Inhibition of v-rel-Induced Oncogenesis through microRNA Targeting


Abstract
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Svoboda, J. Rous sarcoma virus centennial in folia biologica. Folia Biol. 2013, 59, 103–104. [Google Scholar]
- Varmus, H. How tumor virology evolved into cancer biology and transformed oncology. Annu. Rev. Cancer Biol. 2017, 1, 1–18. [Google Scholar] [CrossRef]
- Nair, V.; Fadly, A. Leukosis/sarcoma group. In Diseases of Poultry, 13th ed.; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V., Eds.; John Wiley & Sons, Inc.: Ames, IA, USA, 2013; pp. 553–592. [Google Scholar]
- Pishkari, S.; Paryan, M.; Hashemi, M.; Baldini, E.; Mohammadi-Yeganeh, S. The role of microRNAs in different types of thyroid carcinoma: A comprehensive analysis to find new miRNA supplementary therapies. J. Endocrinol. Investig. 2018, 41, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of tissue-specific microRNAs from mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef]
- Ebert, M.S.; Sharp, P.A. MicroRNA sponges: Progress and possibilities. RNA 2010, 16, 2043–2050. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhou, Y.; Zhang, Y.; He, L.; Xue, C.; Cao, Y. Design of miRNA sponges for MDV-1 as a therapeutic strategy against lymphomas. Oncotarget 2018, 9, 3842–3852. [Google Scholar] [CrossRef] [PubMed]
- Gentner, B.; Schira, G.; Giustacchini, A.; Amendola, M.; Brown, B.D.; Ponzoni, M.; Naldini, L. Stable knockdown of microRNA in vivo by lentiviral vectors. Nat. Methods 2009, 6, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Tay, F.C.; Lim, J.K.; Zhu, H.; Hin, L.C.; Wang, S. Using artificial microRNA sponges to achieve microRNA loss-of-function in cancer cells. Adv. Drug Deliv. Rev. 2015, 81, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.D.; Cantore, A.; Annoni, A.; Sergi, L.S.; Lombardo, A.; Della Valle, P.; D’Angelo, A.; Naldini, L. A microRNA-regulated lentiviral vector mediates stable correction of hemophilia B mice. Blood 2007, 110, 4144–4152. [Google Scholar] [CrossRef] [PubMed]
