A Review of the Ongoing Research on Zika Virus Treatment


Abstract
1. Introduction
2. The ZIKA Virus
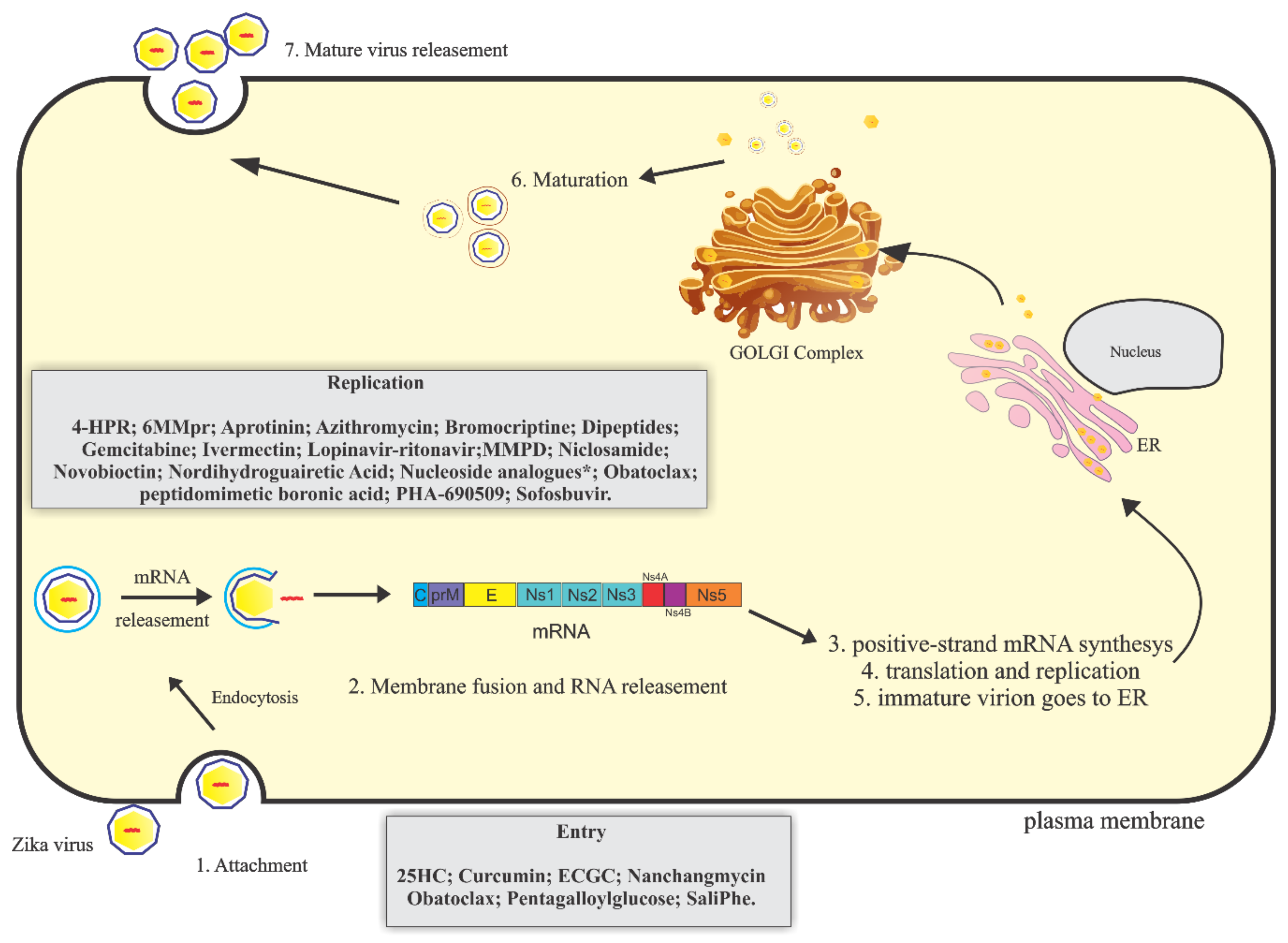
3. Therapeutical Potential of Anti-Zikv Molecules
3.1. Drug Repurposing
3.2. In Silico Interaction Analysis and Natural Drugs
3.2.1. Potential Inhibitors of the Early Stages of Replication
3.2.2. Potential Inhibitors of the Late Stages of Replication
4. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Dick, G.W. Zika Virus (II). Pathogenicity and Physical Properties. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 521–534. [Google Scholar] [CrossRef]
- Vinet, L.; Zhedanov, A. A “missing” family of Classical Orthogonal Polynomials. Emerg. Infect. Dis. 2010, 15, 1347–1350. [Google Scholar] [CrossRef]
- Dick, G.W.; Kitchen, S.; Haddow, A. Zika Virus (I). Isolations and Serological Specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Marano, G.; Pupella, S.; Vaglio, S.; Liumbruno, G.M.; Grazzini, G. Zika Virus and the Never-Ending Story of Emerging Pathogens and Transfusion Medicine. Blood Transfus. 2015, 14, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Simpson, D.I.H. Zika Virus Infection in Man. Trans. R. Soc. Trop. Med. Hyg. 1964, 58, 335–337. [Google Scholar] [CrossRef]
- Aubry, M.; Richard, V.; Green, J.; Broult, J.; Musso, D. Inactivation of Zika Virus in Plasma with Amotosalen and Ultraviolet a Illumination. Transfusion 2016, 56, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.R.; Chen, T.-H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.D.; Schuh, A.J.; Yasuda, C.Y.; Kasper, M.R.; Heang, V.; Huy, R.; Guzman, H.; Tesh, R.B.; Weaver, S.C. Genetic Characterization of Zika Virus Strains: Geographic Expansion of the Asian Lineage. PLoS Negl. Trop. Dis. 2012, 6, e1477. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.G.; Ksiazek, T.G.; Suhandiman; Triwibowo. Zika Virus, a Cause of Fever in Central Java, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 389–393. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.-M. Zika Virus, French Polynesia, South Pacific, 2013. Emerg. Infect. Dis. 2014, 20, 1960. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.W.; Choi, G.K.Y.; Yip, C.C.Y.; Cheng, V.C.C.; Yuen, K.-Y. Zika Fever and Congenital Zika Syndrome: An Unexpected Emerging Arboviral Disease. J. Infect. 2016, 72, 507–524. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Situation Report: Zika Virus, Microcephaly, Guillain-Barré Syndrome, 10 March 2017; WHO: Geneva, Switzerland, 2017; pp. 1–5. [Google Scholar]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an Arbovirus Vector: From the Darkness to the Light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.S.J.; Li, M.; Zhi, I.; Chong, C.S.; Ng, L.C.; Tan, C.H. Aedes (Stegomyia) albopictus (Skuse): A Potential Vector of Zika Virus in Singapore. PLoS Negl. Trop. Dis. 2013, 7, e2348. [Google Scholar] [CrossRef] [PubMed]
