The C-Type Lectin Domain Gene Family in Aedes aegypti and Their Role in Arbovirus Infection



Abstract
1. Introduction
2. Materials and Methods
2.1. C-Type Lectin Identification
2.2. Structural Annotation of A. aegypti CTLD Encoding Genes
2.3. Phylogenetic Analysis
2.4. Gene Expression Analysis
3. Results
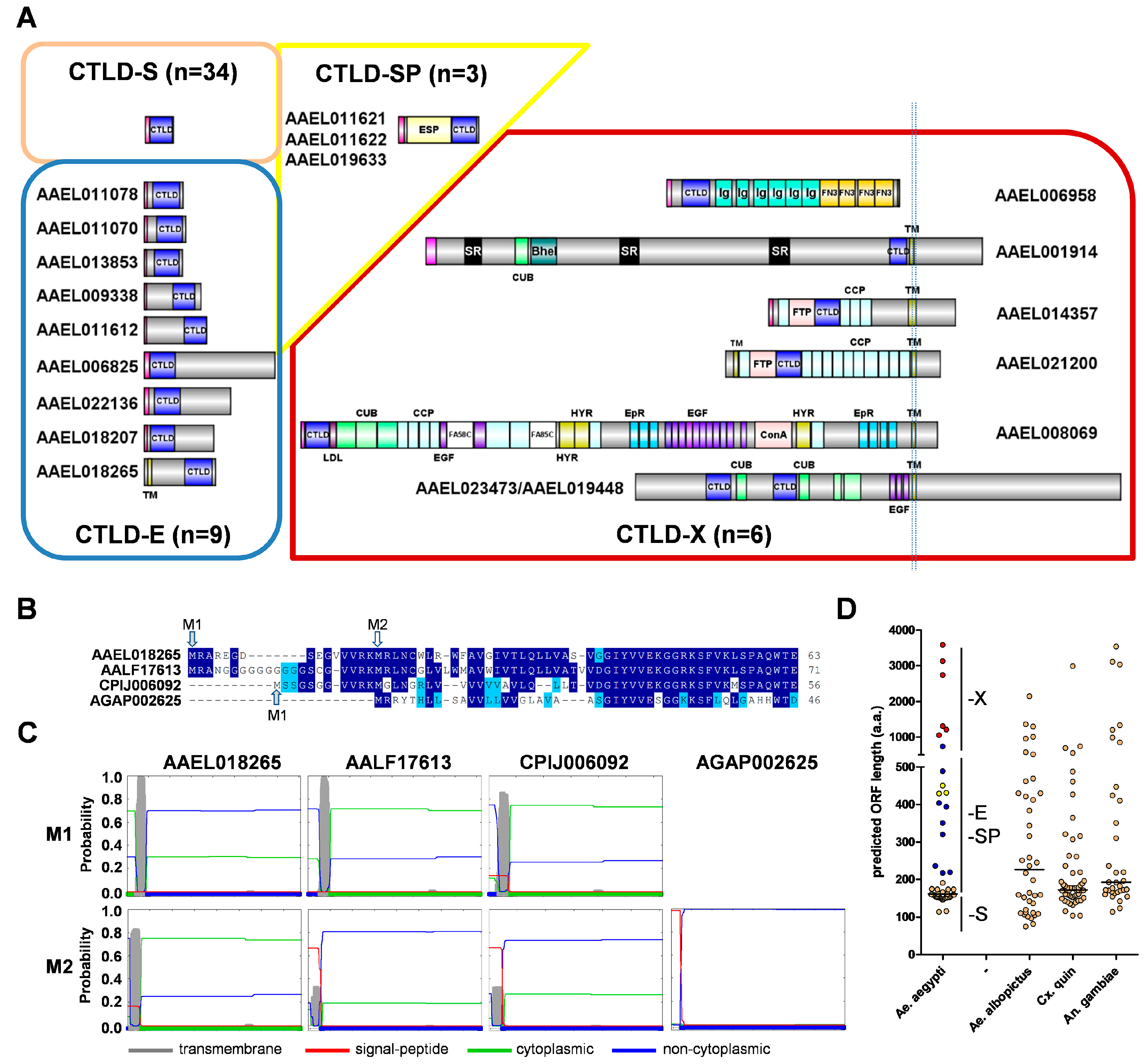
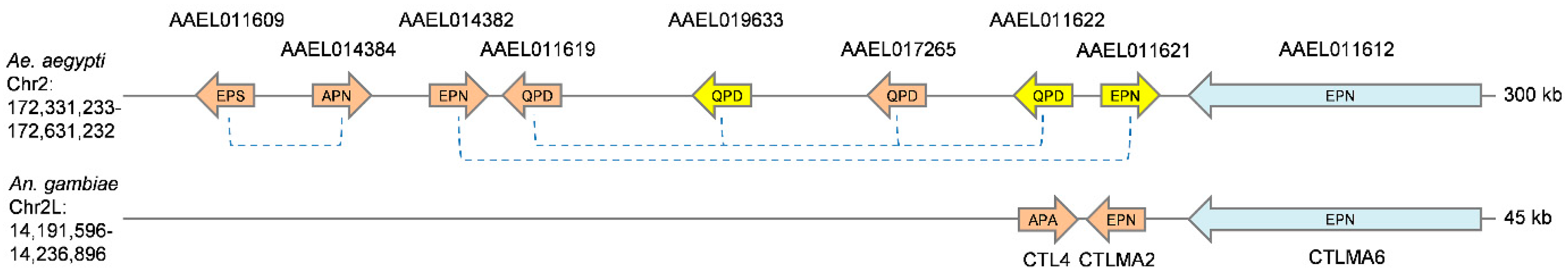
3.1. Identification and Classification of A. aegypti CTLDcps
3.2. Developmental Expression Profile of A. aegypti CTLDcps
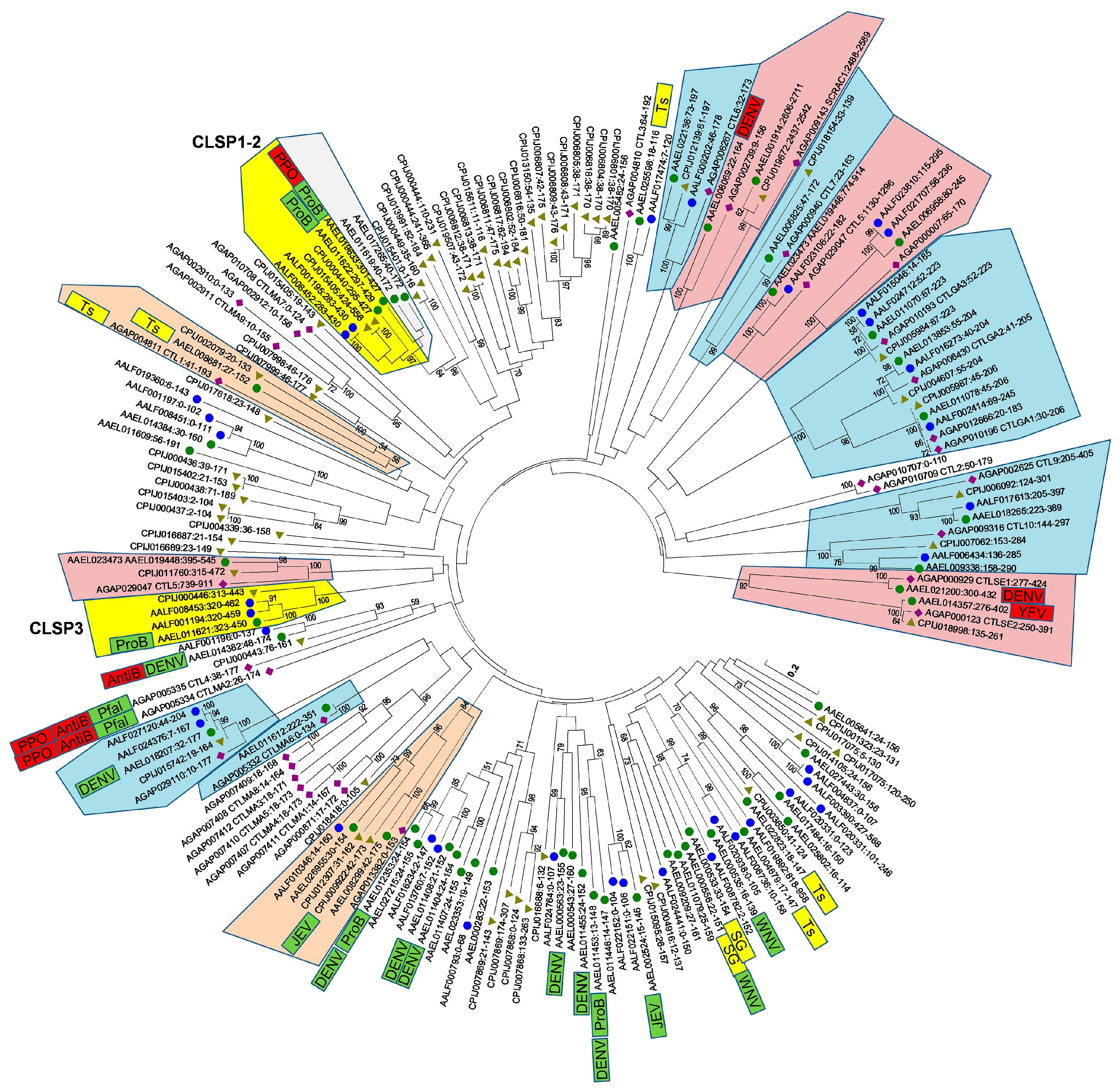
3.3. Phylogeny of Mosquito CTLDs
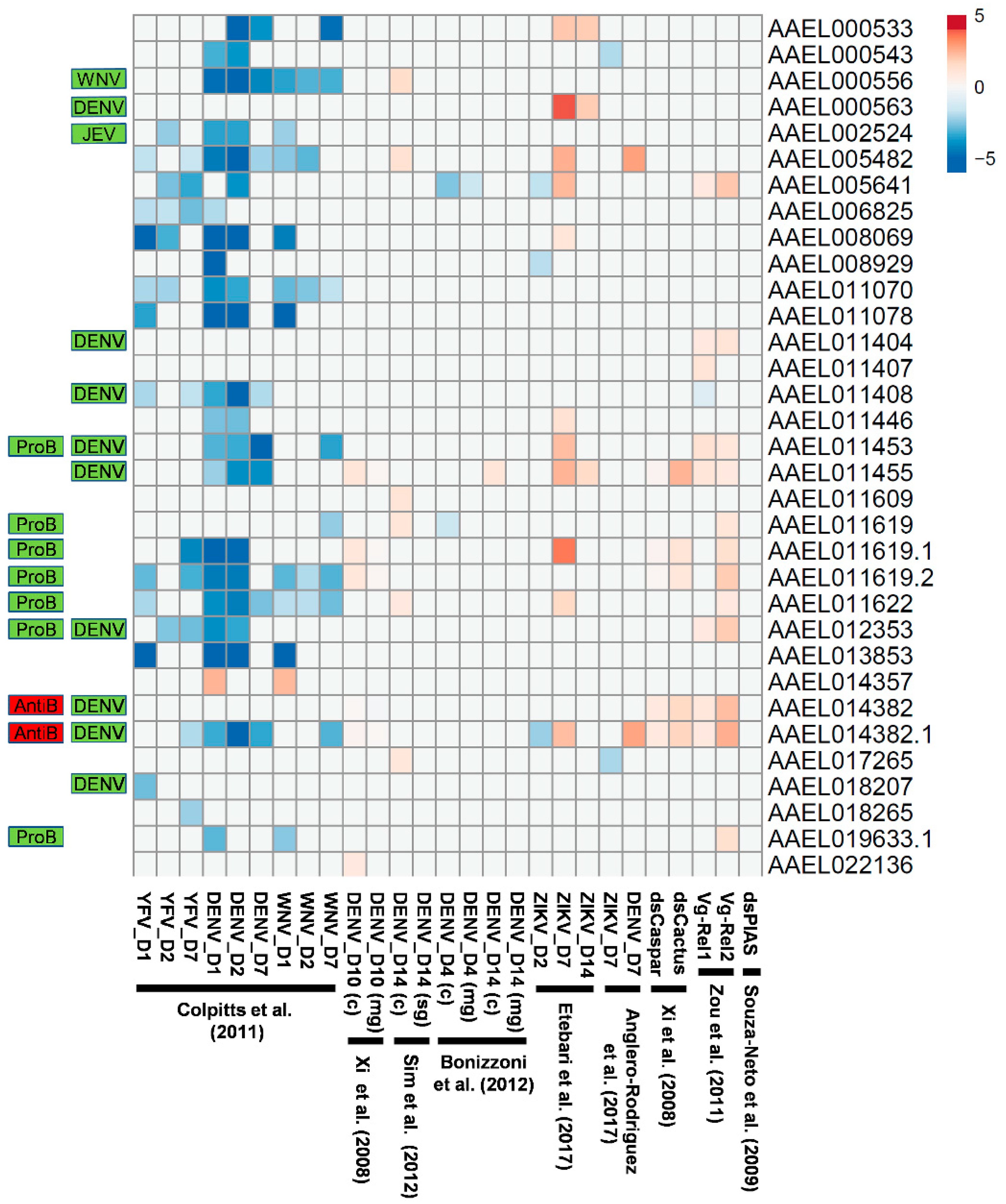
3.4. Virus-Induced Expression of Transcripts Encoding CTLDcps
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cheng, G.; Cox, J.; Wang, P.; Krishnan, M.N.; Dai, J.; Qian, F.; Anderson, J.F.; Fikrig, E. A C-type lectin collaborates with a CD45 phosphatase homolog to facilitate west nile virus infection of mosquitoes. Cell 2010, 142, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Qian, Y.; Jung, Y.S.; Zhou, B.; Cao, R.; Shen, T.; Shao, D.; Wei, J.; Ma, Z.; Chen, P.; et al. mosGCTL-7, a C-type lectin protein, mediates Japanese encephalitis virus infection in mosquitoes. J. Virol. 2017, 91, e01348-16. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, F.; Liu, J.; Xiao, X.; Zhang, S.; Qin, C.; Xiang, Y.; Wang, P.; Cheng, G. Transmission-blocking antibodies against mosquito C-type lectins for dengue prevention. PLoS Pathog. 2014, 10, e1003931. [Google Scholar] [CrossRef] [PubMed]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, R.M.; Kriventseva, E.V.; Meister, S.; Xi, Z.; Alvarez, K.S.; Bartholomay, L.C.; Barillas-Mury, C.; Bian, G.; Blandin, S.; Christensen, B.M.; et al. Evolutionary dynamics of immune-related genes and pathways in disease-vector mosquitoes. Science 2007, 316, 1738–1743. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Xiao, X.; Liu, Y.; Zhang, R.; Liu, J.; Liu, Q.; Wang, P.; Cheng, G. Mosquito C-type lectins maintain gut microbiome homeostasis. Nat. Microbiol. 2016, 1, 16023. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.W.; Zou, Z.; Raikhel, A.S. A new factor in the Aedes aegypti immune response: ClSP2 modulates melanization. EMBO Rep. 2011, 12, 938–943. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.J.; Cao, X.; He, Y.; Hu, Y.; Zhang, X.; Chen, Y.R.; Blissard, G.; Kanost, M.R.; Yu, X.Q.; Jiang, H. Structural features, evolutionary relationships, and transcriptional regulation of C-type lectin-domain proteins in manduca sexta. Insect Biochem. Mol. Biol. 2015, 62, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.J.; Shahzad, T.; Liu, S.; Wu, P.; He, Y.T.; Sun, W.J.; Fan, X.Y.; Yang, Y.F.; Shi, Q.; Yu, X.Q. Identification of C-type lectin-domain proteins (CTLDPs) in silkworm bombyx mori. Dev. Comp. Immunol. 2015, 53, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Calderon, G.I.; Emrich, S.J.; MacCallum, R.M.; Maslen, G.; Dialynas, E.; Topalis, P.; Ho, N.; Gesing, S.; VectorBase, C.; Madey, G.; et al. Vectorbase: An updated bioinformatics resource for invertebrate vectors and other organisms related with human diseases. Nucleic Acids Res. 2015, 43, D707–D713. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Bryant, S.H. CD-Search: Protein domain annotations on the fly. Nucleic Acids Res. 2004, 32, W327–W331. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.R.; Hall, I.M. Bedtools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Helt, G.A.; Reese, J.T.; Munoz-Torres, M.C.; Childers, C.P.; Buels, R.M.; Stein, L.; Holmes, I.H.; Elsik, C.G.; Lewis, S.E. Web Apollo: A web-based genomic annotation editing platform. Genome Biol. 2013, 14, R93. [Google Scholar] [CrossRef] [PubMed]
