Abstract
Bees are important plant pollinators in both natural and agricultural ecosystems. Managed and wild bees have experienced high average annual colony losses, population declines, and local extinctions in many geographic regions. Multiple factors, including virus infections, impact bee health and longevity. The majority of bee-infecting viruses are positive-sense single-stranded RNA viruses. Bee-infecting viruses often cause asymptomatic infections but may also cause paralysis, deformity or death. The severity of infection is governed by bee host immune responses and influenced by additional biotic and abiotic factors. Herein, we highlight studies that have contributed to the current understanding of antiviral defense in bees, including the Western honey bee (Apis mellifera), the Eastern honey bee (Apis cerana) and bumble bee species (Bombus spp.). Bee antiviral defense mechanisms include RNA interference (RNAi), endocytosis, melanization, encapsulation, autophagy and conserved immune pathways including Jak/STAT (Janus kinase/signal transducer and activator of transcription), JNK (c-Jun N-terminal kinase), MAPK (mitogen-activated protein kinases) and the NF-κB mediated Toll and Imd (immune deficiency) pathways. Studies in Dipteran insects, including the model organism Drosophila melanogaster and pathogen-transmitting mosquitos, provide the framework for understanding bee antiviral defense. However, there are notable differences such as the more prominent role of a non-sequence specific, dsRNA-triggered, virus limiting response in honey bees and bumble bees. This virus-limiting response in bees is akin to pathways in a range of organisms including other invertebrates (i.e., oysters, shrimp and sand flies), as well as the mammalian interferon response. Current and future research aimed at elucidating bee antiviral defense mechanisms may lead to development of strategies that mitigate bee losses, while expanding our understanding of insect antiviral defense and the potential evolutionary relationship between sociality and immune function.
1. Introduction
1.1. Bees—Hymenopteran Insects That Play an Important Ecological Role as Plant Pollinators
There are over 4000 bee species in the order Hymenoptera, including those that are social or solitary, native or introduced, managed or wild [1]. Bees are important pollinators of plant species, including agricultural crops (e.g., almonds, apples, cherries, squash, tomatoes) and ecologically important plants. In the United States honey bee pollination is valued at 14.6 billion annually [2] and insect pollination worldwide is valued at $175 billion per year [3]. Due to their abundance and economic importance, most of the research on bee host—virus interactions has focused on honey bees.
Western honey bees (Apis mellifera) are eusocial, cavity nesting bees that are native to Europe, Africa and the Middle East; they were introduced into North America in the late 1600s [4,5,6]. The Eastern honey bee (Apis cerana) is a related but distinct species endemic to Asia and detected in Australia in 2007 [7,8]. Honey bee colonies consist of approximately 35,000 individual bees, including sterile female workers, a few hundred male bees (called drones) and a single reproductive female queen bee [9]. Honey bee colonies typically survive multiple years, while the longevity of individual worker bees depends on their caste (i.e., from six weeks to four months for worker bees, approximately eight weeks for drones and several years for queen bees [9]). The majority of the approximately 2.5 million honey bee colonies in the United States (US) are managed by professional beekeepers and are involved in pollinating the almond crop, which is the largest pollination event in the world [10,11]. Since 2006, US beekeeping operations have suffered approximately 33% annual losses, which is an increase from historic levels of approximately 12–15% [10,12,13,14,15,16,17]. Many biotic and abiotic factors contribute to these losses, including pathogenic infections, mite infestation levels, agrochemical-exposure, management, and lack of quality forage and habitat (reviewed in [18,19,20,21,22,23,24]). Viruses, including deformed wing virus (DWV), are one of the factors that contribute to individual bee and colony deaths.
Bumble bees, including Bombus terrestris and Bombus impatiens, are also important agricultural pollinators of crops such as tomatoes and peppers, as well as blueberries and other ecologically important plant species [25]. Bumble bees are ground nesting bees that live in small annual colonies with distinct solitary and social life cycle phases [26]. Unlike honey bees, bumble bees (B. terrestris) rear one generation per year [27]. This means that the queen survives one year and her reproductive daughters (gynes) start new colonies in the spring after an overwinter period of torpor (called diapause) [28,29]. There are numerous species of bumble bees, some of which have suffered high losses and local extinctions that are partially attributed to habitat destruction and fragmentation, chemical-exposure, pathogens, and climate change, [26,30,31,32,33,34,35,36,37]. The majority of bumble bee host pathogen research has focused on microsporidia (i.e., Nosema bombi) and trypanosomatid (i.e., Crithidia bombi) infections [38,39,40,41,42], though there is a growing body of virus literature, which is featured herein [43,44,45,46,47,48]. Recent metagenomic sequencing analysis of bumble bees (i.e., Bombus terrestris, Bombus cryptarum, and Bombus pascuorum) obtained from several locations in Belgium identified several known bee infecting viruses (e.g., black queen cell virus (BQCV), Varroa destructor virus-1 VDV-1/DWV-B, DWV), including potentially different strains as well as numerous new bee-associated viruses including (+)ssRNA, (−)ssRNA, and dsDNA viruses [49]. Future studies aimed at characterizing the full genome sequences, virion structure, potential pathogenicity, host-specific antiviral responses, and inter-taxa transmission of these viruses will greatly expand our understanding of bee virology [49]. Virus infections of social bees, including honey bees and bumble bees, may impact bee health at the superorganism (i.e., entire colony) and/or individual bee levels. Typically, colony population size is used as a proxy for colony health, whereas pathogen burden, life span, glandular protein content, and queen bee fecundity are used as proxies of individual bee health [12,14,16,50,51,52,53,54,55].
Solitary bees including alfalfa leaf cutter bees (Megachile rotundata), blue orchard or mason bees (Osmia lignaria), and many other native and wild bee species are important plant pollinators. Some are generalist pollinators, whereas others are specialist pollinators that primarily interact with one or just a few plants (reviewed in [56]). Interestingly, numerous studies indicate that agricultural systems that include both managed and native and wild bee species have improved crop yield [57,58,59,60]. Less is known about the health of these bees but in general habitat destruction, pathogenic infections, lack of quality forage, and agrochemical exposure are detrimental to bee health and population size (reviewed in [33]). Therefore, strategies that promote bee health including planting and/or maintaining pollinator forage, maintaining nesting sites (including bare earth for ground nesting bees), and reduced use of chemicals, particularly insecticides, will benefit all bee species. Though more research on the impact of viruses on solitary bees is needed, many studies have shown that these bee species are infected by viruses originally discovered in honey bees [61], as well as viruses and other parasites that may be unique to particular hosts [49]. For example, high throughput sequencing of the metatranscriptomes of eight wild bee species including five solitary bee species (i.e., Andrena cineraria, Osmia bicornis, Osmia cornuta, Andrena fulva, and Andrena haemorrhoa) in Belgium resulted in strong support for bee macula-like 2 virus infection of A. haemorrhoa and detection of several new partial virus genomes including a nege-like virus and a toti-like virus in A. haemorrhoa and O. cornuta, respectively [49].
A current focus of bee virus research is investigating intra- and inter- genera transmission of viruses. Though difficult to investigate, phylogenetic analyses of virus genome sequences obtained from co-foraging bee hosts have indicated that viruses are bidirectionally transmitted between managed and wild bee species [62,63,64,65] (reviewed in [66]). Additional studies are required to determine the extent of virus replication, as opposed to virus prevalence and pathogenesis across bee taxa. Inter-genera virus transmission is likely influenced by virus prevalence and abundance in bee populations, as well as the dynamic composition of bee and forb species in specific geographic regions. In addition, plant-pollinator networks and in turn pathogen transmission between co-foraging bees, are influenced by habitat loss and will likely be influenced by climate change [31,37]. Investigating the co-evolutionary history of specific virus-host pairs, host antiviral immune responses, and viral counter measures in numerous bee species will greatly enhance our understanding of bee virus ecology.
1.2. Bee Viruses
In this review we will use the term “bee virus”, though insect viruses generally have a broad host range and “bee viruses” can infect a variety of bee hosts, as well as ants and mites [67,68,69,70] (reviewed in [66]). Because of their role in agriculture, honey bees (Apis mellifera) are the most investigated bee species and thus the majority of bee-infecting viruses were discovered in honey bee samples. Most bee viruses are positive-sense single-stranded RNA viruses with approximately 30 nm diameter icosahedral capsids. These include Dicistroviruses (black queen cell virus (BQCV), Israeli acute paralysis virus (IAPV)), Iflaviruses (deformed wing virus (DWV), sacbrood virus (SBV), slow bee paralysis virus (SBPV)) and yet-to-be taxonomically classified viruses including chronic bee paralysis virus (CBPV) and the Lake Sinai virus (LSV) group (reviewed in [68,70,71]). Recent sequencing efforts have discovered new bee viruses from additional families (reviewed in [72]) including viruses with negative-sense RNA genomes and enveloped virions [73]. To date, only one bee-infecting DNA virus, Apis mellifera filamentous virus (AmFv), has been described [74]. For a more thorough review of bee virology, see Grozinger and Flenniken [75] and Chen and Siede [70].
Bee viruses are transmitted vertically within species and horizontally, both within species and between different bee genera [62,63,64,70]. Horizontal transmission is facilitated by food transfer (i.e., trophallaxis in social bees) between individual bees within a colony, and between colonies and bee species via the sharing of floral resources (i.e., nectar and pollen) [9,62,76]. Honey bee viruses are also transmitted within and between honey bee colonies by the ectoparasitic mite Varroa destructor (i.e., DWV, IAPV, KBV) [77,78,79,80,81,82,83,84,85]. Several studies suggest that DWV replication in mites and/or mite-mediated virus transmission impacts the diversity of viral genomes at both a geographic scale (i.e., mite induced bottleneck of DWV strains in the Hawaiian Islands [86]) and at the individual bee level [87,88]. Poor honey bee colony health is associated with high mite infestation coupled with DWV infection [83,85,89,90,91,92,93] and the seasonal dynamics of mite infestation and DWV abundance are strongly correlated [12,17,54,85,90,93,94,95]. The potential role of parasite-mediated virus transmission is under-explored for other bee species.