- Calvez, E.; Guillaumot, L.; Millet, L.; Marie, J.; Bossin, H.; Rama, V.; Faamoe, A.; Kilama, S.; Teurlai, M.; Mathieu-Daudé, F.; et al. Genetic Diversity and Phylogeny of Aedes aegypti, the Main Arbovirus Vector in the Pacific. PLoS Negl. Trop. Dis. 2016, 10, e0004374. [Google Scholar] [CrossRef] [PubMed]
- Gasperi, G.; Bellini, R.; Malacrida, A.R.; Crisanti, A.; Dottori, M.; Aksoy, S. A New Threat Looming over the Mediterranean Basin: Emergence of Viral Diseases Transmitted by Aedes albopictus Mosquitoes. PLoS Negl. Trop. Dis. 2012, 6, e1836. [Google Scholar] [CrossRef] [PubMed]
- Petersen, E.; Wilson, M.E.; Touch, S.; McCloskey, B.; Mwaba, P.; Bates, M.; Dar, O.; Mattes, F.; Kidd, M.; Ippolito, G.; et al. Rapid Spread of Zika Virus in The Americas—Implications for Public Health Preparedness for Mass Gatherings at the 2016 Brazil Olympic Games. Int. J. Infect. Dis. 2016, 44, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Darwish, M.A.; Hoogstraal, H.; Roberts, T.J.; Ahmed, I.P.; Omar, F. A Sero-Epidemiological Survey for Certain Arboviruses (Togaviridae) in Pakistan. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 442–445. [Google Scholar] [CrossRef]
- Vorou, R. Zika Virus, Vectors, Reservoirs, Amplifying Hosts, and Their Potential to Spread Worldwide: What We Know and What We Should Investigate Urgently. Int. J. Infect. Dis. 2016, 48, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Nhan, T.; Robin, E.; Roche, C.; Bierlaire, D.; Zisou, K.; Shan Yan, A.; Cao-Lormeau, V.M.; Broult, J. Potential for Zika Virus Transmission through Blood Transfusion Demonstrated during an Outbreak in French Polynesia, November 2013 to February 2014. Eurosurveillance 2014, 19. [Google Scholar] [CrossRef]
- McCarthy, M. Zika Virus Was Transmitted by Sexual Contact in Texas, Health Officials Report. BMJ 2016, 352, i720. [Google Scholar] [CrossRef] [PubMed]
- Ventura, C.V.; Maia, M.; Ventura, B.V.; Van Der Linden, V.; Araújo, E.B.; Ramos, R.C.; Rocha, M.A.W.; Carvalho, M.D.C.G.; Belfort, R., Jr.; Ventura, L.O. Ophthalmological Findings in Infants with Microcephaly and Presumable Intra-Uterus Zika Virus Infection. Arq. Bras. Oftalmol. 2016, 79, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Perreira, J.M.; Chin, C.R.; Feeley, E.M.; Brass, A.L. IFITMs Restrict the Replication of Multiple Pathogenic Viruses. J. Mol. Biol. 2013, 425, 4937–4955. [Google Scholar] [CrossRef] [PubMed]
