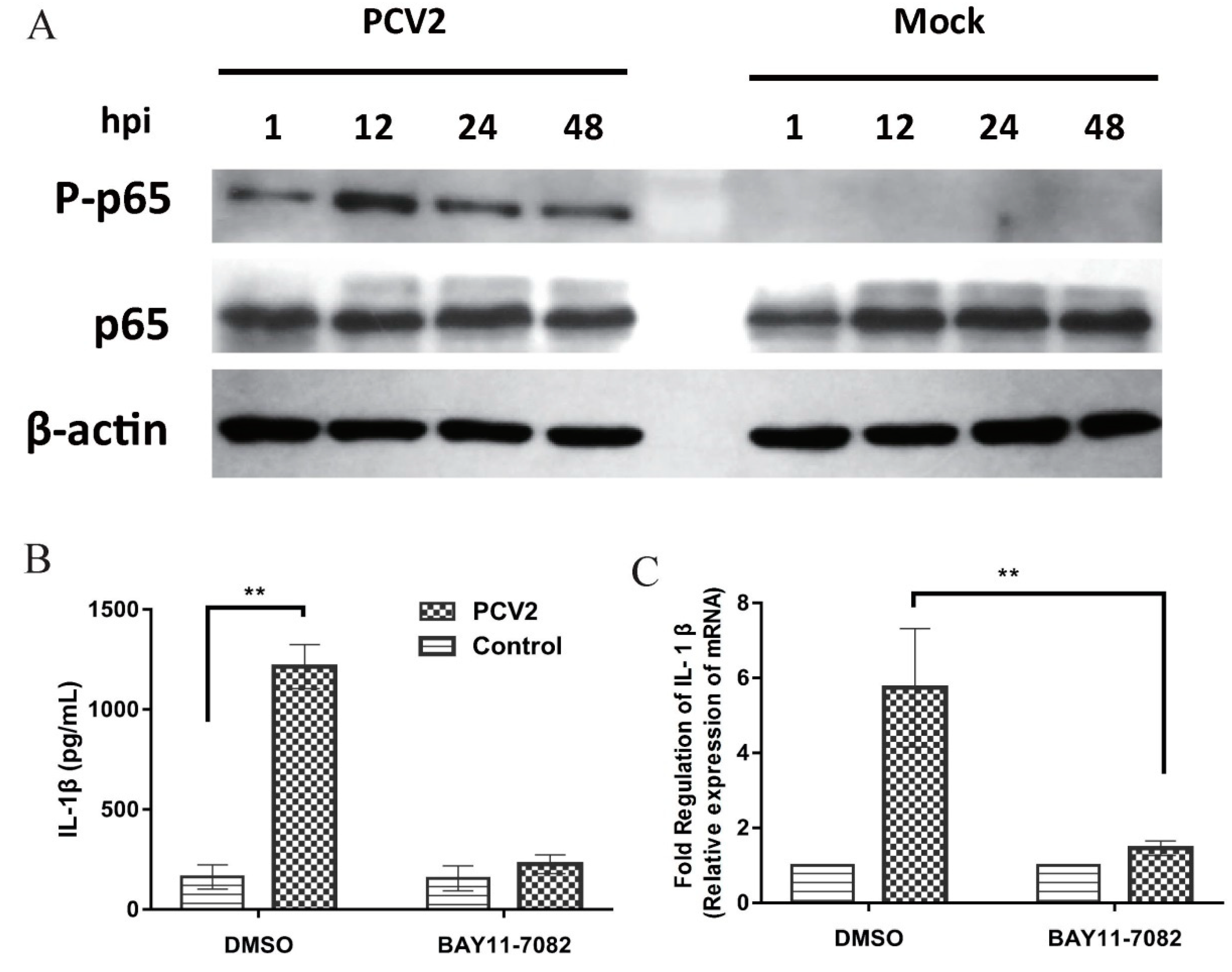
3.5. NF-κB Activation Is Involved in PCV2b-Induced IL-1β Secretion
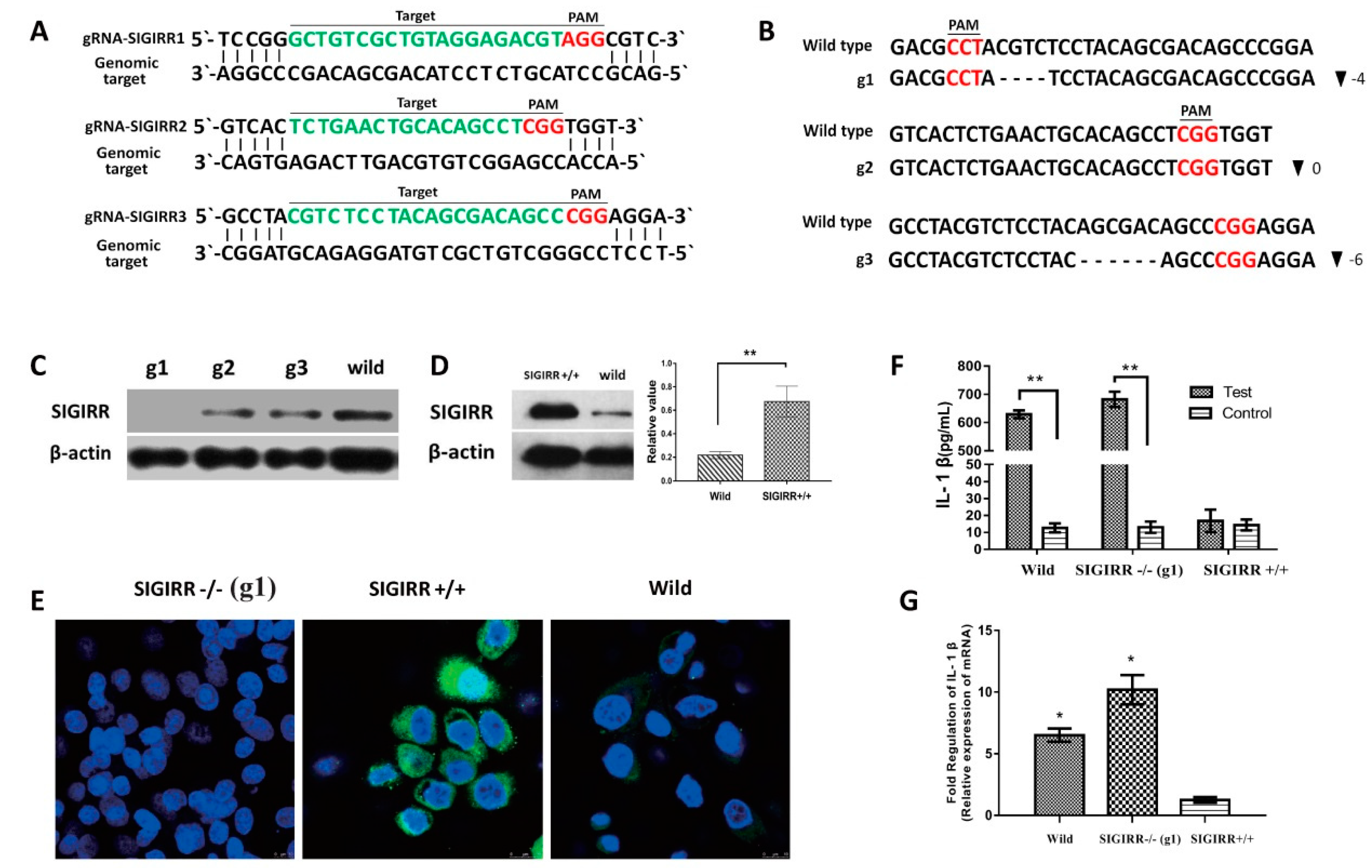
3.6. SIGIRR Is Associated with PCV2b-Induced IL-1β Secretion
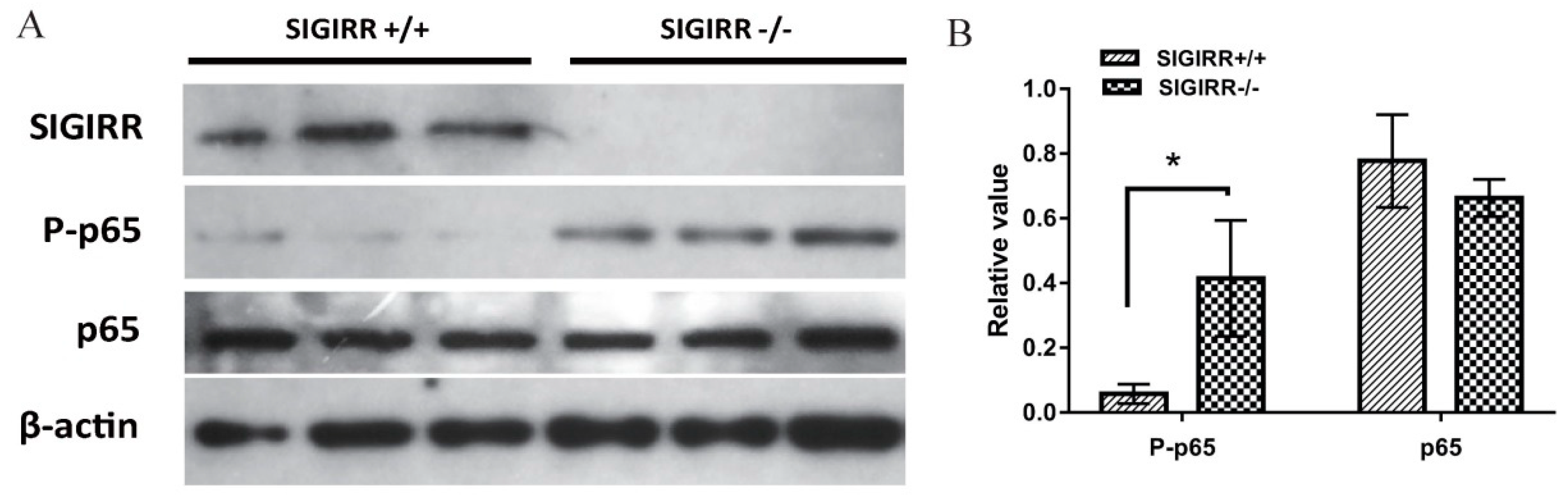
3.7. SIGIRR Is Involved in PCV2-Induced NF-κB Activation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Finsterbusch, T.; Mankertz, A. Porcine circoviruses-small but powerful. Virus Res. 2009, 143, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Segales, J.; Kekarainen, T.; Cortey, M. The natural history of porcine circovirus type 2: From an inoffensive virus to a devastating swine disease? Vet. Microbiol. 2013, 165, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Wang, F.; Guo, X.; Yang, H. Porcine circovirus type 2 and its associated diseases in china. Virus Res. 2012, 164, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J. Spread like a wildfire--the omnipresence of porcine circovirus type 2 (pcv2) and its ever-expanding association with diseases in pigs. Virus Res. 2012, 164, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Darwich, L.; Mateu, E. Immunology of porcine circovirus type 2 (pcv2). Virus Res. 2012, 164, 61–67. [Google Scholar] [CrossRef]
- Segales, J. Porcine circovirus type 2 (pcv2) infections: Clinical signs, pathology and laboratory diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef]
- Darwich, L.; Pie, S.; Rovira, A.; Segales, J.; Domingo, M.; Oswald, I.P.; Mateu, E. Cytokine mrna expression profiles in lymphoid tissues of pigs naturally affected by postweaning multisystemic wasting syndrome. J. Gen. Virol. 2003, 84, 2117–2125. [Google Scholar] [CrossRef]
- Vincent, I.E.; Carrasco, C.P.; Herrmann, B.; Meehan, B.M.; Allan, G.M.; Summerfield, A.; McCullough, K.C. Dendritic cells harbor infectious porcine circovirus type 2 in the absence of apparent cell modulation or replication of the virus. J. Virol. 2003, 77, 13288–13300. [Google Scholar] [CrossRef]
- Yu, S.; Opriessnig, T.; Kitikoon, P.; Nilubol, D.; Halbur, P.G.; Thacker, E. Porcine circovirus type 2 (pcv2) distribution and replication in tissues and immune cells in early infected pigs. Vet. Immunol. Immunopathol. 2007, 115, 261–272. [Google Scholar] [CrossRef]