Virus infections in bees are primarily asymptomatic or they may result in deformity, paralysis, and/or death (reviewed in [70,89,96,97]) [98,99,100]. The extent of viral pathogenesis is influenced by biotic and abiotic stressors, including the synergistic negative effects of co-infection with multiple pathogens and/or agrochemical exposure, and governed by co-evolved host-virus interactions [37,101] (reviewed in [75]). The mechanisms of bee antiviral defense, which are described in greater detail below, include conserved immune pathways (i.e., Jak/STAT (Janus kinase/signal transducer and activator of transcription), JNK (c-Jun N-terminal kinase), MAPK (mitogen-activated protein kinases), NF-κB (i.e., Dorsal/Relish) mediated Toll and Imd pathways, RNA-trigged responses (i.e., RNA interference (RNAi) and a non-sequence-specific dsRNA mediated mechanism), autophagy, endocytosis, and melanization) (reviewed in [102,103]) (Figure 1).
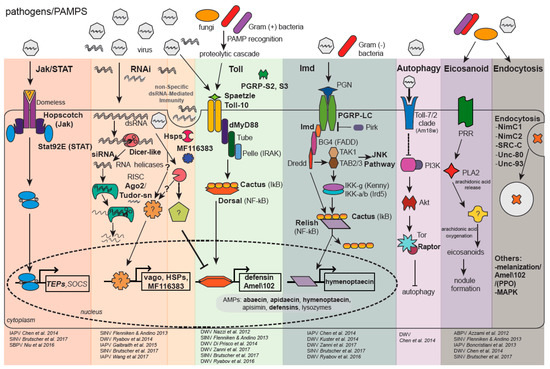
Figure 1.
Honey Bee Immune Pathways—Highlighting Genes Implicated in Antiviral Immune Responses. The honey bee genome encodes major members of insect immune pathways including: Jak/STAT (Janus kinase/Signal Transducer and Activator of Transcription); RNAi (RNA interference); Toll via NF-κB (Nuclear Factor κB/Dorsal); Imd (Immune deficiency) via NF-κB/Relish; JNK (c-Jun N-terminal kinase); and MAPK (Mitogen-Activated Protein Kinases), as well as orthologues of genes involved in the heat shock response (Hsp), autophagy, eicosanoid biosynthesis, endocytosis, melanization, and prophenoloxidase (PPO) response. Bold text indicates genes and proteins differentially expressed in virus-infected honey bees and/or bumble bees. The first step in immune activation is host recognition of pathogen-associated molecular patterns (PAMPs) including viral dsRNA, bacterial peptidoglycans, and fungal β-glucans. In general, the Toll pathway is involved in defense against Gram(+) bacteria and fungi and the Imd pathway is activated by Gram(−) bacteria, but specific host-pathogen interactions are unique. This is particularly true for host—virus interactions since data from fruit-flies, mosquitoes, and honey bees indicate differential activation of immune genes and pathways. The Jak/STAT pathway is activated via ligand binding to the Domeless receptor; while Drosophila melanogaster (Dm) express Domeless ligands (unpaired, upd, upd2, and upd3), a honey bee upd orthologue has not been identified. Following Domeless-ligand binding, Hopscotch Janus kinases are transphosphorylated, leading to phosphorylation and dimerization of STAT92E-like proteins. Activated STATs transcriptionally regulate antimicrobial effectors TEP7 (Thioester-containing protein 7), TEPA, TEPB, and the Jak/STAT inhibitor SOCS (Suppressor of Cytokine Signaling). The honey bee genome also encodes for D-PIAS (Protein Inhibitor of Activated STAT), another inhibitor of the Jak/STAT pathway. The RNAi-pathway is initiated by DmDicer-2 cleavage of viral dsRNA into 21–22 bp siRNAs; AmDicer-like shares ~30% aa identity with DmDicer-2. The siRNAs are then loaded into Ago2 (Argonaute-2), the catalytic component of the RISC (RNA Induced Silencing Complex). A single strand of the siRNA is retained in the RISC and used to specifically target cognate viral genome sequences for cleavage. In addition, DmDicer-2 serves as a dsRNA sensor that mediates a signal transduction cascade resulting in increased expression of DmVago, which suppresses viral replication. AmDicer-like may serve as a dsRNA sensor, as honey bees have a vago orthologue which is up-regulated in DWV-infected honey bees, but not Sindbis-GFP-infected honey bees. In B. terrestris, Vago limits viral infection in fat bodies in a Dicer-dependent manner. Though the mechanism(s) of non-specific dsRNA-mediated antiviral responses in bees require additional characterization, a putative serine/threonine cyclin-dependent kinase (MF 116383) is involved in this virus-limiting response in honey bees. Additionally, several members of the heat shock protein family exhibit increased expression in Sindbis-GFP infected honey bees (i.e., hsp90, activator of hsp90, 60 kda hsp, 10 kda hsp, hsp83-like, hsc70-4, and hsf5), while dsRNA alone resulted in increased expression of hsp90. The Toll pathway is activated by a family of pathogen recognition receptors (PRRs) (e.g., peptidoglycan receptor proteins (PGRPs) and Gram(−) binding proteins) that bind fungal and bacterial PAMPs. The Toll pathway is activated in some insect host-virus combinations, although the activation mechanism is unknown. Following PAMP binding, a serine protease cascade results in cleavage of pro-Spaetzle into mature Spaetzle. The honey bee genome encodes two putative spaetzle orthologues, which bind the membrane-anchored Toll receptor. Toll dimerization results in the recruitment of dMyD88, Tube and Pelle. Pelle is likely involved in degradation of NF-κB inhibitors (e.g., Cactus-1, Cactus-2, Cactus-3), resulting in the release of transcription factors Dorsal-1A and Dorsal-1B. Nuclear translocation of Dorsal results in increased expression of antimicrobial peptides (AMPs) and Amel\102. The Imd pathway is activated by Peptidoglycan recognition protein LC (PGRP-LC) binding to diaminopimelic-containing peptidoglycan of Gram(−) bacteria, followed by activation of the adaptor protein Immune deficiency (Imd), Relish phosphorylation by the IKK complex (IkB kinase) and cleavage of Relish by the caspase Dredd (Death-related ced-3/Nedd2-like). Relish transcriptionally regulates expression of AMPs and other genes involved in antimicrobial defense. The JNK pathway is also activated by TAK (Transforming growth factor-activated kinase 1) and TAB2/3 (TAK binding protein 2 and 3), resulting in AMP expression and/or apoptosis. In Drosophila, binding of vesicular stomatitis virus to the Toll-7 receptor promotes autophagy, likely by inhibiting the PI3/Akt/Tor (phosphatidylinositol 3-kinase/Protein kinase B/Target of rapamycin) pathway which suppresses autophagy. The honey bee genome encodes for one gene in the Toll-7/2 clade, 18-wheeler (Am18w), which shares ~49% aa identity with DmToll-7 and ~45% aa identity with DmToll-2. The role of the Am18w protein in antiviral defense and autophagy in honey bees is unknown. In insects, Eicosanoid biosynthesis begins with the induction of PLA2 (Phospholipase 2) from signal cascades downstream of viral, fungal, or bacterial PAMP recognition. Activated PLA2 hydrolyzes arachidonic acid (AA) from cellular phospholipids. Eicosonoid production likely occurs via oxidation of AA by an unidentified enzyme. Eicosanoids are critical for nodulation and aid in phagocytosis, micro-aggregation, adhesion, and release of prophenoloxidase (PPO) from hemocytes. Experimental evidence also suggests endocytosis, melanization and MAPK pathways are involved in honey bee antiviral defense. Adapted with permission from Brutscher et al., Current Opinion in Insect Science, 2015 [102].
1.3. Bee Virology
The epidemiology of bee viruses has not been thoroughly investigated, though there have been several insightful studies including the German Bee Monitoring Project [90], apiary level surveillance programs carried out by the US Bee Informed Partnership [12] and the Ministry of Agriculture in Spain [104], honey bee colony health and virus prevalence and abundance studies [12,13,14,15,16,17,53,54,55,97,105,106,107,108,109,110], and the ongoing Canadian National Honey Bee Health Survey [111]. These and other longitudinal monitoring studies have been instrumental in beginning to define how honey bee health relates to the prevalence and abundance of viruses, which varies by season, sampling date, and geographical location [54,105]. Several studies indicate that at particular times of the year weak colonies are associated with higher pathogen levels, including IAPV, LSV2, DWV, Nosema ceranae, and mites [12,13,14,15,16,53,54,55,90,95,97,106,109,110,112,113,114], though additional monitoring efforts are required to determine impacts of virus infection at the colony level.
To examine the impact of viruses on individual bees, Bailey, Ball, and others used semi-purified viruses from infected bees/pupae to catalogue the symptoms associated with particular viruses [70,100,115]. Symptomatic virus infections in honey bees include a “hairless” or “greasy” phenotype associated with CBPV, wing deformity and shortened or bloated abdomens caused by DWV, paralysis associated with IAPV and acute bee paralysis virus (ABPV) [116] and complications with larval development due to SBV and BQCV infections [70]. In addition, asymptomatic or covert infections may cause more subtle symptoms, such as the precocious foraging behavior and reduced lifespan associated with DWV-infections [117]. To date, only two infectious bee virus clones, BQCV [118] and DWV [119] have been described and only the DWV clone is currently available. Bee cell culture includes the use of primary cells obtained from embryos, larvae, and adults for short-term studies and a single immortalized cell line [120], which can be difficult to maintain [121]. Therefore, most bee virus research is carried out using viruses isolated from both naturally and experimentally infected bees and/or bee pupae [115]. Isolating viruses from naturally infected bees typically includes several co-purified viruses [115,121,122], though nearly pure virus isolates have been obtained (e.g., LSV2 [112]). Likewise, propagation of DWV and IAPV in pupae has resulted in relatively pure virus preparations [97,123,124,125]. The lack of infectious clones and robust cell culture models has made investigating bee viruses challenging but the use of model viruses, including Sindbis-GFP [126,127] and flock house virus (FHV) [128], and semi-purified virus inocula in individual bee and cultured cell experiments have provided insight into the mechanisms of antiviral defense [61,121,126,127,128,129].