- Samarasekera, U.; Triunfol, M. Concern over Zika Virus Grips the World. Lancet 2016, 387, 521–524. [Google Scholar] [CrossRef]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Sirohi, D.; Chen, Z.; Sun, L.; Klose, T.; Pierson, T.C.; Rossmann, M.G.; Kuhn, R.J. The 3.8 Å Resolution Cryo-EM Structure of Zika Virus. Science 2016, 5316, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus Genome Organization, Expression, and Replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Buathong, R.; Hermann, L.; Thaisomboonsuk, B.; Rutvisuttinunt, W.; Klungthong, C.; Chinnawirotpisan, P.; Manasatienkij, W.; Nisalak, A.; Fernandez, S.; Yoon, I.K.; et al. Detection of Zika Virus Infection in Thailand, 2012–2014. Am. J. Trop. Med. Hyg. 2015, 93, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Van Hemert, F.; Berkhout, B. Nucleotide Composition of the Zika Virus RNA Genome and Its Codon Usage. Virol. J. 2016, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lindenbach, B.D.; Rice, C.M. Molecular Biology of Flavivirus. Adv. Virus Res. 2003, 59, 23–61. [Google Scholar] [CrossRef] [PubMed]
- Bollati, M.; Alvarez, K.; Assenberg, R.; Baronti, C.; Canard, B.; Cook, S.; Coutard, B.; Decroly, E.; de Lamballerie, X.; Gould, E.A.; et al. Structure and Functionality in Flavivirus NS-Proteins: Perspectives for Drug Design. Antiviral Res. 2010, 87, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Shapshak, P.; Somboonwit, C.; Foley, B.T.; Alrabaa, S.F.; Wills, T.; Sinnott, J.T. Zika Virus. In Global Virology I—Identifying and Investigating Viral Diseases; Springer: New York, NY, USA, 2015; pp. 477–500. [Google Scholar]
- Adiga, R. Phylogenetic Analysis of the NS5 Gene of Zika Virus. J. Med. Virol. 2016, 88, 1821–1826. [Google Scholar] [CrossRef] [PubMed]
- Cugola, F.R.; Fernandes, I.R.; Russo, F.B.; Freitas, B.C.; Dias, J.L.M.; Guimarães, K.P.; Benazzato, C.; Almeida, N.; Pignatari, G.C.; Romero, S.; et al. The Brazilian Zika Virus Strain Causes Birth Defects in Experimental Models. Nature 2016, 534, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Valderramos, S.G.; Wu, A.; Ouyang, S.; Li, C.; Brasil, P.; Bonaldo, M.; Coates, T.; Nielsen-Saines, K.; Jiang, T.; et al. From Mosquitos to Humans: Genetic Evolution of Zika Virus. Cell Host Microbe 2016, 19, 561–565. [Google Scholar] [CrossRef] [PubMed]
- WHO. Current Zika Product Pipeline; WHO: Geneva, Switzerland, 2016; pp. 1–16. [Google Scholar]
- Atif, M.; Azeem, M.; Sarwar, M.R.; Bashir, A. Zika Virus Disease: A Current Review of the Literature. Infection 2016, 44, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control (CDC). Zika: The Basics of the Virus and How to Protect against It; Centers for Disease Control: Atlanta, GA, USA, 2016.