- Gilpin, D.F.; McCullough, K.; Meehan, B.M.; McNeilly, F.; McNair, I.; Stevenson, L.S.; Foster, J.C.; Ellis, J.A.; Krakowka, S.; Adair, B.M.; et al. In vitro studies on the infection and replication of porcine circovirus type 2 in cells of the porcine immune system. Vet. Immunol. Immunopathol. 2003, 94, 149–161. [Google Scholar] [CrossRef]
- Basta, S.; Carrasco, C.P.; Knoetig, S.M.; Rigden, R.C.; Gerber, H.; Summerfield, A.; McCullough, K.C. Porcine alveolar macrophages: Poor accessory or effective suppressor cells for t-lymphocytes. Vet. Immunol. Immunopathol. 2000, 77, 177–190. [Google Scholar] [CrossRef]
- Chae, J.S.; Choi, K.S. Proinflammatory cytokine expression in the lung of pigs with porcine circovirus type 2-associated respiratory disease. Res. Vet. Sci 2011, 90, 321–323. [Google Scholar] [CrossRef] [PubMed]
- Riva, F.; Bonavita, E.; Barbati, E.; Muzio, M.; Mantovani, A.; Garlanda, C. Tir8/sigirr is an interleukin-1 receptor/toll like receptor family member with regulatory functions in inflammation and immunity. Front. Immunol. 2012, 3, 322. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Tagliabue, A. The interleukin-1 receptor family. Semin. Immunol. 2013, 25, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Drexler, S.K.; Kong, P.; Inglis, J.; Williams, R.O.; Garlanda, C.; Mantovani, A.; Yazdi, A.S.; Brennan, F.; Feldmann, M.; Foxwell, B.M.J. Sigirr/tir-8 is an inhibitor of toll-like receptor signaling in primary human cells and regulates inflammation in models of rheumatoid arthritis. Arthritis Rheum. 2010, 62, 2249–2261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wu, X.L.; Zhao, Y.F.; Deng, Z.X.; Qian, G.S. Sigirr inhibits toll-like receptor 4, 5, 9-mediated immune responses in human airway epithelial cells. Mol. Biol. Rep. 2011, 38, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Gulen, M.F.; Qin, J.; Yao, J.; Bulek, K.; Kish, D.; Altuntas, C.Z.; Wald, D.; Ma, C.; Zhou, H.; et al. The toll-interleukin-1 receptor member sigirr regulates colonic epithelial homeostasis, inflammation, and tumorigenesis. Immunity 2007, 26, 461–475. [Google Scholar] [CrossRef]