- Kall, L.; Krogh, A.; Sonnhammer, E.L. Advantages of combined transmembrane topology and signal peptide prediction–The phobius web server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xie, Y.; Ma, J.; Luo, X.; Nie, P.; Zuo, Z.; Lahrmann, U.; Zhao, Q.; Zheng, Y.; Zhao, Y.; et al. IBS: An illustrator for the presentation and visualization of biological sequences. Bioinformatics 2015, 31, 3359–3361. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Akbari, O.S.; Antoshechkin, I.; Amrhein, H.; Williams, B.; Diloreto, R.; Sandler, J.; Hay, B.A. The developmental transcriptome of the mosquito Aedes aegypti, an invasive species and major arbovirus vector. G3 Genes Genomes Genet. 2013, 3, 1493–1509. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. Featurecounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Ramirez, J.L.; Dimopoulos, G. Dengue virus infection of the Aedes aegypti salivary gland and chemosensory apparatus induces genes that modulate infection and blood-feeding behavior. PLoS Pathog. 2012, 8, e1002631. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Souza-Neto, J.; Xi, Z.; Kokoza, V.; Shin, S.W.; Dimopoulos, G.; Raikhel, A. Transcriptome analysis of Aedes aegypti transgenic mosquitoes with altered immunity. PLoS Pathog. 2011, 7, e1002394. [Google Scholar] [CrossRef] [PubMed]
- Colpitts, T.M.; Cox, J.; Vanlandingham, D.L.; Feitosa, F.M.; Cheng, G.; Kurscheid, S.; Wang, P.; Krishnan, M.N.; Higgs, S.; Fikrig, E. Alterations in the Aedes aegypti transcriptome during infection with West Nile, dengue and yellow fever viruses. PLoS Pathog. 2011, 7, e1002189. [Google Scholar] [CrossRef] [PubMed]
- Anglero-Rodriguez, Y.I.; Talyuli, O.A.; Blumberg, B.J.; Kang, S.; Demby, C.; Shields, A.; Carlson, J.; Jupatanakul, N.; Dimopoulos, G. An Aedes aegypti-associated fungus increases susceptibility to dengue virus by modulating gut trypsin activity. Elife 2017, 6, e28844. [Google Scholar] [CrossRef] [PubMed]