- Chan, J.F.W.; Chik, K.K.H.; Yuan, S.; Yip, C.C.Y.; Zhu, Z.; Tee, K.M.; Tsang, J.O.L.; Chan, C.C.S.; Poon, V.K.M.; Lu, G.; et al. Novel Antiviral Activity and Mechanism of Bromocriptine as a Zika Virus NS2B-NS3 Protease Inhibitor. Antiviral Res. 2017, 141, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Lee, E.M.; Wen, Z.; Cheng, Y.; Huang, W.-K.; Qian, X.; Tcw, J.; Kouznetsova, J.; Ogden, S.C.; Hammack, C.; et al. Identification of Small-Molecule Inhibitors of Zika Virus Infection and Induced Neural Cell Death via a Drug Repurposing Screen. Nat. Publ. Gr. 2016, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Barrows, N.J.; Campos, R.K.; Powell, S.T.; Prasanth, K.R.; Schott-Lerner, G.; Soto-Acosta, R.; Galarza-Muñoz, G.; McGrath, E.L.; Urrabaz-Garza, R.; Gao, J.; Wu, P.; et al. A Screen of FDA-Approved Drugs for Inhibitors of Zika Virus Infection. Cell Host Microbe 2016, 20, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Delvecchio, R.; Higa, L.; Pezzuto, P.; Valadão, A.; Garcez, P.; Monteiro, F.; Loiola, E.; Dias, A.; Silva, F.; Aliota, M.; et al. Chloroquine, an Endocytosis Blocking Agent, Inhibits Zika Virus Infection in Different Cell Models. Viruses 2016, 8, 322. [Google Scholar] [CrossRef] [PubMed]
- Mounce, B.C.; Cesaro, T.; Carrau, L.; Vallet, T.; Vignuzzi, M. Curcumin Inhibits Zika and Chikungunya Virus Infection by Inhibiting Cell Binding. Antiviral Res. 2017, 142, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.M.; Batista, M.N.; Braga, A.C.S.; Nogueira, M.L.; Rahal, P. The Green Tea Molecule EGCG Inhibits Zika Virus Entry. Virology 2016, 496, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Arbuckle, J.H.; Gardina, P.J.; Gordon, D.N.; Hickman, H.D.; Yewdell, J.W.; Pierson, T.C.; Myers, T.G.; Kristie, T.M. Inhibitors of the Histone Methyltransferases EZH2/1 Induce a Potent Antiviral State and Suppress Infection by Diverse Viral Pathogens. MBio 2017, 8, e01141-17. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, S.; Cooper, L.; Rubio, A.; Pagani, I.; Capobianchi, M.R.; Ippolito, G.; Pelletier, J.; Meneghetti, M.C.Z.; Lima, M.A.; Skidmore, M.A.; et al. Heparin Prevents Zika Virus Induced-Cytopathic Effects in Human Neural Progenitor Cells. Antiviral Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Rausch, K.; Hackett, B.A.; Weinbren, N.L.; Reeder, S.M.; Sadovsky, Y.; Hunter, C.A.; Schultz, D.C.; Coyne, C.B.; Cherry, S. Screening Bioactives Reveals Nanchangmycin as a Broad Spectrum Antiviral Active against Zika Virus. Cell Rep. 2017, 18, 804–815. [Google Scholar] [CrossRef] [PubMed]
- Kuivanen, S.; Bespalov, M.M.; Nandania, J.; Ianevski, A.; Velagapudi, V.; De Brabander, J.K.; Kainov, D.E.; Vapalahti, O. Obatoclax, Saliphenylhalamide and Gemcitabine Inhibit Zika Virus Infection in Vitro and Differentially Affect Cellular Signaling, Transcription and Metabolism. Antiviral Res. 2016, 139, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, P.; Perin, P.; Menzel, N.; Banda, D.; Pfaender, S.; Alves, M.P.; Thiel, V.; Meulemann, P.; Colpitts, C.C.; Schang, L.M.; et al. Pentagalloylglucose, a Highly Bioavailable Polyphenolic Compound Present in Cortex Moutan, Efficiently Blocks Hepatitis C Virus Entry. Antiviral Res. 2017, 147, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Deng, Y.; Wang, S.; Ma, F.; Aliyari, R.; Huang, X.-Y.; Zhang, N.-N.; Watanabe, M.; Dong, H.-L.; Liu, P.; et al. 25-Hydroxycholesterol Protects Host against Zika Virus Infection and Its Associated Microcephaly in a Mouse Model. Immunity 2017, 46, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Sacramento, C.Q.; De Melo, G.R.; De Freitas, C.S.; Marttorelli, A.; Ferreira, A.C.; Barbosa-lima, G.; Volotão, M.; Nunes, E.P.; Tschoeke, D.A.; Leomil, L. The Clinically Approved Antiviral Drug Sofosbuvir Inhibits Zika Virus Replication. Nat. Publ. Gr. 2017, 1–11. [Google Scholar] [CrossRef]