- Garlanda, C.; Riva, F.; Polentarutti, N.; Buracchi, C.; Sironi, M.; De Bortoli, M.; Muzio, M.; Bergottini, R.; Scanziani, E.; Vecchi, A.; et al. Intestinal inflammation in mice deficient in tir8, an inhibitory member of the il-1 receptor family. Proc. Natl. Acad. Sci. USA 2004, 101, 3522–3526. [Google Scholar] [CrossRef]
- Garlanda, C.; Anders, H.J.; Mantovani, A. Tir8/sigirr: An il-1r/tlr family member with regulatory functions in inflammation and t cell polarization. Trends Immunol. 2009, 30, 439–446. [Google Scholar] [CrossRef]
- Olvera, A.; Ballester, M.; Nofrarias, M.; Sibila, M.; Aragon, V. Differences in phagocytosis susceptibility in haemophilus parasuis strains. Vet. Res. 2009, 40, 24. [Google Scholar] [CrossRef]
- Weingartl, H.M.; Sabara, M.; Pasick, J.; van Moorlehem, E.; Babiuk, L. Continuous porcine cell lines developed from alveolar macrophages: Partial characterization and virus susceptibility. J. Virol. Methods 2002, 104, 203–216. [Google Scholar] [CrossRef]
- Yang, J.; Tan, H.L.; Gu, L.Y.; Song, M.L.; Wu, Y.Y.; Peng, J.B.; Lan, Z.B.; Wei, Y.Y.; Hu, T.J. Sophora subprosrate polysaccharide inhibited cytokine/chemokine secretion via suppression of histone acetylation modification and nf-kappab activation in pcv2 infected swine alveolar macrophage. Int. J. Biol. Macromol. 2017, 104, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Tischer, I.; Peters, D.; Rasch, R.; Pociuli, S. Replication of porcine circovirus: Induction by glucosamine and cell cycle dependence. Arch. Virol. 1987, 96, 39–57. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Shang, Y.; Yin, S.; Wang, D.; Cai, J.; Gong, Z.; Serge, M.; Liu, X. A phage-displayed single domain antibody fused to alkaline phosphatase for detection of porcine circovirus type 2. J. Virol. Methods 2015, 213, 84–92. [Google Scholar] [CrossRef]
- Duan, D.; Zhang, S.; Li, X.; Guo, H.; Chen, M.; Zhang, Y.; Han, J.; Lv, Y. Activation of the tlr/myd88/nf-kappab signal pathway contributes to changes in il-4 and il-12 production in piglet lymphocytes infected with porcine circovirus type 2 in vitro. PLoS ONE 2014, 9, e97653. [Google Scholar] [CrossRef]