- Etebari, K.; Hegde, S.; Saldana, M.A.; Widen, S.G.; Wood, T.G.; Asgari, S.; Hughes, G.L. Global transcriptome analysis of Aedes aegypti mosquitoes in response to Zika virus infection. mSphere 2017, 2, e00456-17. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M.; Martin-Martin, I.; Arca, B.; Calvo, E. A deep insight into the sialome of male and female Aedes aegypti mosquitoes. PLoS ONE 2016, 11, e0151400. [Google Scholar] [CrossRef] [PubMed]
- Salamov, A.A.; Nishikawa, T.; Swindells, M.B. Assessing protein coding region integrity in cDNA sequencing projects. Bioinformatics 1998, 14, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Liu, Y.; Zhang, X.; Wang, J.; Li, Z.; Pang, X.; Wang, P.; Cheng, G. Complement-related proteins control the flavivirus infection of Aedes aegypti by inducing antimicrobial peptides. PLoS Pathog. 2014, 10, e1004027. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.G.; Jiang, X.; Gu, J.; Xu, M.; Wu, Y.; Deng, Y.; Zhang, C.; Bonizzoni, M.; Dermauw, W.; Vontas, J.; et al. Genome sequence of the Asian Tiger mosquito, Aedes albopictus, reveals insights into its biology, genetics, and evolution. Proc. Natl. Acad. Sci. USA 2015, 112, E5907–E5915. [Google Scholar] [CrossRef] [PubMed]
- Arensburger, P.; Megy, K.; Waterhouse, R.M.; Abrudan, J.; Amedeo, P.; Antelo, B.; Bartholomay, L.; Bidwell, S.; Caler, E.; Camara, F.; et al. Sequencing of Culex quinquefasciatus establishes a platform for mosquito comparative genomics. Science 2010, 330, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.A.; Subramanian, G.M.; Halpern, A.; Sutton, G.G.; Charlab, R.; Nusskern, D.R.; Wincker, P.; Clark, A.G.; Ribeiro, J.M.; Wides, R.; et al. The genome sequence of the malaria mosquito Anopheles gambiae. Science 2002, 298, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Whiten, S.R.; Ray, W.K.; Helm, R.F.; Adelman, Z.N. Characterization of the adult Aedes aegypti early midgut peritrophic matrix proteome using LC-MS. PLoS ONE 2018, 13, e0194734. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.A.; Nolan, T.; Fischer, B.; Pinder, A.; Crisanti, A.; Russell, S. A comprehensive gene expression atlas of sex- and tissue-specificity in the malaria vector, Anopheles gambiae. BMC Genom. 2011, 12, 296. [Google Scholar] [CrossRef] [PubMed]
- Osta, M.A.; Christophides, G.K.; Kafatos, F.C. Effects of mosquito genes on Plasmodium development. Science 2004, 303, 2030–2032. [Google Scholar] [CrossRef] [PubMed]
- Schnitger, A.K.; Yassine, H.; Kafatos, F.C.; Osta, M.A. Two C-type lectins cooperate to defend Anopheles gambiae against gram-negative bacteria. J. Biol. Chem. 2009, 284, 17616–17624. [Google Scholar] [CrossRef] [PubMed]
- Souza-Neto, J.A.; Sim, S.; Dimopoulos, G. An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc. Natl. Acad. Sci. USA 2009, 106, 17841–17846. [Google Scholar] [CrossRef] [PubMed]
- Bonizzoni, M.; Dunn, W.A.; Campbell, C.L.; Olson, K.E.; Marinotti, O.; James, A.A. Complex modulation of the Aedes aegypti transcriptome in response to dengue virus infection. PLoS ONE 2012, 7, e50512. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Souza-Neto, J.; Torres Cosme, R.; Rovira, J.; Ortiz, A.; Pascale, J.M.; Dimopoulos, G. Reciprocal tripartite interactions between the aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl. Trop. Dis. 2012, 6, e1561. [Google Scholar] [CrossRef] [PubMed]