- Yuan, S.; Chan, J.F.-W.; Den-Haan, H.; Chik, K.K.-H.; Zhang, A.J.; Chan, C.C.-S.; Poon, V.K.-M.; Yip, C.C.-Y.; Mak, W.W.-N.; Zhu, Z.; et al. Structure-Based Discovery of Clinically Approved Drugs as Zika Virus NS2B-NS3 Protease Inhibitors That Potently Inhibit Zika Virus Infection In Vitro and In Vivo. Antiviral Res. 2017, 145, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Hansen, G.; Nitsche, C.; Klein, C.D.; Zhang, L.; Hilgenfeld, R. Crystal Structure of Zika Virus NS2B-NS3 Protease in Complex with a Boronate Inhibitor. Science 2016, 353, 503–505. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, K.; Wu, C.; Chen, C.; Hu, C.; Buzovetsky, O.; Wang, Z.; Ji, X.; Xiong, Y.; Yang, H. Mechanisms of Activation and Inhibition of Zika Virus NS2B-NS3 Protease. Cell Res. 2016, 26, 1260–1263. [Google Scholar] [CrossRef] [PubMed]
- Barbosa-Lima, G.; Moraes, A.M.; da Araújo, A.S.; da Silva, E.T.; de Freitas, C.S.; Vieira, Y.R.; Marttorelli, A.; Neto, J.C.; Bozza, P.T.; de Souza, M.V.N.; Souza, T.M.L. 2,8-Bis(trifluoromethyl)quinoline Analogs Show Improved Anti-Zika Virus Activity, Compared to Mefloquine. Eur. J. Med. Chem. 2017, 127, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Smith, J.; Bukreyeva, N.; Koma, T.; Manning, J.T.; Kalkeri, R.; Kwong, A.D.; Paessler, S. Merimepodib, an IMPDH Inhibitor, Suppresses Replication of Zika Virus and Other Emerging Viral Pathogens. Antiviral Res. 2018, 149, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Byler, K.G.; Ogungbe, I.V.; Setzer, W.N. In-Silico Screening for Anti-Zika Virus Phytochemicals. J. Mol. Graph. Model. 2016, 69, 78–91. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, O.V.; Félix, D.M.; de Mendonça, L.R.; de Araújo, C.M.C.S.; de Oliveira Franca, R.F.; Cordeiro, M.T.; Silva Júnior, A.; Pena, L.J. The Thiopurine Nucleoside Analogue 6-Methylmercaptopurine Riboside (6MMPr) Effectively Blocks Zika Virus Replication. Int. J. Antimicrob. Agents 2017, 50, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Merino-Ramos, T.; Jiménez de Oya, N.; Saiz, J.; Martín-Acebes, M.A. Antiviral Activity of Nordihydroguaiaretic Acid and Its Derivative Tetra-O-Methyl Nordihydroguaiaretic Acid against West Nile Virus and Zika Virus. Antimicrob. Agents Chemother. 2017, 61, e00376-17. [Google Scholar] [CrossRef] [PubMed]
- Eyer, L.; Nencka, R.; Huvarová, I.; Palus, M.; Joao Alves, M.; Gould, E.A.; De Clercq, E.; Růžek, D. Nucleoside Inhibitors of Zika Virus. J. Infect. Dis. 2016, 214, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, E.; Pezzullo, M.; De Burghgraeve, T.; Kaptein, S.; Pastorino, B.; Dallmeier, K.; De lamballerie, X.; Neyts, J.; Hanson, A.M.; Frick, D.N.; et al. Ivermectin Is a Potent Inhibitor of Flavivirus Replication Specifically Targeting NS3 Helicase Activity: New Prospects for an Old Drug. J. Antimicrob. Chemother. 2012, 67, 1884–1894. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Zachariah, M.; Harris, E. Mycophenolic Acid Inhibits Dengue Virus Infection by Preventing Replication of Viral RNA. Virology 2002, 304, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Retallack, H.; Di Lullo, E.; Arias, C.; Knopp, K.A.; Laurie, M.T.; Sandoval-Espinosa, C.; Leon, W.R.M.; Krencik, R.; Ullian, E.M.; Spatazza, J.; et al. Zika virus cell tropism in the developing human brain and inhibition by azithromycin. Proc. Natl. Acad. Sci. USA 2016, 113, 14408–14413. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhu, X.; Ji, X.; Quanquin, N.; Deng, Y.Q.; Tian, M.; Aliyari, R.; Zuo, X.; Yuan, L.; Afridi, S.K.; et al. Chloroquine, a FDA-approved Drug, Prevents Zika Virus Infection and its Associated Congenital Microcephaly in Mice. EBioMedicine 2017, 24, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Perachon, S.; Schwartz, J.C.; Sokoloff, P. Functional Potencies of New Antiparkinsonian Drugs at Recombinant Human Dopamine D1, D2 and D3 receptors. Eur. J. Pharmacol. 1999, 366, 293–300. [Google Scholar] [CrossRef]
- Ginther, O.J.; Santos, V.G.; Mir, R.A.; Beg, M.A. Role of LH in the Progesterone Increase during the Bromocriptine-Induced Prolactin Decrease in Heifers. Theriogenology 2012, 78, 1969–1976. [Google Scholar] [CrossRef] [PubMed]