- Yin, S.; Sun, S.; Yang, S.; Shang, Y.; Cai, X.; Liu, X. Self-assembly of virus-like particles of porcine circovirus type 2 capsid protein expressed from escherichia coli. Virol. J. 2010, 7, 166. [Google Scholar] [CrossRef]
- Du, Q.; Huang, Y.; Wang, T.; Zhang, X.; Chen, Y.; Cui, B.; Li, D.; Zhao, X.; Zhang, W.; Chang, L.; et al. Porcine circovirus type 2 activates pi3k/akt and p38 mapk pathways to promote interleukin-10 production in macrophages via cap interaction of gc1qr. Oncotarget 2016, 7, 17492–17507. [Google Scholar] [CrossRef]
- Qin, Y.; Li, H.H.; Qiao, J.Y. Tlr2/myd88/nf-kappa b signalling pathway regulates il-8 production in porcine alveolar macrophages infected with porcine circovirus 2. J. Gen. Virol. 2016, 97, 445–452. [Google Scholar] [CrossRef]
- Yin, S.H.; Yang, S.L.; Shang, Y.J.; Cai, X.P.; Liu, X.T. Development and optimization of multiplex- pcr for simultaneous detection of porcine pseudorabies virus, porcine parvovirus, and porcine circovirus type 2. Int. J. Appl. Res. Vet. M. 2012, 10, 273–279. [Google Scholar]
- Yang, S.; Shang, Y.; Wang, D.; Yin, S.; Cai, J.; Liu, X. Diagnosis of porcine circovirus type 2 infection with a combination of immunomagnetic beads, single-domain antibody, and fluorescent quantum dot probes. Arch. Virol. 2015, 160, 2325–2334. [Google Scholar] [CrossRef]
- Zhang, H.L.; Liu, C.; Cheng, S.; Wang, X.F.; Li, W.T.; Charreyre, C.; Audonnet, J.C.; He, Q.G. Porcine cd74 is involved in the inflammatory response activated by nuclear factor kappa b during porcine circovirus type 2 (pcv-2) infection. Arch. Virol. 2013, 158, 2285–2295. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zhang, S.; Zhang, Y.; Chen, M.; Lv, Y. Porcine circovirus type 2 increases interleukin-1beta and interleukin-10 production via the myd88-nf-kappa b signaling pathway in porcine alveolar macrophages in vitro. J. Vet. Sci. 2017, 18, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Chae, C. A review of porcine circovirus 2-associated syndromes and diseases. Vet. J. 2005, 169, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Langohr, I. Current state of knowledge on porcine circovirus type 2-associated lesions. Vet. Pathol. 2013, 50, 23–38. [Google Scholar] [CrossRef]