- Edge, R.E.; Falls, T.J.; Brown, C.W.; Lichty, B.D.; Atkins, H.; Bell, J.C. A let-7 microRNA-sensitive vesicular stomatitis virus demonstrates tumor-specific replication. Mol. Ther. 2008, 16, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.J.; Hadac, E.M.; Greiner, S.; Russell, S.J. Engineering microRNA responsiveness to decrease virus pathogenicity. Nat. Med. 2008, 14, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.J.; Russell, S.J. MicroRNAs and the regulation of vector tropism. Mol. Ther. 2009, 17, 409–416. [Google Scholar] [CrossRef] [PubMed]
- TenOever, B.R. RNA viruses and the host microRNA machinery. Nat. Rev. Microbiol. 2013, 11, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.D.; Venneri, M.A.; Zingale, A.; Sergi Sergi, L.; Naldini, L. Endogenous microRNA regulation suppresses transgene expression in hematopoietic lineages and enables stable gene transfer. Nat. Med. 2006, 12, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.; Kunitomi, M.; Vignuzzi, M.; Saksela, K.; Andino, R. Harnessing endogenous miRNAs to control virus tissue tropism as a strategy for developing attenuated virus vaccines. Cell Host Microbe 2008, 4, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Cawood, R.; Wong, S.L.; Di, Y.; Baban, D.F.; Seymour, L.W. MicroRNA controlled adenovirus mediates anti-cancer efficacy without affecting endogenous microRNA activity. PLoS ONE 2011, 6, e16152. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.J.; Hadac, E.M.; Cullen, B.R.; Russell, S.J. MicroRNA antagonism of the picornaviral life cycle: Alternative mechanisms of interference. PLoS Pathog. 2010, 6, e1000820. [Google Scholar] [CrossRef] [PubMed]
- Perez, J.T.; Pham, A.M.; Lorini, M.H.; Chua, M.A.; Steel, J.; tenOever, B.R. MicroRNA-mediated species-specific attenuation of influenza a virus. Nat. Biotechnol. 2009, 27, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Teterina, N.L.; Maximova, O.A.; Kenney, H.; Liu, G.; Pletnev, A.G. MicroRNA-based control of tick-borne flavivirus neuropathogenesis: Challenges and perspectives. Antivir. Res. 2016, 127, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.H. The rcas vector system. Folia Biol. 2004, 50, 107–119. [Google Scholar]