- Avirutnan, P.; Hauhart, R.E.; Marovich, M.A.; Garred, P.; Atkinson, J.P.; Diamond, M.S. Complement-mediated neutralization of dengue virus requires mannose-binding lectin. mBio 2011, 2, e00276-11. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.L.; de Wet, B.J.; Martinez-Pomares, L.; Radcliffe, C.M.; Dwek, R.A.; Rudd, P.M.; Gordon, S. The mannose receptor mediates dengue virus infection of macrophages. PLoS Pathog. 2008, 4, e17. [Google Scholar] [CrossRef]
- Tassaneetrithep, B.; Burgess, T.H.; Granelli-Piperno, A.; Trumpfheller, C.; Finke, J.; Sun, W.; Eller, M.A.; Pattanapanyasat, K.; Sarasombath, S.; Birx, D.L.; et al. DC-SIGN (CD209) mediates dengue virus infection of human dendritic cells. J. Exp. Med. 2003, 197, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Blitvich, B.J.; Firth, A.E. Insect-specific flaviviruses: A systematic review of their discovery, host range, mode of transmission, superinfection exclusion potential and genomic organization. Viruses 2015, 7, 1927–1959. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Hu, Y.; Xing, L.S.; Jiang, H.; Hu, S.N.; Raikhel, A.S.; Zou, Z. A critical role for CLSP2 in the modulation of antifungal immune response in mosquitoes. PLoS Pathog. 2015, 11, e1004931. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CLSP-1 (Signal P and SP Domain Only) | |||||
| CTLD-SP | Gene ID | Length | Evalue | Score | % Identity |
| CLSP-1 | AAEL011622-PA | 270 | 0 | 1442 | 100% |
| CLSP-2 | AAEL019633-PA | 270 | 5.00E-180 | 1305 | 90% |
| CLSP-3 | AAEL011621-PA | 253 | 9.00E-64 | 533 | 38.90% |
| AAEL019963-PA | 262 | 2.00E-55 | 471 | 34.20% | |
| AAEL005748-PA | 244 | 1.00E-52 | 454 | 37.60% | |
| AAEL018109-PB | 245 | 8.00E-51 | 450 | 37.90% | |
| AAEL002276-PA | 241 | 7.00E-51 | 436 | 36.10% | |
| AAEL001077-PA | 245 | 5.00E-48 | 432 | 38.70% | |
| AAEL010773-PA | 246 | 7.00E-48 | 431 | 36.40% | |
| AAEL001084-PA | 245 | 2.00E-47 | 427 | 38.30% | |
| CLSP-2 (Signal P and SP Domain Only) | |||||
| CLSP-2 | AAEL019633-PA | 270 | 0 | 1441 | 100% |