The genomes of several bee species including Apis mellifera [130], Apis cerana [7], Bombus terrestris, Bombus impatiens [131], and Megachile rotundata have been sequenced and partially annotated [132]. Genomic information facilitates the use of molecular techniques (e.g., high through-put sequencing, qRT-PCR, cloning, and RNA interference-mediated gene knock-down) to investigate and understand bee host—virus interactions at both the colony and individual levels.
3. Conclusions
RNA virus infection results in the production of dsRNA molecules (e.g., virus replicative intermediates, RNA secondary structure, dsRNA genomes) in host cells. This viral pathogen PAMP is recognized as non-self by PRRs across diverse taxa, including plants, invertebrates (e.g., oysters, shrimp, nematodes, ticks, fruit flies, sand flies, mosquitos, wasps, and bees) and mammals [176,212,213,214,215,216,217,218,219,220]. Recognition of dsRNA by host PRRs induces distinctive antiviral responses across different hosts. These include virus-specific RNAi in plants, nematodes, and arthropods and non-sequence specific dsRNA-mediated induction of pathways that result in an “antiviral state” that limits virus replication (e.g., mammalian interferon response, C. gigas oyster type I interferon-like response, A. mellifera and B. terrestris virus-limiting responses). The extent to which these pathways are involved varies for each co-evolved virus—host pair. Overall the role of RNAi is greater in plants, fruit-flies, and mosquitos, while the role of other immune pathways including Jak/STAT, JNK, Toll, and non-sequence specific dsRNA-triggered pathways are more important in bees, oysters, and mammals. Importantly, these generalities are only true for the specific species that have been studied including Arabidopsis thaliana, Xanthomonas oryzae, D. melanogaster, A. gambiae, A. mellifera, B. terrestris, C. gigas, Mus musculus, Homo sapiens, and others.
The antiviral response(s) that are most important for any particular host-virus pair cannot be assumed based on broad organismal classification (e.g., insects, invertebrates, mammals)—each must be empirically determined. Since all host-virus interactions are inexorably complicated by the history of conflict and evolution shared (or not shared in the case of model viruses) by the host and a virus. These unique histories are reflected in the differential transcriptional responses and extent of parallel activation and/or cross-talk between host immune pathways, as well as identification of the virus-evolved counter defense mechanisms, including suppressors of RNAi and other immune pathways [140,221].
The parallels that exist between the antiviral responses, including the general, non-sequence specific dsRNA-triggered induction of an antiviral state, in organisms separated by large evolutionary distances including honey bees, bumble bees, sand flies, shrimp, oysters, and mammals, are very intriguing. Furthermore, it is interesting to hypothesize that in social bees this response may have evolved to rapidly respond to viruses and limit their transmission in the crowded hive environment and in the context of behaviors (e.g., trophallaxis) that promote virus transmission between individuals within the super-organism. Investigation of antiviral defense in some of the thousands of under-explored bee species, will further our understanding of the general and specific mechanisms that bees have evolved to combat specific viruses. This short review highlights studies that have contributed to our current understanding of bee antiviral defense mechanisms. Under-explored, burgeoning research areas include elucidation of the roles of alternative splicing [127,222], epigenetic regulation [125,223], and transgenerational immune priming in bee antiviral defense [224,225]. Further examination of antiviral RNAi, including immune memory as a consequence of RNA virus integration into the bee genome [226] and potential transposon-mediated amplification of virus-targeting secondary RNAs (as described in Drosophila melanogaster [182,183,184]) is also important. Lessons learned from evolutionary distant organisms, including those described in this special issue of Viruses focused on “Antiviral Defense in Invertebrates”, may help guide these studies.
Funding
This review article received no external funding.
Acknowledgments
The Flenniken Laboratory is supported by the National Sciences Foundation CAREER Program, the United States Department of Agriculture National Institute of Food and Agriculture, Agriculture and Food Research Initiative (USDA-NIFA-AFRI) Program, Montana Department of Agriculture Specialty Crop Block Grant Program, Hatch Multistate Funding (NC-1173), and Project Apis m. We would like to thank members of the Flenniken Laboratory, Sandra Barroso Arevalo (Universidad Complutense de Madrid), Laura Brutscher (University of California-Davis), and Francesco Nazzi (Università di Udine) for reviewing this manuscript prior to publication.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript and in the decision to publish the results.
References
- Michener, C.D. The Bees of the World, 2nd ed.; John Hopkins University Press: Baltimore, MD, USA; London, UK, 2007. [Google Scholar]
- Calderone, N.W. Insect pollinated crops, insect pollinators and us agriculture: Trend analysis of aggregate data for the period 1992–2009. PLoS ONE 2012, 7, e37235. [Google Scholar] [CrossRef] [PubMed]
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Cridland, J.M.; Tsutsui, N.D.; Ramirez, S.R. The complex demographic history and evolutionary origin of the western honey bee, Apis mellifera. Genome Biol. Evol. 2017, 9, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, W.S. A history of the introduction of honey bee races into the United-States 2. Am. Bee J. 1989, 129, 664–667. [Google Scholar]
- Han, F.; Wallberg, A.; Webster, M.T. From where did the western honeybee (Apis mellifera) originate? Ecol. Evol. 2012, 2, 1949–1957. [Google Scholar] [CrossRef] [PubMed]
- Diao, Q.; Sun, L.; Zheng, H.; Zeng, Z.; Wang, S.; Xu, S.; Zheng, H.; Chen, Y.; Shi, Y.; Wang, Y.; et al. Genomic and transcriptomic analysis of the asian honeybee Apis cerana provides novel insights into honeybee biology. Sci. Rep. 2018, 8, 822. [Google Scholar] [CrossRef] [PubMed]
- Government, T.A. Status of the Asian Honey Bee in Mainland Australia. Available online: http://www.agriculture.gov.au/pests-diseases-weeds/bees/the-asian-honey-bee-in-australia (accessed on 2 July 2018).
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1991. [Google Scholar]
- Van Engelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef] [PubMed]
- Flenniken, M.L. Almond Almanac; Almond Board of California: Modesto, CA, USA, 2014. [Google Scholar]
- Traynor, K.S.; Rennich, K.; Forsgren, E.; Rose, R.; Pettis, J.; Kunkel, G.; Madella, S.; Evans, J.; Lopez, D.; van Engelsdorp, D. Multiyear survey targeting disease incidence in US honey bees. Apidologie 2016, 47, 325–347. [Google Scholar] [CrossRef]
- Spleen, A.M.; Lengerich, E.J.; Rennich, K.; Caron, D.; Rose, R.; Pettis, J.S.; Henson, M.; Wilkes, J.T.; Wilson, M.; Stitzinger, J.; et al. A national survey of managed honey bee 2011–2012 winter colony losses in the United States: Results from the bee informed partnership. J. Apicult. Res. 2015, 52, 44–53. [Google Scholar] [CrossRef]
- Steinhauer, N.A.; Rennich, K.; Wilson, M.E.; Caron, D.M.; Lengerich, E.J.; Pettis, J.S.; Rose, R.; Skinner, J.A.; Tarpy, D.R.; Wilkes, J.T.; et al. A national survey of managed honey bee 2012–2013 annual colony losses in the USA: Results from the bee informed partnership. J. Apicult. Res. 2015, 53, 1–18. [Google Scholar] [CrossRef]
- Van Engelsdorp, D.; Caron, D.; Hayes, J.; Underwood, R.; Henson, M.; Rennich, K.; Spleen, A.; Andree, M.; Snyder, R.; Lee, K.; et al. A national survey of managed honey bee 2010–2011 winter colony losses in the USA: Results from the bee informed partnership. J. Apicult. Res. 2015, 51, 115–124. [Google Scholar] [CrossRef]
- Van Engelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.; et al. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar]
- Van Engelsdorp, D.; Hayes, J.; Underwood, R.M.; Pettis, J. A survey of honey bee colony losses in the U.S., fall 2007 to spring 2008. PLoS ONE 2008, 3, e4071. [Google Scholar]
- McMenamin, A.J.; Brutscher, L.M.; Glenny, W.; Flenniken, M.L. Abiotic and biotic factors affecting the replication and pathogenicity of bee viruses. Curr. Opin. Insect Sci. 2016, 16, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Vaudo, A.D.; Tooker, J.F.; Grozinger, C.M.; Patch, H.M. Bee nutrition and floral resource restoration. Curr. Opin. Insect Sci. 2015, 10, 133–141. [Google Scholar] [CrossRef] [PubMed]
- DeGrandi-Hoffman, G.; Chen, Y. Nutrition, immunity and viral infections in honey bees. Curr. Opin. Insect Sci. 2015, 10, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Barron, A.B. Death of the bee hive: Understanding the failure of an insect society. Curr. Opin. Insect Sci. 2015, 10, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M. Honey bee toxicology. Annu. Rev. Entomol. 2015, 60, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M.; Ellis, M.D.; Mullin, C.A.; Frazier, M. Pesticides and honey bee toxicity—USA. Apidologie 2010, 41, 312–331. [Google Scholar] [CrossRef]