- Kato, F.; Ishida, Y.; Oishi, S.; Fujii, N.; Watanabe, S.; Vasudevan, S.G.; Tajima, S.; Takasaki, T.; Suzuki, Y.; Ichiyama, K.; et al. Novel Antiviral Activity of Bromocriptine against Dengue Virus Replication. Antiviral Res. 2016, 131, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Keller, T.H.; Luo, D. Zika Virus Protease: An Antiviral Drug Target. Trends Microbiol. 2017, 25, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ren, J.; Nocadello, S.; Rice, A.J.; Ojeda, I.; Light, S.; Minasov, G.; Vargas, J.; Nagarathnam, D.; Anderson, W.F.; et al. Identification of Novel Small Molecule Inhibitors against NS2B/NS3 Serine Protease from Zika Virus. Antiviral Res. 2017, 139, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Z.; Phoo, W.W.; Loh, Y.R.; Wang, W.; Liu, S.; Chen, M.W.; Hung, A.W.; Keller, T.H.; Luo, D.; et al. Structural Dynamics of Zika Virus NS2B-NS3 Protease Binding to Dipeptide Inhibitors. Structure 2017, 25, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Setting. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Söderholm, S.; Anastasina, M.; Islam, M.M.; Tynell, J.; Poranen, M.M.; Bamford, D.H.; Stenman, J.; Julkunen, I.; Šauliene, I.; De Brabander, J.K.; et al. Immuno-Modulating Properties of Saliphenylhalamide, SNS-032, Obatoclax, and Gemcitabine. Antiviral Res. 2016, 126, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Nance, C.L.; Siwak, E.B.; Shearer, W.T. Preclinical Development of the Green Tea Catechin, Epigallocatechin Gallate, as an HIV-1 Therapy. J. Allergy Clin. Immunol. 2009, 123, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, C.E.; Wen, G.Y.; Xu, W.; Jia, J.H.; Rohan, L.; Corbo, C.; Di Maggio, V.; Jenkins, E.C.; Hillier, S. Epigallocatechin Gallate Inactivates Clinical Isolates of Herpes Simplex Virus. Antimicrob. Agents Chemother. 2008, 52, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Song, J.M.; Lee, K.H.; Seong, B.L. Antiviral Effect of Catechins in Green Tea on Influenza Virus. Antiviral Res. 2005, 68, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Calland, N.; Dubuisson, J.; Rouillé, Y.; Séron, K. Hepatitis C Virus and Natural Compounds: A New Antiviral Approach? Viruses 2012, 4, 2197–2217. [Google Scholar] [CrossRef] [PubMed]
- Raekiansyah, M.; Buerano, C.C.; Luz, M.A.D.; Morita, K. Inhibitory Effect of the Green Tea Molecule EGCG against Dengue Virus Infection. Arch. Virol. 2018, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, Y.; Guo, J.; Wang, P.; Zhang, L.; Xiao, G.; Wang, W. Screening of FDA-Approved Drugs for Inhibitors of Japanese Encephalitis Virus Infection. J. Virol. 2017, 91, e01055-17. [Google Scholar] [CrossRef] [PubMed]
- Shome, S.; Das Talukdar, A.; Choudhury, M.D.; Bhattacharya, M.K.; Upadhyaya, H. Curcumin as Potential Therapeutic Natural Product: A Nanobiotechnological Perspective. J. Pharm. Pharmacol. 2016, 68, 1481–1500. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E. Guanosine Analogues as Anti-Herpesvirus Agents. Nucleosides Nucleotides Nucleic Acids 2000, 19, 1531–1541. [Google Scholar] [CrossRef] [PubMed]
- Beadle, J.R.; Wan, W.B.; Ciesla, S.L.; Keith, K.A.; Hartline, C.; Kern, E.R.; Hosteller, K.Y. Synthesis and Antiviral Evaluation of Alkoxyalkyl Derivatives of 9-(S)-(3-Hydroxy-2-Phosphonomethoxypropyl)adenine against Cytomegalovirus and Orthopoxviruses. J. Med. Chem. 2006, 49, 2010–2015. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Asano, M.; Matsumoto, A.; Sugiyama, M.; Nishida, N.; Tanaka, E.; Inoue, T.; Sakamoto, M.; Enomoto, N.; Shirasaki, T.; et al. Induction of IFN-λ3 as an Additional Effect of Nucleotide, Not Nucleoside, Analogues: A New Potential Target for HBV Infection. Gut 2016, 67, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Santiago, B.I.; Chen, H.; Asif, G.; Beltran, T.; Mao, S.; Hurwitz, S.J.; Grier, J.; McClure, H.M.; Chu, C.K.; Liotta, D.C.; et al. Pharmacology and Pharmacokinetics of the Antiviral Agent Beta-D-2′,3′-dideoxy-3′-oxa-5-Fluorocytidine in Cells and Rhesus Monkeys. Antimicrob. Agents Chemother. 2005, 49, 2589–2597. [Google Scholar] [CrossRef] [PubMed]