- Chang, H.W.; Jeng, C.R.; Lin, T.L.; Liu, J.J.; Chiou, M.T.; Tsai, Y.C.; Chia, M.Y.; Jan, T.R.; Pang, V.F. Immunopathological effects of porcine circovirus type 2 (pcv2) on swine alveolar macrophages by in vitro inoculation. Vet. Immunol. Immunopathol. 2006, 110, 207–219. [Google Scholar] [CrossRef]
- Li, W.; Liu, S.; Wang, Y.; Deng, F.; Yan, W.; Yang, K.; Chen, H.; He, Q.; Charreyre, C.; Audoneet, J.C. Transcription analysis of the porcine alveolar macrophage response to porcine circovirus type 2. BMC Genom. 2013, 14, 353. [Google Scholar] [CrossRef]
- Hibbs, M.L.; Quilici, C.; Kountouri, N.; Seymour, J.F.; Armes, J.E.; Burgess, A.W.; Dunn, A.R. Mice lacking three myeloid colony-stimulating factors (g-csf, gm-csf, and m-csf) still produce macrophages and granulocytes and mount an inflammatory response in a sterile model of peritonitis. J. Immunol. 2007, 178, 6435–6443. [Google Scholar] [CrossRef]
- Becher, B.; Tugues, S.; Greter, M. Gm-csf: From growth factor to central mediator of tissue inflammation. Immunity 2016, 45, 963–973. [Google Scholar] [CrossRef]
- Dougan, M.; Dranoff, G.; Dougan, S.K. Gm-csf, il-3, and il-5 family of cytokines: Regulators of inflammation. Immunity 2019, 50, 796–811. [Google Scholar] [CrossRef]
- Huffman, J.A.; Hull, W.M.; Dranoff, G.; Mulligan, R.C.; Whitsett, J.A. Pulmonary epithelial cell expression of gm-csf corrects the alveolar proteinosis in gm-csf-deficient mice. J. Clin. Investig. 1996, 97, 649–655. [Google Scholar] [CrossRef]
- Fujiu, K.; Shibata, M.; Nakayama, Y.; Ogata, F.; Matsumoto, S.; Noshita, K.; Iwami, S.; Nakae, S.; Komuro, I.; Nagai, R.; et al. A heart-brain-kidney network controls adaptation to cardiac stress through tissue macrophage activation. Nat. Med. 2017, 23, 611. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Bonecchi, R.; Locati, M. Tuning inflammation and immunity by chemokine sequestration: Decoys and more. Nat. Rev. Immunol. 