- Simone-Finstrom, M.; Li-Byarlay, H.; Huang, M.H.; Strand, M.K.; Rueppell, O.; Tarpy, D.R. Migratory management and environmental conditions affect lifespan and oxidative stress in honey bees. Sci. Rep. 2016, 6, 810. [Google Scholar] [CrossRef] [PubMed]
- Velthuis, H.H.W.; van Doorn, A. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef]
- Goulson, D. Bumblebees: Their Behaviour and Ecology; Oxford University Press: Oxford, UK; New York, NY, USA, 2003. [Google Scholar]
- Goulson, D. Effects of introduced bees on native ecosystems. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 1–26. [Google Scholar] [CrossRef]
- Amsalem, E.; Galbraith, D.A.; Cnaani, J.; Teal, P.E.A.; Grozinger, C.M. Conservation and modification of genetic and physiological toolkits underpinning diapause in bumble bee queens. Mol. Ecol. 2015, 24, 5596–5615. [Google Scholar] [CrossRef] [PubMed]
- Amsalem, E.; Grozinger, C.M.; Padilla, M.; Hefetz, A. The physiological and genomic bases of bumble bee social behaviour. In Genomics, Physiology and Behaviour of Social Insects; Elsevier: London, UK, 2015; Volume 48, pp. 37–93. [Google Scholar]
- Owen, R.E.; Otterstatter, M.C.; Carter, R.V.; Farmer, A.; Colla, S.R.; O’Toole, N. Significant expansion of the distribution of the bumble bee Bombus moderatus (hymenoptera: Apidae) in alberta over 20 years. Can. J. Zool.-Revue Can. Zool. 2012, 90, 133–138. [Google Scholar] [CrossRef]
- Burkle, L.A.; Marlin, J.C.; Knight, T.M. Plant-pollinator interactions over 120 years: Loss of species, co-occurrence, and function. Science 2013, 339, 1611–1615. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botias, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [PubMed]
- Hatten, T.D.; Looney, C.; Strange, J.P.; Bosque-Perez, N.A. Bumble bee fauna of palouse prairie: Survey of native bee pollinators in a fragmented ecosystem. J. Insect Sci. 2013, 13, 26. [Google Scholar] [CrossRef] [PubMed]
- Woodard, S.H. Bumble bee ecophysiology: Integrating the changing environment and the organism. Curr. Opin. Insect Sci. 2017, 22, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant-pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Meeus, I.; Pisman, M.; Smagghe, G.; Piot, N. Interaction effects of different drivers of wild bee decline and their influence on host-pathogen dynamics. Curr. Opin. Insect Sci. 2018, 26, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.; Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef] [PubMed]
- Barribeau, S.M.; Schmid-Hempel, P. Qualitatively different immune response of the bumblebee host, Bombus terrestris, to infection by different genotypes of the trypanosome gut parasite, Crithidia bombi. Infect. Genet. Evol. 2013, 20, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Cordes, N.; Huang, W.-F.; Strange, J.P.; Cameron, S.A.; Griswold, T.L.; Lozier, J.D.; Solter, L.F. Interspecific geographic distribution and variation of the pathogens Nosema bombi and crithidia species in United States bumble bee populations. J. Invertebr. Pathol. 2012, 109, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Barribeau, S.M.; Sadd, B.M.; du Plessis, L.; Schmid-Hempel, P. Gene expression differences underlying genotype-by-genotype specificity in a host–parasite system. Proc. Natl. Acad. Sci. USA 2014, 111, 3496–3501. [Google Scholar] [CrossRef] [PubMed]
- Imhoof, B.; Schmid-Hempel, P. Colony success of the bumble bee, Bombus terrestris, in relation to infections by two protozoan parasites, Crithidia bombi and Nosema bombi. Insectes Soc. 2014, 46, 233–238. [Google Scholar] [CrossRef]
- Parmentier, L.; Smagghe, G.; de Graaf, D.C.; Meeus, I. Varroa destructor macula-like virus, lake sinai virus and other new RNA viruses in wild bumblebee hosts (Bombus pascuorum, Bombus lapidarius, and Bombus pratorum). J. Invertebr. Pathol. 2016, 134, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Smagghe, G.; De Coninck, D.I.M.; Van Nieuwerburgh, F.; Deforce, D.; Meeus, I. In vivo study of dicer-2-mediated immune response of the small interfering rna pathway upon systemic infections of virulent and avirulent viruses in Bombus terrestris. Insect Biochem. Mol. Biol. 2016, 70, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Meeus, I.; Brown, M.J.F.; de Graaf, D.C.; Smagghe, G. Effects of invasive parasites on bumble bee declines. Conserv. Biol. 2011, 25, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Meeus, I.; de Miranda, J.R.; de Graaf, D.C.; Wäckers, F.; Smagghe, G. Effect of oral infection with kashmir bee virus and israeli acute paralysis virus on bumblebee (Bombus terrestris) reproductive success. J. Invertebr. Pathol. 2014, 121, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Piot, N.; Snoeck, S.; Vanlede, M.; Smagghe, G.; Meeus, I. The effect of oral administration of dsrna on viral replication and mortality in Bombus terrestris. Viruses 2015, 7, 3172–3185. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Cappelle, K.; de Miranda, J.R.; Smagghe, G.; Meeus, I. Analysis of reference gene stability after israeli acute paralysis virus infection in bumblebees Bombus terrestris. J. Invertebr. Pathol. 2014, 115, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Schoonvaere, K.; Smagghe, G.; Francis, F.; de Graaf, D.C. Study of the metatranscriptome of eight social and solitary wild bee species reveals novel viruses and bee parasites. Front. Microbiol. 2018, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Sagili, R.R.; Burgett, D.M. Evaluating Honey Bee Colonies for Pollination: A Guide for Commercial Growers and Beekeepers; Oregon State University Extension Service: Corvallis, OR, USA, 2011; pp. 1–8. [Google Scholar]
- Amiri, E.; Meixner, M.; Nielsen, S.L.; Kryger, P. Four categories of viral infection describe the health status of honey bee colonies. PLoS ONE 2015, 10, e0140272. [Google Scholar] [CrossRef] [PubMed]
- Pettis, J.S.; Rice, N.; Joselow, K.; vanEngelsdorp, D.; Chaimanee, V. Colony failure linked to low sperm viability in honey bee (Apis mellifera) queens and an exploration of potential causative factors. PLoS ONE 2016, 11, e0147220. [Google Scholar]
- Nguyen, B.K.; Ribière, M.; vanEngelsdorp, D.; Snoeck, C.; Saegerman, C.; Kalkstein, A.L.; Schurr, F.; Brostaux, Y.; Faucon, J.-P.; Haubruge, E. Effects of honey bee virus prevalence, Varroa destructor load and queen condition on honey bee colony survival over the winter in Belgium. J. Apicult. Res. 2015, 50, 195–202. [Google Scholar] [CrossRef]
- Glenny, W.; Cavigli, I.; Daughenbaugh, K.F.; Radford, R.; Kegley, S.E.; Flenniken, M.L. Honey bee (Apis mellifera) colony health and pathogen composition in migratory beekeeping operations involved in California almond pollination. PLoS ONE 2017, 12, e0182814. [Google Scholar] [CrossRef] [PubMed]
- Cavigli, I.; Daughenbaugh, K.F.; Martin, M.; Lerch, M.; Banner, K.; Garcia, E.; Brutscher, L.M.; Flenniken, M.L. Pathogen prevalence and abundance in honey bee colonies involved in almond pollination. Apidologie 2016, 47, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Brosi, B.J. Pollinator specialization: From the individual to the community. New Phytol. 2016, 210, 1190–1194. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Greenleaf, S.S.; Kremen, C. Wild bees enhance honey bees’ pollination of hybrid sunflower. Proc. Natl. Acad. Sci. USA 2006, 103, 13890–13895. [Google Scholar] [CrossRef] [PubMed]
- Brittain, C.; Williams, N.; Kremen, C.; Klein, A.M. Synergistic effects of non-apis bees and honey bees for pollination services. Proc. R. Soc. B-Biol. Sci. 2013, 280, 20122767. [Google Scholar] [CrossRef] [PubMed]
- Winfree, R.; Williams, N.M.; Dushoff, J.; Kremen, C. Native bees provide insurance against ongoing honey bee losses. Ecol. Lett. 2007, 10, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Dolezal, A.G.; Hendrix, S.D.; Scavo, N.A.; Carrillo-Tripp, J.; Harris, M.A.; Wheelock, M.J.; O’Neal, M.E.; Toth, A.L. Honey bee viruses in wild bees: Viral prevalence, loads, and experimental inoculation. PLoS ONE 2016, 11, e0166190. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Levitt, A.L.; Rajotte, E.G.; Holmes, E.C.; Ostiguy, N.; vanEngelsdorp, D.; Lipkin, W.I.; Depamphilis, C.W.; Toth, A.L.; Cox-Foster, D.L. RNA viruses in hymenopteran pollinators: Evidence of inter-taxa virus transmission via pollen and potential impact on non-apis hymenopteran species. PLoS ONE 2010, 5, e14357. [Google Scholar] [CrossRef] [PubMed]
- McMahon, D.P.; Fürst, M.A.; Caspar, J.; Theodorou, P.; Brown, M.J.F.; Paxton, R.J. A sting in the spit: Widespread cross-infection of multiple rna viruses across wild and managed bees. J. Anim. Ecol. 2015, 84, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Fürst, M.A.; McMahon, D.P.; Osborne, J.L.; Paxton, R.J.; Brown, M.J.F. Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature 2014, 506, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Levitt, A.L.; Singh, R.; Cox-Foster, D.L.; Rajotte, E.; Hoover, K.; Ostiguy, N.; Holmes, E.C. Cross-species transmission of honey bee viruses in associated arthropods. Virus Res. 2013, 176, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Tehel, A.; Brown, M.J.; Paxton, R.J. Impact of managed honey bee viruses on wild bees. Curr. Opin. Virol. 2016, 19, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Bigot, D.; Dalmon, A.; Roy, B.; Hou, C.; Germain, M.; Romary, M.; Deng, S.; Diao, Q.; Weinert, L.A.; Cook, J.M.; et al. The discovery of halictivirus resolves the sinaivirus phylogeny. J. Gen. Virol. 2017, 98, 2864–2875. [Google Scholar] [CrossRef] [PubMed]