- Asif, G.; Hurwitz, S.J.; Obikhod, A.; Delinsky, D.; Narayanasamy, J.; Chu, C.K.; McClure, H.M.; Schinazi, R.F. Pharmacokinetics of the Anti-Human Immunodeficiency Virus Agent 1-(Beta-d-Dioxolane)thymine in Rhesus Monkeys. Antimicrob. Agents Chemother. 2007, 51, 2424–2429. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, H.; Boudinot, F.D.; Chu, C.K.; Mcclure, H.M. Pharmacokinetics of 2-Aminopurine Dioxolane and D-2-Amino-6-Chloropurine Dioxolane and Their Antiviral Metabolite-Dioxolane Guanine in Rhesus Monkeys. Antimicrob. Agentes Chemother. 1996, 40, 2332–2336. [Google Scholar]
- Mao, S.; Bouygues, M.; Welch, C.; Biba, M.; Chilenski, J.; Schinazi, R.F.; Liotta, D.C. Synthesis of Enantiomerically Pure D-FDOC, an Anti-HIV Agent. Bioorg. Med. Chem. Lett. 2004, 14, 4991–4994. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.S.; Tomassini, J.E.; Bosserman, M.; Getty, K.; Stahlhut, M.W.; Eldrup, A.B.; Bhat, B.; Hall, D.; Simcoe, A.L.; LaFemina, R.; et al. Inhibition of Hepatitis C Virus RNA Replication by 2′-modified Nucleoside Analogs. J. Biol. Chem. 2003, 278, 11979–11984. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Nettles, R.E.; Belema, M.; Snyder, L.B.; Nguyen, V.N.; Fridell, R.A.; Serrano-Wu, M.H.; Langley, D.R.; Sun, J.H.; O′Boyle II, D.R.; et al. Chemical genetics strategy identifies an HCV NS5A inhibitor with a potent clinical effect. Nature 2010, 465, 96. [Google Scholar] [CrossRef] [PubMed]
- Gentile, I.; Maraolo, A.E.; Buonomo, A.R.; Zappulo, E.; Borgia, G. The discovery of sofosbuvir: a revolution for therapy of chronic hepatitis C. Expert Opin. Drug Discov. 2015, 10, 1363–1377. [Google Scholar] [CrossRef] [PubMed]
- Mesci, P.; Macia, A.; Moore, S.M.; Shiryaev, S.A.; Pinto, A.; Huang, C.T.; Tejwani, L.; Fernandes, I.R.; Suarez, N.A.; Kolar, M.J.; et al. Blocking Zika Virus Vertical Transmission. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Pryor, M.J.; Rawlinson, S.M.; Butcher, R.E.; Barton, C.L.; Waterhouse, T.A.; Vasudevan, S.G.; Bardin, P.G.; Wright, P.J.; Jans, D.A.; Davidson, A.D. Nuclear Localization of Dengue Virus Nonstructural Protein 5 Through Its Importin α/β–Recognized Nuclear Localization Sequences Is Integral to Viral Infection. Traffic 2007, 8, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, S.N.Y.; Smith, K.; Forwood, J.K.; Jans, D.A. Nuclear Import Inhibitor N-(4-Hydroxyphenyl) Retinamide Targets Zika Virus (ZIKV) Nonstructural Protein 5 to Inhibit ZIKV Infection. Biochem. Biophys. Res. Commun. 2017, 493, 1–5. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Compound | Validated Cell/Test | Anti-ZIKV Strategy | Reference | |
|---|---|---|---|---|
| Drug Repurposing | Bromocriptine | Vero cells | Inhibition of NS2B-NS3 protease | [39] |
| Emricasan | Gliobastoma SBN-19, hNPCS and human astrocytes cells | Reduces cellular apoptosis by inhibition of caspase-3 activities | [40] | |
| Ivermectin, Daptomycin, Mycophenolic acid (MPA), Sertraline, Pyrimethamine, Cyclosporine A, Azathioprine, Mefloquine | HuH 7 cells | Unknow | [41] | |
| Nicosamide, PHA-690509 | Gliobastoma SBN-19, hNPCS and human astrocytes cells | Inhibit viral replication | [40] | |
| Chloroquine | Vero, hNSC and hBMEC cells | Inhibits early stages of replicative cycle | [42] | |
| Bromocriptine | Vero cells | Inhibition of NS2B-NS3 protease | [39] | |
| Potential Inhibitors of the early and late stages of replication | Curcumin | HeLa cells | ZIKV entry and virucidal effect | [43] |
| Epigalocatequina galato | Vero cells | Inhibits the viral entry | [44] | |
| GSK126 | Telomerase reverse transcriptase-immortalized HFF cells | Strongly inhibited ZIKV infection in pre-treated cells | [45] | |
| Heparin | human neural progenitor cells (hNPCs) | Inhibits caspase 3 activity mediated by ZIKV infection. | [46] | |