2006, 6, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Chae, C. Expression of monocyte chemoattractant protein-1 but not interleukin-8 in granulomatous lesions in lymph nodes from pigs with naturally occurring postweaning multisystemic wasting syndrome. Vet. Pathol. 2003, 40, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Chae, C. Expression of monocyte chemoattractant protein-1 and macrophage inflammatory protein-1 in porcine circovirus 2-induced granulomatous inflammation. J. Comp. Pathol. 2004, 131, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.C.; Jeng, C.R.; Hsiao, S.H.; Chang, H.W.; Liu, J.J.; Chang, C.C.; Lin, C.M.; Chia, M.Y.; Pang, V.F. Porcine circovirus type 2 (pcv2) induces cell proliferation, fusion, and chemokine expression in swine monocytic cells in vitro. Vet. Res. 2010, 41, 60. [Google Scholar] [CrossRef] [PubMed]
- Gerszten, R.E.; Garcia-Zepeda, E.A.; Lim, Y.C.; Yoshida, M.; Ding, H.A.; Gimbrone, M.A., Jr.; Luster, A.D.; Luscinskas, F.W.; Rosenzweig, A. Mcp-1 and il-8 trigger firm adhesion of monocytes to vascular endothelium under flow conditions. Nature 1999, 398, 718–723. [Google Scholar] [CrossRef]
- Baggiolini, M.; Dewald, B.; Moser, B. Interleukin-8 and related chemotactic cytokines--cxc and cc chemokines. Adv. Immunol. 1994, 55, 97–179. [Google Scholar]
- Segales, J.; Domingo, M.; Chianini, F.; Majo, N.; Dominguez, J.; Darwich, L.; Mateu, E. Immunosuppression in postweaning multisystemic wasting syndrome affected pigs. Vet. Microbiol. 2004, 98, 151–158. [Google Scholar] [CrossRef]
- Dinarello, C.A. Proinflammatory cytokines. Chest 2000, 118, 503–508. [Google Scholar] [CrossRef]
- Sipos, W.; Duvigneau, J.C.; Willheim, M.; Schilcher, F.; Hartl, R.T.; Hofbauer, G.; Exel, B.; Pietschmann, P.; Schmoll, F. Systemic cytokine profile in feeder pigs suffering from natural postweaning multisystemic wasting syndrome (pmws) as determined by semiquantitative rt-pcr and flow cytometric intracellular cytokine detection. Vet. Immunol. Immunopathol. 2004, 99, 63–71. [Google Scholar] [CrossRef]
- Qin, J.Z.; Qian, Y.C.; Yao, J.H.; Grace, C.; Li, X.X. Sigirr inhibits interleukin-1 receptor- and toll-like receptor 4-mediated signaling through different mechanisms. J. Biol. Chem. 2005, 280, 25233–25241. [Google Scholar] [CrossRef] [PubMed]