- Brutscher, L.M.; McMenamin, A.J.; Flenniken, M.L. The buzz about honey bee viruses. PLoS Pathog. 2016, 12, e1005757. [Google Scholar] [CrossRef] [PubMed]
- Boncristiani, H.F.; Di Prisco, G.; Pettis, J.S.; Hamilton, M.; Chen, Y. Molecular approaches to the analysis of deformed wing virus replication and pathogenesis in the honey bee, Apis mellifera. Virol. J. 2009, 6, 221. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Siede, R. Honey bee viruses. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2007; Volume 70, pp. 33–80. [Google Scholar]
- Genersch, E.; Aubert, M. Emerging and re-emerging viruses of the honey bee (Apis mellifera L.). Vet. Res. 2010, 41, 54. [Google Scholar] [CrossRef] [PubMed]
- McMenamin, A.J.; Flenniken, M.L. Recently identified bee viruses and their impact on bee pollinators. Curr. Opin. Insect Sci. 2018, 26, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Remnant, E.J.; Shi, M.; Buchmann, G.; Blacquiere, T.; Holmes, E.C.; Beekman, M.; Ashe, A. A diverse range of novel rna viruses in geographically distinct honey bee populations. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, L.; Cornman, S.; Hartmann, U.; Cousserans, F.; Evans, J.; de Miranda, J.; Neumann, P. The Apis mellifera filamentous virus genome. Viruses 2015, 7, 3798–3815. [Google Scholar] [CrossRef] [PubMed]
- Grozinger, C.M.; Flenniken, M.L. Bee viruses: Ecology, pathogenicity, and impacts. Annu. Rev. Entomol. 2018, in press. [Google Scholar]
- Mazzei, M.; Carrozza, M.L.; Luisi, E.; Forzan, M.; Giusti, M.; Sagona, S.; Tolari, F.; Felicioli, A. Infectivity of dwv associated to flower pollen: Experimental evidence of a horizontal transmission route. PLoS ONE 2014, 9, e113448. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Evans, J.; Feldlaufer, M. Horizontal and vertical transmission of viruses in the honey bee, Apis mellifera. J. Invertebr. Pathol. 2006, 92, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Shen, M. Intricate transmission routes and interactions between picorna-like viruses (kashmir bee virus and sacbrood virus) with the honeybee host and the parasitic varroa mite. J. Gen. Virol. 2005, 86, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Yang, X.; Cox-Foster, D.; Cui, L. The role of varroa mites in infections of kashmir bee virus (KBV) and deformed wing virus (DWV) in honey bees. Virology 2005, 342, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Schroder, M.; Gisder, S.; Genersch, E. Vertical-transmission routes for deformed wing virus of honeybees (Apis mellifera). J. Gen. Virol. 2007, 88, 2329–2336. [Google Scholar] [CrossRef] [PubMed]
- Amiri, E.; Meixner, M.; Büchler, R.; Kryger, P. Chronic bee paralysis virus in honeybee queens: Evaluating susceptibility and infection routes. Viruses 2014, 6, 1188–1201. [Google Scholar] [CrossRef] [PubMed]
- Amiri, E.; Kryger, P.; Meixner, M.D.; Strand, M.K.; Tarpy, D.R.; Rueppell, O. Quantitative patterns of vertical transmission of deformed wing virus in honey bees. PLoS ONE 2018, 13, e0195283. [Google Scholar] [CrossRef] [PubMed]
- Gisder, S.; Aumeier, P.; Genersch, E. Deformed wing virus: Replication and viral load in mites (Varroa destructor). J. Gen. Virol. 2009, 90, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Nazzi, F.; Le Conte, Y. Ecology of Varroa destructor, the major ectoparasite of the western honey bee, Apis mellifera. Annu. Rev. Entomol. 2016, 61, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Di Prisco, G.; Annoscia, D.; Margiotta, M.; Ferrara, R.; Varricchio, P.; Zanni, V.; Caprio, E.; Nazzi, F.; Pennacchio, F. A mutualistic symbiosis between a parasitic mite and a pathogenic virus undermines honey bee immunity and health. Proc. Natl. Acad. Sci. USA 2016, 113, 3203–3208. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Wood, G.R.; Fannon, J.M.; Moore, J.D.; Bull, J.C.; Chandler, D.; Mead, A.; Burroughs, N.; Evans, D.J. A virulent strain of deformed wing virus (DWV) of honeybees (Apis mellifera) prevails after Varroa destructor-mediated, or in vitro, transmission. PLoS Pathog. 2014, 10, e1004230. [Google Scholar] [CrossRef] [PubMed]
- McMahon, D.P.; Natsopoulou, M.E.; Doublet, V.; Fürst, M.; Weging, S.; Brown, M.J.F.; Gogol-Döring, A.; Paxton, R.J. Elevated virulence of an emerging viral genotype as a driver of honeybee loss. Proc. R. Soc. B-Biol. Sci. 2016, 283, 20160811. [Google Scholar] [CrossRef] [PubMed]
- De Miranda, J.R.; Genersch, E. Deformed wing virus. J. Invertebr. Pathol. 2010, 103, S48–S61. [Google Scholar] [CrossRef] [PubMed]
- Genersch, E.; von der Ohe, W.; Kaatz, H.; Schroeder, A.; Otten, C.; Büchler, R.; Berg, S.; Ritter, W.; Mühlen, W.; Gisder, S.; et al. The german bee monitoring project: A long term study to understand periodically high winter losses of honey bee colonies. Apidologie 2010, 41, 332–352. [Google Scholar] [CrossRef]
- Annoscia, D.; Brown, S.; Di Prisco, G.; De Paoli, E.; Del Fabbro, S.; Zanni, V.; Galbraith, D.; Caprio, E.; Grozinger, C.M.; Pennacchio, F.; et al. Haemolymph removal by the parasite Varroa destructor can trigger the proliferation of the deformed wing virus in mite infested bees (Apis mellifera), contributing to enhanced pathogen virulence. bioRxiv 2018, 257667. [Google Scholar] [CrossRef]
- Nazzi, F.; Pennacchio, F. Disentangling multiple interactions in the hive ecosystem. Trends Parasitol. 2014, 30, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Nazzi, F.; Brown, S.P.; Annoscia, D.; Del Piccolo, F.; Di Prisco, G.; Varricchio, P.; Della Vedova, G.; Cattonaro, F.; Caprio, E.; Pennacchio, F. Synergistic parasite-pathogen interactions mediated by host immunity can drive the collapse of honeybee colonies. PLoS Pathog. 2012, 8, e1002735. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Steinhauer, N.; Travis, D.A.; Meixner, M.D.; Deen, J.; vanEngelsdorp, D. Honey bee surveillance: A tool for understanding and improving honey bee health. Curr. Opin. Insect Sci. 2015, 10, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Asensio, I.; Vicente-Rubiano, M.; Munoz, M.J.; Fernandez-Carrion, E.; Sanchez-Vizcaino, J.M.; Carballo, M. Importance of ecological factors and colony handling for optimizing health status of apiaries in mediterranean ecosystems. PLoS ONE 2016, 11, e0164205. [Google Scholar] [CrossRef] [PubMed]
- Olivier, V.; Blanchard, P.; Chaouch, S.; Lallemand, P.; Schurr, F.; Celle, O.; Dubois, E.; Tordo, N.; Thiéry, R.; Houlgatte, R.; et al. Molecular characterisation and phylogenetic analysis of chronic bee paralysis virus, a honey bee virus. Virus Res. 2008, 132, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Pettis, J.S.; Corona, M.; Chen, W.P.; Li, C.J.; Spivak, M.; Visscher, P.K.; DeGrandi-Hoffman, G.; Boncristiani, H.; Zhao, Y.; et al. Israeli acute paralysis virus: Epidemiology, pathogenesis and implications for honey bee health. PLoS Pathog. 2014, 10, e1004261. [Google Scholar] [CrossRef] [PubMed]
- Bailey, L.; Fernando, E.F. Effects of sacbrood virus on adult honey-bees. Ann. Appl. Biol. 1972, 72, 27–35. [Google Scholar] [CrossRef]
- Bailey, L. Acute bee-paralysis virus in adult honey bees injected with sacbrood virus. Virology 1967, 33, 368. [Google Scholar] [CrossRef]
- Bailey, L. Honey bee pathology. Annu. Rev. Entomol. 1968, 13, 191–212. [Google Scholar] [CrossRef]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [PubMed]
- Brutscher, L.M.; Daughenbaugh, K.F.; Flenniken, M.L. Antiviral defense mechanisms in honey bees. Curr. Opin. Insect Sci. 2015, 10, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Spain Ministry of Agriculture—Ministerio de Agricultura y Pesca. Informe de Resultados del Programa de Vigilancia Sobre las Pérdidas de Colonias de Abejas 2012–2015; Spain Ministry of Agriculture: Madrid, Spain, 2016. [Google Scholar]
- Runckel, C.; Flenniken, M.L.; Engel, J.C.; Ruby, J.G.; Ganem, D.; Andino, R.; DeRisi, J.L. Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses, nosema, and crithidia. PLoS ONE 2011, 6, e20656. [Google Scholar] [CrossRef] [PubMed]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.K.; Mignon, J.; Laget, D.; de Graaf, D.C.; Jacobs, F.J.; vanEngelsdorp, D.; Brostaux, Y.; Saegerman, C.; Haubruge, E. Honey bee colony losses in Belgium during the 2008–2009 winter. J. Apicult. Res. 2015, 49, 337–339. [Google Scholar] [CrossRef]
- Lee, K.V.; Steinhauer, N.; Rennich, K.; Wilson, M.E.; Tarpy, D.R.; Caron, D.M.; Rose, R.; Delaplane, K.S.; Baylis, K.; Lengerich, E.J.; et al. A national survey of managed honey bee 2013–2014 annual colony losses in the USA. Apidologie 2015, 46, 292–305. [Google Scholar] [CrossRef]
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; vanEngelsdorp, D.; Evans, J.D. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 7, e43562. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Canadian National Honey Bee Health Survey, GPRC National Bee Diagnostic Centre. Available online: https://www.gprc.ab.ca/doc.php?d=2016NHBHS (accessed on 23 May 2018).