| Nanchangmycin | U2OS cells | Blocks viral entry blocking clathrin-mediated endocytosis | [47] | |
| Obatoclax | Human telomerase reverse transcriptase-immortalized retinal pigment (RPE) | Inhibits endocytic uptake of ZIKV and viral protein synthesisPrevents caspase 8, 3 and 7 activationProtects the phosphorylation status of p27 phosphoprotein | [48] | |
| Pentagalloylglucose (PPG) | Vero B4 | ZIKV entry | [49] | |
| SaliPhe | human telomerase reverse transcriptase-immortalized retinal pigment (RPE) | Inhibits endocytic uptake of ZIKV and protein viral synthesisPrevents caspase 8, 3 and 7 activation | [48] | |
| 25-Hydroxycholesterol (25 HC) | BHKK-21 | Inhibits ZIKV internalization | [50] | |
| Sofosbuvir | HNPCs, Huh-7, SH-5YSY, Vero cells. Neurosphere, in silico | Nucleoside inhibitor; binds too amino acid residues critical for ribonucleotide incorporation; interacts strongly with ZIKV RNA polymerase | [51] | |
| Novobioctin, lopinavir-ritonavir | Vero/Huh-7/in silico | Inhibition of NS2B-NS3 protease | [52] | |
| Peptidomimetic boronic acid | Huh-7/in silico | Inhibition of NS2B-NS3 protease | [53] | |
| Aprotinin | In silico | Inhibition of NS2B-NS3 protease | [54] | |
| N-(4-hydroxyphenyl) retinamide (4-HPR) | Vero cells | Inhibition of viral replication probably interacting with NS5 protein | [55] | |
| Merimepodib (MMPD) | Huh7 | Inhibits ZIKV RNA replication inhibiting IMPDH (inosine-5′-monophosphate dehydrogenase) | [56] | |
| Gemcitabine | Human telomerase reverse transcriptase-immortalized retinal pigment (RPE) | Interferes with transcription of viral RNAInhibits viral protein synthesisPrevents caspase 8, 3 and 7 activationChanges the phosphorylation status of the CREB phosphoprotein affected by ZIKV infection | [48] | |
| Cimiphenol, Cimiracemate B, Rosemarinic acid | In silico | high affinity with NS5 methyl transferase | [57] | |
| 6-methylmercaptopurine riboside (6 MMpr) | Vero and SII—SY5Y neuronal cells | 6 MMpr used during viral RNA synthesis reducing the viral infectivity caused by mutations and viral replication errors | [58] | |
| 4,7-digalloylcatechin, Prenylated chalcone, 2,4,4-trihydroxy-3,3-diprenylchalcone,Bis-indole alkaloid flinderoleLignan di-O-demethyl isoguaiacin | In silico | High affinity with NS5 RNA-dependent RNA polymerase | [57] | |
| Bis-indole alkaloids flinderole A and flinderole B | In silico | High affinity with NS2B-NS3 protease | [57] | |
| Cimiphenol, Cimiracemate B, Rosemarinic acid | In silico | High affinity with NS5 methyl transferase | [57] | |
| Isoquinoline alkaloid cassiarin D3-Omethyldiplacone, Exiguaflavanone A, Sesqui terpenoid lactucopicrinAurone kanzonol V, Chalconesvangusticornin BBalsacone Bkanzonol Ylignans hibalactone kaerophyllin | In silico | Inhibition of NS3 helicase ATP site | [57] | |
| Nordihydroguairetic Acid | Vero | Disturbs the lipid metabolism | [59] | |
| Nucleoside analogues: 7-desaza-2′-C-methyladenosine (7-desaza-2′-CMA)2′-C-methyladenosine (2′-CMA)2′-C-methylcytidine (2′ CMC)2′-C-methylguanosine (2′-CMG)2′-C-methyluridine (2′-CMU) | Vero, Human neuroblastoma (UKF-NB-4 and porcine kidney (PS) cells | Reduce cytopathic effect, cell death and inhibit the viral replication | [60] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Da Silva, S.; Oliveira Silva Martins, D.; Jardim, A.C.G. A Review of the Ongoing Research on Zika Virus Treatment. Viruses 2018, 10, 255. https://doi.org/10.3390/v10050255
Da Silva S, Oliveira Silva Martins D, Jardim ACG. A Review of the Ongoing Research on Zika Virus Treatment. Viruses. 2018; 10(5):255. https://doi.org/10.3390/v10050255
Chicago/Turabian StyleDa Silva, Suely, Daniel Oliveira Silva Martins, and Ana Carolina Gomes Jardim. 2018. "A Review of the Ongoing Research on Zika Virus Treatment" Viruses 10, no. 5: 255. https://doi.org/10.3390/v10050255
APA StyleDa Silva, S., Oliveira Silva Martins, D., & Jardim, A. C. G. (2018). A Review of the Ongoing Research on Zika Virus Treatment. Viruses, 10(5), 255. https://doi.org/10.3390/v10050255