- Yao, Y.; Vasoya, D.; Kgosana, L.; Smith, L.P.; Gao, Y.; Wang, X.; Watson, M.; Nair, V. Activation of gga-miR-155 by reticuloendotheliosis virus T strain and its contribution to transformation. J. Gen. Virol. 2017, 98, 810–820. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs modulate hematopoietic lineage differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Carraro, G.; El Agha, E.; Mukhametshina, R.; Chao, C.M.; Rizvanov, A.; Barreto, G.; Bellusci, S. Generation and validation of miR-142 knock out mice. PLoS ONE 2015, 10, e0136913. [Google Scholar] [CrossRef] [PubMed]
- Lambeth, L.S.; Yao, Y.; Smith, L.P.; Zhao, Y.; Nair, V. MicroRNAs 221 and 222 target p27Kip1 in marek’s disease virus-transformed tumour cell line MSB-1. J. Gen. Virol. 2009, 90, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T.D.; Wolenski, F.S. NF-κB: Where did it come from and why? Immunol. Rev. 2012, 246, 14–35. [Google Scholar] [CrossRef] [PubMed]
- Liss, A.S.; Tiwari, R.; Kralova, J.; Bose, H.R., Jr. Cell transformation by v-rel reveals distinct roles of AP-1 family members in Rel/NF-κB oncogenesis. Oncogene 2010, 29, 4925–4937. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 years of NF-κB: A blossoming of relevance to human pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Hrdlickova, R.; Nehyba, J.; Humphries, E.H. V-rel induces expression of three avian immunoregulatory surface receptors more efficiently than c-rel. J. Virol. 1994, 68, 308–319. [Google Scholar] [PubMed]
- Fenton, S.P.; Reddy, M.R.; Bagust, T.J. Single and concurrent avian leukosis virus infections with avian leukosis virus-J and avian leukosis virus-A in Australian meat-type chickens. Avian Pathol. 2005, 34, 48–54. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bolisetty, M.T.; Dy, G.; Tam, W.; Beemon, K.L. Reticuloendotheliosis virus strain T induces miR-155, which targets JARID2 and promotes cell survival. J. Virol. 2009, 83, 12009–12017. [Google Scholar] [CrossRef] [PubMed]