- Daughenbaugh, K.F.; Martin, M.; Brutscher, L.M.; Cavigli, I.; Garcia, E.; Lavin, M.; Flenniken, M.L. Honey bee infecting lake sinai viruses. Viruses 2015, 7, 3285–3309. [Google Scholar] [CrossRef] [PubMed]
- Van der Zee, R.; Pisa, L.; Andonov, S.; Brodschneider, R.; Charriere, J.-D.; Chlebo, R.; Coffey, M.F.; Crailsheim, K.; Dahle, B.; Gajda, A.; et al. Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey, for the winters of 2008–2209 and 2009–2010. J. Apicult. Res. 2015, 51, 100–114. [Google Scholar] [CrossRef]
- Van der Zee, R.; Gray, A.; Pisa, L.; de Rijk, T. An observational study of honey bee colony winter losses and their association with Varroa destructor, neonicotinoids and other risk factors. PLoS ONE 2015, 10, e0131611. [Google Scholar] [CrossRef] [PubMed]
- De Miranda, J.R.; Bailey, L.; Ball, B.V.; Blanchard, P.; Budge, G.E.; Chejanovsky, N.; Chen, Y.-P.; Gauthier, L.; Genersch, E.; de Graaf, D.C.; et al. Standard methods for virus research in Apis mellifera. J. Apicult. Res. 2015, 52, 1–56. [Google Scholar] [CrossRef]
- De Miranda, J.R.; Cordoni, G.; Budge, G. The acute bee paralysis virus–kashmir bee virus–israeli acute paralysis virus complex. J. Invertebr. Pathol. 2010, 103, S30–S47. [Google Scholar] [CrossRef] [PubMed]
- Benaets, K.; Van Geystelen, A.; Cardoen, D.; De Smet, L.; de Graaf, D.C.; Schoofs, L.; Larmuseau, M.H.D.; Brettell, L.E.; Martin, S.J.; Wenseleers, T. Covert deformed wing virus infections have long-term deleterious effects on honeybee foraging and survival. Proc. R. Soc. B-Biol. Sci. 2017, 284, 20162149. [Google Scholar] [CrossRef] [PubMed]
- Benjeddou, M.; Leat, N.; Allsopp, M.; Davison, S. Development of infectious transcripts and genome manipulation of black queen-cell virus of honey bees. J. Gen. Virol. 2002, 83, 3139–3146. [Google Scholar] [CrossRef] [PubMed]
- Lamp, B.; Url, A.; Seitz, K.; Eichhorn, J.; Riedel, C.; Sinn, L.J.; Indik, S.; Köglberger, H.; Rümenapf, T. Construction and rescue of a molecular clone of deformed wing virus (dwv). PLoS ONE 2016, 11, e0164639. [Google Scholar] [CrossRef] [PubMed]
- Goblirsch, M.J.; Spivak, M.S.; Kurtti, T.J. A cell line resource derived from honey bee (Apis mellifera) embryonic tissues. PLoS ONE 2013, 8, e69831. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Tripp, J.; Dolezal, A.G.; Goblirsch, M.J.; Miller, W.A.; Toth, A.L.; Bonning, B.C. In vivo and in vitro infection dynamics of honey bee viruses. Sci. Rep. 2016, 6, S50. [Google Scholar] [CrossRef] [PubMed]
- Boncristiani, H.F.; Evans, J.D.; Chen, Y.; Pettis, J.; Murphy, C.; Lopez, D.L.; Simone-Finstrom, M.; Strand, M.; Tarpy, D.R.; Rueppell, O. In vitro infection of pupae with israeli acute paralysis virus suggests disturbance of transcriptional homeostasis in honey bees (Apis mellifera). PLoS ONE 2013, 8, e73429. [Google Scholar] [CrossRef] [PubMed]
- Skubnik, K.; Novacek, J.; Fuzik, T.; Pridal, A.; Paxton, R.J.; Plevka, P. Structure of deformed wing virus, a major honey bee pathogen. Proc. Natl. Acad. Sci. USA 2017, 114, 3210–3215. [Google Scholar] [CrossRef] [PubMed]
- Organtini, L.J.; Shingler, K.L.; Ashley, R.E.; Capaldi, E.A.; Durrani, K.; Dryden, K.A.; Makhov, A.M.; Conway, J.F.; Pizzorno, M.C.; Hafenstein, S. Honey bee deformed wing virus structures reveal that conformational changes accompany genome release. J. Virol. 2017, 91, e01795–16. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, D.A.; Yang, X.; Nino, E.L.; Yi, S.; Grozinger, C. Parallel epigenomic and transcriptomic responses to viral infection in honey bees (Apis mellifera). PLoS Pathog. 2015, 11, e1004713. [Google Scholar] [CrossRef] [PubMed]
- Brutscher, L.M.; Daughenbaugh, K.F.; Flenniken, M.L. Virus and dsrna-triggered transcriptional responses reveal key components of honey bee antiviral defense. Sci. Rep. 2017, 7, 6448. [Google Scholar] [CrossRef] [PubMed]
- Flenniken, M.L.; Andino, R. Non-specific dsrna-mediated antiviral response in the honey bee. PLoS ONE 2013, 8, e77263. [Google Scholar] [CrossRef] [PubMed]
- O'Neal, S.T.; Swale, D.R.; Anderson, T.D. Atp-sensitive inwardly rectifying potassium channel regulation of viral infections in honey bees. Sci. Rep. 2017, 7, 614. [Google Scholar] [CrossRef] [PubMed]
- O'Neal, S.T.; Brewster, C.C.; Bloomquist, J.R.; Anderson, T.D. Amitraz and its metabolite modulate honey bee cardiac function and tolerance to viral infection. J. Invertebr. Pathol. 2017, 149, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Maleszka, R.; Hartfelder, K.; Worley, K.C.; Amdam, G.V.; Bitondi, M.M.G.; Collins, A.M.; Cristino, A.S.; Michael, H.; Lattorff, G.; et al. Insights into social insects from the genome of the honeybee Apis mellifera. Nature 2006, 443, 931–949. [Google Scholar]
- Sadd, B.M.; Barribeau, S.M.; Bloch, G.; de Graaf, D.C.; Dearden, P.; Elsik, C.G.; Gadau, J.; Grimmelikhuijzen, C.J.P.; Hasselmann, M.; Lozier, J.D.; et al. The genomes of two key bumblebee species with primitive eusocial organization. Genome Biol. 2015, 16, 227. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Torres, M.C.; Reese, J.T.; Childers, C.P.; Bennett, A.K.; Sundaram, J.P.; Childs, K.L.; Anzola, J.M.; Milshina, N.; Elsik, C.G. Hymenoptera genome database: Integrated community resources for insect species of the order hymenoptera. Nucleic Acids Res. 2010, 39, D658–D662. [Google Scholar] [CrossRef] [PubMed]
- Merkling, S.H.; van Rij, R.P. Analysis of resistance and tolerance to virus infection in drosophila. Nat. Protocols 2015, 10, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- Merkling, S.H.; van Rij, R.P. Beyond rnai: Antiviral defense strategies in drosophila and mosquito. J. Insect Physiol. 2013, 59, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Kingsolver, M.B.; Hardy, R.W. Making connections in insect innate immunity. Proc. Natl. Acad. Sci. USA 2012, 109, 18639–18640. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.A. The immune response of drosophila. Nature 2003, 426, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Imler, J.L.; Hoffmann, J.A. Toll receptors in innate immunity. Trends Cell Biol. 2001, 11, 304–311. [Google Scholar] [CrossRef]
- Lamiable, O.; Imler, J.-L. Induced antiviral innate immunity in drosophila. Curr. Opin. Microbiol. 2014, 20, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Antiviral immunity and virus-mediated antagonism in disease vector mosquitoes. Trends Microbiol. 2018, 26, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Blair, C.; Olson, K. The role of rna interference (rnai) in arbovirus-vector interactions. Viruses 2015, 7, 820–843. [Google Scholar] [CrossRef] [PubMed]
- Imler, J.-L.; Bulet, P. Antimicrobial peptides in drosophila: Structures, activities and gene regulation. In Mechanisms of Epithelial Defense; KARGER: Basel, Switzerland, 2005; Volume 86, pp. 1–21. [Google Scholar]
- Lourenco, A.P.; Florecki, M.M.; Simões, Z.L.P.; Evans, J.D. Silencing of Apis mellifera dorsal genes reveals their role in expression of the antimicrobial peptide defensin-1. Insect Mol. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Danihlík, J.; Aronstein, K.; Petřivalský, M. Antimicrobial peptides: A key component of honey bee innate immunity. J. Apicult. Res. 2016, 54, 123–136. [Google Scholar] [CrossRef]
- Schlüns, H.; Crozier, R.H. Relish regulates expression of antimicrobial peptide genes in the honeybee, Apis mellifera, shown by rna interference. Insect Mol. Biol. 2007, 16, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Riddell, C.; Adams, S.; Schmid-Hempel, P.; Mallon, E.B. Differential expression of immune defences is associated with specific host-parasite interactions in insects. PLoS ONE 2009, 4, e7621. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Shi, M.; Chen, X.-X. Antimicrobial peptide evolution in the asiatic honey bee apis cerana. PLoS ONE 2009, 4, e4239. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Choo, Y.M.; Lee, K.S.; Yoon, H.J.; Kim, I.; Je, Y.H.; Sohn, H.D.; Jin, B.R. Cloning and expression profiling of four antibacterial peptide genes from the bumblebee Bombus ignitus. Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 2008, 150, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Doublet, V.; Poeschl, Y.; Gogol-Döring, A.; Alaux, C.; Annoscia, D.; Aurori, C.; Barribeau, S.M.; Bedoya-Reina, O.C.; Brown, M.J.F.; Bull, J.C.; et al. Unity in defence: Honeybee workers exhibit conserved molecular responses to diverse pathogens. BMC Genom. 2017, 18, 207. [Google Scholar] [CrossRef] [PubMed]