| CLSP-1 | AAEL011622-PA | 270 | 4.00E-180 | 1306 | 90% |
| CLSP-3 | AAEL011621-PA | 262 | 2.00E-65 | 544 | 37.70% |
| AAEL019963-PA | 262 | 2.00E-52 | 451 | 34.60% | |
| AAEL018109-PB | 245 | 8.00E-50 | 443 | 37.90% | |
| AAEL010773-PA | 246 | 1.00E-48 | 436 | 35.50% | |
| AAEL001084-PA | 245 | 1.00E-47 | 429 | 37.90% | |
| AAEL005748-PA | 244 | 1.00E-48 | 426 | 35.90% | |
| AAEL001077-PA | 245 | 9.00E-46 | 415 | 38.20% | |
| AAEL006253-PC | 243 | 2.00E-45 | 412 | 35.10% | |
| CLSP-3 (Signal P and SP Domain Only) | |||||
| CLSP-3 | AAEL011621-PA | 270 | 0 | 1435 | 100% |
| AAEL011375-PB | 234 | 8.00E-67 | 559 | 44.90% | |
| AAEL010773-PA | 233 | 4.00E-62 | 534 | 43.20% | |
| AAEL018109-PB | 259 | 2.00E-58 | 505 | 37.30% | |
| AAEL002880-PA | 266 | 1.00E-57 | 499 | 37.50% | |
| AAEL002254-PA | 263 | 5.00E-57 | 498 | 39.10% | |
| AAEL019963-PA | 237 | 4.00E-59 | 497 | 39.20% | |
| CLSP-2 | AAEL019633-PA | 257 | 1.00E-58 | 497 | 36.60% |
| AAEL002276-PA | 242 | 1.00E-59 | 496 | 41.10% | |
| CLSP-1 | AAEL011622-PA | 238 | 1.00E-57 | 490 | 38.30% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adelman, Z.N.; Myles, K.M. The C-Type Lectin Domain Gene Family in Aedes aegypti and Their Role in Arbovirus Infection. Viruses 2018, 10, 367. https://doi.org/10.3390/v10070367
Adelman ZN, Myles KM. The C-Type Lectin Domain Gene Family in Aedes aegypti and Their Role in Arbovirus Infection. Viruses. 2018; 10(7):367. https://doi.org/10.3390/v10070367
Chicago/Turabian StyleAdelman, Zach N., and Kevin M. Myles. 2018. "The C-Type Lectin Domain Gene Family in Aedes aegypti and Their Role in Arbovirus Infection" Viruses 10, no. 7: 367. https://doi.org/10.3390/v10070367
APA StyleAdelman, Z. N., & Myles, K. M. (2018). The C-Type Lectin Domain Gene Family in Aedes aegypti and Their Role in Arbovirus Infection. Viruses, 10(7), 367. https://doi.org/10.3390/v10070367