- Fawley, J.; Cuna, A.; Menden, H.L.; McElroy, S.; Umar, S.; Welak, S.R.; Gourlay, D.M.; Li, X.; Sampath, V. Single-immunoglobulin interleukin-1-related receptor regulates vulnerability to tlr4-mediated necrotizing enterocolitis in a mouse model. Pediatric Res. 2018, 83, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Garlanda, C.; Riva, F.; Bonavita, E.; Gentile, S.; Mantovani, A. Decoys and regulatory “receptors” of the il-1/toll-like receptor superfamily. Front. Immunol. 2013, 4, 180. [Google Scholar] [CrossRef] [PubMed]
- Wald, D.; Qin, J.; Zhao, Z.; Qian, Y.; Naramura, M.; Tian, L.; Towne, J.; Sims, J.E.; Stark, G.R.; Li, X. Sigirr, a negative regulator of toll-like receptor-interleukin 1 receptor signaling. Nat. Immunol. 2003, 4, 920–927. [Google Scholar] [CrossRef]
- Lv, Y.; Zhang, X.; Sun, Y.; Zhang, S. Activation of nf-kappab contributes to production of pig-major acute protein and serum amyloid a in pigs experimentally infected with porcine circovirus type 2. Res. Vet. Sci 2013, 95, 1235–1240. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Liu, B.; Yin, S.; Shang, Y.; Zhang, X.; Khan, M.U.Z.; Liu, X.; Cai, J. Porcine Circovirus Type 2 Induces Single Immunoglobulin Interleukin-1 Related Receptor (SIGIRR) Downregulation to Promote Interleukin-1β Upregulation in Porcine Alveolar Macrophage. Viruses 2019, 11, 1021. https://doi.org/10.3390/v11111021
Yang S, Liu B, Yin S, Shang Y, Zhang X, Khan MUZ, Liu X, Cai J. Porcine Circovirus Type 2 Induces Single Immunoglobulin Interleukin-1 Related Receptor (SIGIRR) Downregulation to Promote Interleukin-1β Upregulation in Porcine Alveolar Macrophage. Viruses. 2019; 11(11):1021. https://doi.org/10.3390/v11111021
Chicago/Turabian StyleYang, Shunli, Baohong Liu, Shuanghui Yin, Youjun Shang, Xinming Zhang, Muhammad Umar Zafar Khan, Xiangtao Liu, and Jianping Cai. 2019. "Porcine Circovirus Type 2 Induces Single Immunoglobulin Interleukin-1 Related Receptor (SIGIRR) Downregulation to Promote Interleukin-1β Upregulation in Porcine Alveolar Macrophage" Viruses 11, no. 11: 1021. https://doi.org/10.3390/v11111021