- Zanni, V.; Galbraith, D.A.; Annoscia, D.; Grozinger, C.M.; Nazzi, F. Transcriptional signatures of parasitization and markers of colony decline in varroa-infested honey bees (Apis mellifera). Insect Biochem. Mol. Biol. 2017, 87, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Fannon, J.M.; Moore, J.D.; Wood, G.R.; Evans, D.J. The iflaviruses sacbrood virus and deformed wing virus evoke different transcriptional responses in the honeybee which may facilitate their horizontal or vertical transmission. PeerJ 2016, 4, e1591. [Google Scholar] [CrossRef] [PubMed]
- Steinmann, N.; Corona, M.; Neumann, P.; Dainat, B. Overwintering is associated with reduced expression of immune genes and higher susceptibility to virus infection in honey bees. PLoS ONE 2015, 10, e0129956. [Google Scholar] [CrossRef] [PubMed]
- Merkling, S.H.; Overheul, G.J.; van Mierlo, J.T.; Arends, D.; Gilissen, C.; van Rij, R.P. The heat shock response restricts virus infection in drosophila. Sci. Rep. 2015, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, T.; Takeuchi, A.; Siomi, H.; Siomi, M.C. A direct role for hsp90 in pre-risc formation in drosophila. Nat. Struct. Mol. Biol. 2010, 17, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Sim, C.; Hong, Y.S.; Tsetsarkin, K.A.; Vanlandingham, D.L.; Higgs, S.; Collins, F.H. Anopheles gambiae heat shock protein cognate 70b impedes o’nyong-nyong virus replication. BMC Genom. 2007, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Mockel, N.; Gisder, S.; Genersch, E. Horizontal transmission of deformed wing virus: Pathological consequences in adult bees (Apis mellifera) depend on the transmission route. J. Gen. Virol. 2011, 92, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Bowen-Walker, P.L.; Martin, S.J.; Gunn, A. The transmission of deformed wing virus between honeybees (Apis mellifera L.) by the ectoparasitic mite Varroa jacobsoni oud. J. Invertebr. Pathol. 1999, 73, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Nazzi, F.; Pennacchio, F. Honey bee antiviral immune barriers as affected by multiple stress factors: A novel paradigm to interpret colony health decline and collapse. Viruses 2018, 10, 159. [Google Scholar] [CrossRef] [PubMed]
- Kuster, R.D.; Boncristiani, H.F.; Rueppell, O. Immunogene and viral transcript dynamics during parasitic Varroa destructor mite infection of developing honey bee (Apis mellifera) pupae. J. Exp. Biol. 2014, 217, 1710–1718. [Google Scholar] [CrossRef] [PubMed]
- Chejanovsky, N.; Ophir, R.; Schwager, M.S.; Slabezki, Y.; Grossman, S.; Cox-Foster, D. Characterization of viral sirna populations in honey bee colony collapse disorder. Virology 2014, 454, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.D.; Eu, Y.J.; Whyard, S.; Currie, R.W. Reduction in deformed wing virus infection in larval and adult honey bees (Apis mellifera L.) by double-stranded rna ingestion. Insect Mol. Biol. 2012, 21, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Won, S.; Chtarbanova, S.; Squiban, B.; Ocorr, K.; Bodmer, R.; Beutler, B.; Hoffmann, J.A.; Imler, J.L. Atp-sensitive potassium channel (katp)-dependent regulation of cardiotropic viral infections. Proc. Natl. Acad. Sci. USA 2011, 108, 12024–12029. [Google Scholar] [CrossRef] [PubMed]
- Palmer, W.H.; Varghese, F.S.; van Rij, R.P. Natural variation in resistance to virus infection in dipteran insects. Viruses 2018, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Rückert, C.; Bell-Sakyi, L.; Fazakerley, J.K.; Fragkoudis, R. Antiviral responses of arthropod vectors: An update on recent advances. Virusdisease 2014, 25, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vargas, I.; Travanty, E.A.; Keene, K.M.; Franz, A.W.E.; Beaty, B.J.; Blair, C.D.; Olson, K.E. Rna interference, arthropod-borne viruses, and mosquitoes. Virus Res. 2004, 102, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.-W. RNA-based antiviral immunity. Nat. Rev. Immunol. 2010, 10, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Fire, A. RNA-triggered gene silencing. Trends Genet. 1999, 15, 358–363. [Google Scholar] [CrossRef]
- Flenniken, M.L.; Kunitomi, M.; Tassetto, M.; Andino, R. The antiviral role of rna interference. In Insect Virology; Asgari, S., Johnson, K.N., Eds.; Caister Academic Press: Norfolk, UK, 2010; pp. 367–388. [Google Scholar]
- Hammond, S.M. Argonaute2, a link between genetic and biochemical analyses of rnai. Science 2001, 293, 1146–1150. [Google Scholar] [CrossRef] [PubMed]
- Blair, C.D. Mosquito rnai is the major innate immune pathway controlling arbovirus infection and transmission. Future Microbiol. 2011, 6, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Bronkhorst, A.W.; van Rij, R.P. The long and short of antiviral defense: Small RNA-based immunity in insects. Curr. Opin. Virol. 2014, 7, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Brutscher, L.M.; Flenniken, M.L. RNAi and antiviral defense in the honey bee. J. Immunol. Res. 2015, 2015, 941897. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, C.; Dishongh, R.; Moore, S.C.; Whitt, M.A.; Chow, M.; Machaca, K. Rna interference is an antiviral defence mechanism in Caenorhabditis elegans. Nature 2005, 436, 1044–1047. [Google Scholar] [CrossRef] [PubMed]
- Zamore, P.D. Ancient pathways programmed by small rnas. Science 2002, 296, 1265–1269. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded rna in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. Rna silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ye, X.; Jiang, F.; Liang, C.; Chen, D.; Peng, J.; Kinch, L.N.; Grishin, N.V.; Liu, Q. C3po, an endoribonuclease that promotes rnai by facilitating risc activation. Science 2009, 325, 750–753. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Induction and suppression of rna silencing by an animal virus. Science 2002, 296, 1319–1321. [Google Scholar] [CrossRef] [PubMed]
- Van Rij, R.P.; Saleh, M.C.; Berry, B.; Foo, C.; Houk, A.; Antoniewski, C.; Andino, R. The rna silencing endonuclease argonaute 2 mediates specific antiviral immunity in Drosophila melanogaster. Genes Dev. 2006, 20, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.-C.; van Rij, R.P.; Hekele, A.; Gillis, A.; Foley, E.; O’Farrell, P.H.; Andino, R. The endocytic pathway mediates cell entry of dsrna to induce rnai silencing. Nat. Cell Biol. 2006, 8, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.-C.; Tassetto, M.; van Rij, R.P.; Goic, B.; Gausson, V.; Berry, B.; Jacquier, C.; Antoniewski, C.; Andino, R. Antiviral immunity in drosophila requires systemic RNA interference spread. Nature 2009, 458, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Goic, B.; Vodovar, N.; Mondotte, J.A.; Monot, C.; Frangeul, L.; Blanc, H.; Gausson, V.; Vera-Otarola, J.; Cristofari, G.; Saleh, M.-C. Rna-mediated interference and reverse transcription control the persistence of rna viruses in the insect model drosophila. Nat. Immunol. 2013, 14, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Tassetto, M.; Kunitomi, M.; Andino, R. Circulating immune cells mediate a systemic rnai-based adaptive antiviral response in drosophila. Cell 2017, 169, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Poirier, E.Z.; Goic, B.; Tome-Poderti, L.; Frangeul, L.; Boussier, J.; Gausson, V.; Blanc, H.; Vallet, T.; Loyd, H.; Levi, L.I.; et al. Dicer-2-dependent generation of viral DNA from defective genomes of rna viruses modulates antiviral immunity in insects. Cell Host Microbe 2018, 23, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Y.; Yan, X.; Han, R. Prevention of chinese sacbrood virus infection in Apis cerana using RNA interference. Curr. Microbiol. 2010, 61, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Paldi, N.; Shafir, S.; Kalev, H.; Tsur, E.; Glick, E.; Sela, I. Iapv, a bee-affecting virus associated with colony collapse disorder can be silenced by dsrna ingestion. Insect Mol. Biol. 2009, 18, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Nunes, F.; Aleixo, A.; Barchuk, A.; Bomtorin, A.; Grozinger, C.; Simões, Z. Non-target effects of green fluorescent protein (gfp)-derived double-stranded rna (dsrna-gfp) used in honey bee rna interference (RNAi) assays. Insects 2013, 4, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, J.G.; Duan, J.J. Rnai-based insecticidal crops: Potential effects on nontarget species. Bioscience 2013, 63, 657–665. [Google Scholar] [CrossRef]
- Cappelle, K.; Smagghe, G.; Dhaenens, M.; Meeus, I. Israeli acute paralysis virus infection leads to an enhanced rna interference response and not its suppression in the bumblebee Bombus terrestris. Viruses 2016, 8, 334. [Google Scholar] [CrossRef] [PubMed]
- Jarosch, A.; Moritz, R.F.A. Rna interference in honeybees: Off-target effects caused by dsrna. Apidologie 2012, 43, 128–138. [Google Scholar] [CrossRef]
- Hunter, W.; Ellis, J.; vanEngelsdorp, D.; Hayes, J.; Westervelt, D.; Glick, E.; Williams, M.; Sela, I.; Maori, E.; Pettis, J.; et al. Large-scale field application of rnai technology reducing israeli acute paralysis virus disease in honey bees (Apis mellifera, hymenoptera: Apidae). PLoS Pathog. 2010, 6, e1001160. [Google Scholar] [CrossRef] [PubMed]
- Beye, M.; Hartel, S.; Hagen, A.; Hasselmann, M.; Omholt, S.W. Specific developmental gene silencing in the honey bee using a homeobox motif. Insect Mol. Biol. 2002, 11, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Aronstein, K.; Saldivar, E. Characterization of a honey bee toll related receptor gene am18w and its potential involvement in antimicrobial immune defense. Apidologie 2005, 36, 3–14. [Google Scholar] [CrossRef]
- Patel, A.; Fondrk, M.K.; Kaftanoglu, O.; Emore, C.; Hunt, G.; Frederick, K.; Amdam, G.V. The making of a queen: Tor pathway is a key player in diphenic caste development. PLoS ONE 2007, 2, e509. [Google Scholar] [CrossRef] [PubMed]
- Barchuk, A.R.; Figueiredo, V.L.C.; Simões, Z.L.P. Downregulation of ultraspiracle gene expression delays pupal development in honeybees. J. Insect Physiol. 2008, 54, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, K.A.; Ihle, K.E.; Frederick, K.; Fondrk, M.K.; Smedal, B.; Hartfelder, K.; Amdam, G.V. Insulin-like peptide genes in honey bee fat body respond differently to manipulation of social behavioral physiology. J. Exp. Biol. 2011, 214, 1488–1497. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Baker, N.; Amdam, G.V. Rnai-mediated double gene knockdown and gustatory perception measurement in honey bees (Apis mellifera). J. Vis. Exp. 2013. [Google Scholar] [CrossRef] [PubMed]
- Ihle, K.E.; Fondrk, M.K.; Page, R.E.; Amdam, G.V. Genotype effect on lifespan following vitellogenin knockdown. Exp. Gerontol. 2015, 61, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.M.; Ihle, K.E.; Fondrk, M.K.; Page, R.E.; Amdam, G.V. The gene vitellogenin has multiple coordinating effects on social organization. PLoS Biol. 2007, 5, e62. [Google Scholar] [CrossRef] [PubMed]
- Deddouche, S.; Matt, N.; Budd, A.; Mueller, S.; Kemp, C.; Galiana-Arnoux, D.; Dostert, C.; Antoniewski, C.; Hoffmann, J.A.; Imler, J.-L. The dexd/h-box helicase dicer-2 mediates the induction of antiviral activity in drosophila. Nat. Immunol. 2008, 9, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Paradkar, P.N.; Trinidad, L.; Voysey, R.; Duchemin, J.B.; Walker, P.J. Secreted vago restricts west nile virus infection in culex mosquito cells by activating the Jak-stat pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 18915–18920. [Google Scholar] [CrossRef] [PubMed]
- Paradkar, P.N.; Duchemin, J.B.; Voysey, R.; Walker, P.J. Dicer-2-dependent activation of culex vago occurs via the traf-rel2 signaling pathway. PLoS Negl. Trop. Dis. 2014, 8, e2823. [Google Scholar] [CrossRef] [PubMed]
- Pichlmair, A.; Schulz, O.; Tan, C.P.; Rehwinkel, J.; Kato, H.; Takeuchi, O.; Akira, S.; Way, M.; Schiavo, G.; Reis e Sousa, C. Activation of mda5 requires higher-order rna structures generated during virus infection. J. Virol. 2009, 83, 10761–10769. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Rig-i-like antiviral protein in flies. Nat. Immunol. 2008, 9, 1327–1328. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The rna helicase rig-i has an essential function in double-stranded rna-induced innate antiviral responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Kingsolver, M.B.; Huang, Z.; Hardy, R.W. Insect antiviral innate immunity: Pathways, effectors, and connections. J. Mol. Biol. 2013, 425, 4921–4936. [Google Scholar] [CrossRef] [PubMed]
- Cardinal, S.; Straka, J.; Danforth, B.N. Comprehensive phylogeny of apid bees reveals the evolutionary origins and antiquity of cleptoparasitism. Proc. Natl. Acad. Sci. USA 2010, 107, 16207–16211. [Google Scholar] [CrossRef] [PubMed]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.-L.; Barthel, A.; et al. Rna interference in lepidoptera: An overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Boisson, B.; Jacques, J.C.; Choumet, V.; Martin, E.; Xu, J.; Vernick, K.; Bourgouin, C. Gene silencing in mosquito salivary glands by rnai. FEBS Lett. 2006, 580, 1988–1992. [Google Scholar] [CrossRef] [PubMed]
- Nayak, A.; Berry, B.; Tassetto, M.; Kunitomi, M.; Acevedo, A.; Deng, C.; Krutchinsky, A.; Gross, J.; Antoniewski, C.; Andino, R. Cricket paralysis virus antagonizes argonaute 2 to modulate antiviral defense in drosophila. Nat. Struct. Mol. Biol. 2010, 17, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Smagghe, G.; Meeus, I. The role of a single gene encoding the single von willebrand factor c-domain protein (svc) in bumblebee immunity extends beyond antiviral defense. Insect Biochem. Mol. Biol. 2017, 91, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Ronald, P.C.; Beutler, B. Plant and animal sensors of conserved microbial signatures. Science 2010, 330, 1061–1064. [Google Scholar] [CrossRef] [PubMed]
- Coffman, S.R.; Lu, J.; Guo, X.; Zhong, J.; Jiang, H.; Broitman-Maduro, G.; Li, W.-X.; Lu, R.; Maduro, M.; Ding, S.-W. Caenorhabditis elegans rig-i homolog mediates antiviral RNA interference downstream of dicer-dependent biogenesis of viral small interfering rnas. mBio 2017, 8, e00264. [Google Scholar] [CrossRef] [PubMed]
- Ashe, A.; Sarkies, P.; Le Pen, J.; Tanguy, M.; Miska, E.A. Antiviral rna interference against orsay virus is neither systemic nor transgenerational in Caenorhabditis elegans. J. Virol. 2015, 89, 12035–12046. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Maduro, M.; Li, F.; Li, H.W.; Broitman-Maduro, G.; Li, W.X.; Ding, S.W. Animal virus replication and RNAi-mediated antiviral silencing in Caenorhabditis elegans. Nature 2005, 436, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Schnettler, E.; Tykalová, H.; Watson, M.; Sharma, M.; Sterken, M.G.; Obbard, D.J.; Lewis, S.H.; McFarlane, M.; Bell-Sakyi, L.; Barry, G.; et al. Induction and suppression of tick cell antiviral rnai responses by tick-borne flaviviruses. Nucleic Acids Res. 2014, 42, 9436–9446. [Google Scholar] [CrossRef] [PubMed]
- Green, T.J.; Speck, P. Antiviral defense and innate immune memory in the oyster. Viruses 2018, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- Robalino, J.; Bartlett, T.C.; Chapman, R.W.; Gross, P.S.; Browdy, C.L.; Warr, G.W. Double-stranded RNA and antiviral immunity in marine shrimp: Inducible host mechanisms and evidence for the evolution of viral counter-responses. Dev. Comp. Immunol. 2007, 31, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Pitaluga, A.N.; Mason, P.W.; Traub-Cseko, Y.M. Non-specific antiviral response detected in RNA-treated cultured cells of the sandfly, Lutzomyia longipalpis. Dev. Comp. Immunol. 2008, 32, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Martins-da-Silva, A.; Telleria, E.; Batista, M.; Marchini, F.; Traub-Csekö, Y.; Tempone, A. Identification of secreted proteins involved in nonspecific dsrna-mediated Lutzomyia longipalpis ll5 cell antiviral response. Viruses 2018, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Van Mierlo, J.T.; van Cleef, K.W.R.; van Rij, R.P. Defense and counterdefense in the RNAi-based antiviral immune system in insects. In Antiviral RNAi. Methods in Molecular Biology (Methods and Protocols); van Rij, R., Ed.; Humana Press: New York, NY, USA, 2011; Volume 721, pp. 3–22. [Google Scholar]
- De Maio, F.A.; Risso, G.; Iglesias, N.G.; Shah, P.; Pozzi, B.; Gebhard, L.G.; Mammi, P.; Mancini, E.; Yanovsky, M.J.; Andino, R.; et al. The dengue virus ns5 protein intrudes in the cellular spliceosome and modulates splicing. PLoS Pathog. 2016, 12, e1005841. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jorda, M.; Jones, P.L.; Maleszka, R.; Ling, X.; Robertson, H.M.; Mizzen, C.A.; Peinado, M.A.; Robinson, G.E. Functional cpg methylation system in a social insect. Science 2006, 314, 645–647. [Google Scholar] [CrossRef] [PubMed]
- Tidbury, H.J.; Pedersen, A.B.; Boots, M. Within and transgenerational immune priming in an insect to a DNA virus. Proc. Biol. Sci. 2011, 278, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Gourbal, B.; Pinaud, S.; Beckers, G.J.M.; Van Der Meer, J.W.M.; Conrath, U.; Netea, M.G. Innate immune memory: An evolutionary perspective. Immunol. Rev. 2018, 283, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Tanne, E.; Sela, I. Reciprocal sequence exchange between non-retro viruses and hosts leading to the appearance of new host phenotypes. Virology 2007, 362, 342–349. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).