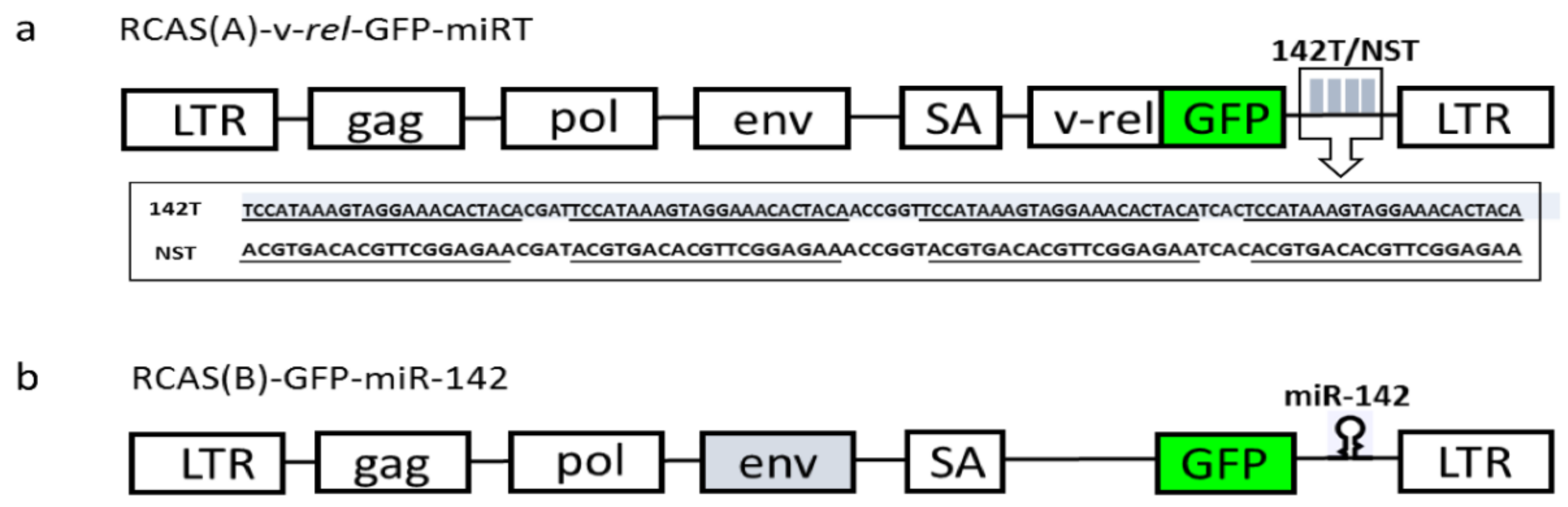{kind=link}
{kind=link}
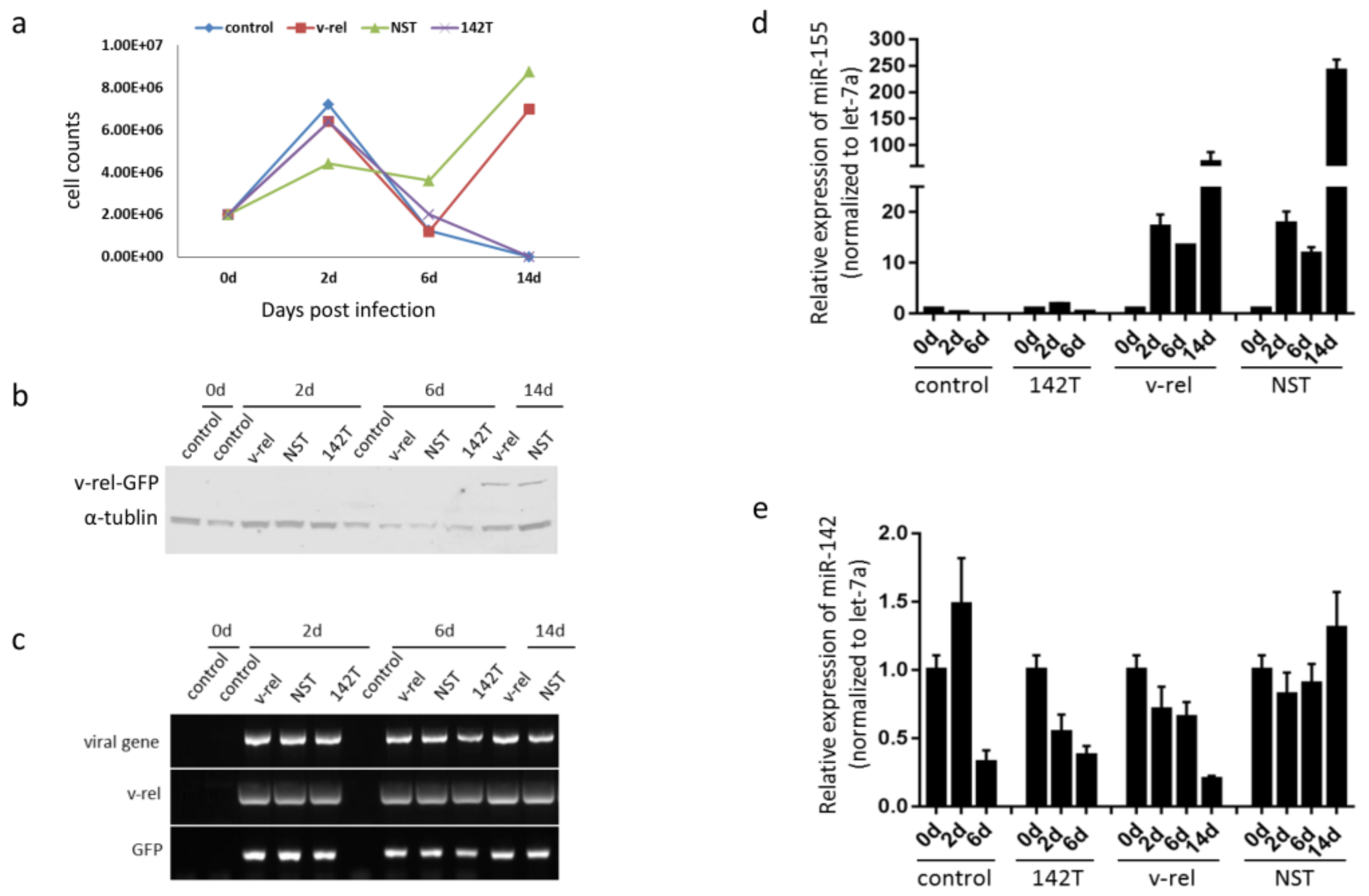{kind=link}
| Primer | Sequence (5’–3’) |
|---|---|
| miR-142T-F | GGCCGCTCCATAAAGTAGGAAACACTACACGATTCCATAAAGTAGGAAACACTACAACCGGTT CCATAAAGTAGGAAACACTACATCACTCCATAAAGTAGGAAACACTACAA |
| miR-142T-R | GGCCGCACGTGACACGTTCGGAGAACGATACGTGACAC GTTCGGAGAAACCGGTACGTGACACGTTCGGAGAATCA CACGTGACACGTTCGGAGAAA |
| miR-NST-F | GGCCGCACGTGACACGTTCGGAGAACGATACGTGACAC GTTCGGAGAAACCGGTACGTGACACGTTCGGAGAATCA CACGTGACACGTTCGGAGAAA |
| miR-NST-R | CTAGTTTCTCCGAACGTGTCACGTGTGATTCTCCGAACG TGTCACGTACCGGTTTCTCCGAACGTGTCACGTATCGTTCTCCGAACGTGTCACGTGC |
| miR-142-F | GGCCATAATGGCCGGGATGTCCCCTGTGCCCCACTC |
| miR-142-R | GGCCATAATGGCCAGGCGGCCAGCACAGAACTCCTAC |
| GFP-F | ATGGTGAGCAAGGGCGA |
| GFP-R | CCGGTGGTGCAGATGAAC |
| v-rel-F | ATGGACTTTCTCACCAACCTCCG |
| v-rel-R | CGAACGATACCCGACTTG |
| HA | GGATGAGGTGACTAAGAAAG |
| envA | AGAGAAAGAGGGGCGTCTAAGGAGA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Y.; Zhang, Y.; Tang, N.; Pedrera, M.; Shen, Z.; Nair, V. Inhibition of v-rel-Induced Oncogenesis through microRNA Targeting. Viruses 2018, 10, 242. https://doi.org/10.3390/v10050242
Yao Y, Zhang Y, Tang N, Pedrera M, Shen Z, Nair V. Inhibition of v-rel-Induced Oncogenesis through microRNA Targeting. Viruses. 2018; 10(5):242. https://doi.org/10.3390/v10050242
Chicago/Turabian StyleYao, Yongxiu, Yaoyao Zhang, Na Tang, Miriam Pedrera, Zhiqiang Shen, and Venugopal Nair. 2018. "Inhibition of v-rel-Induced Oncogenesis through microRNA Targeting" Viruses 10, no. 5: 242. https://doi.org/10.3390/v10050242
APA StyleYao, Y., Zhang, Y., Tang, N., Pedrera, M., Shen, Z., & Nair, V. (2018). Inhibition of v-rel-Induced Oncogenesis through microRNA Targeting. Viruses, 10(5), 242. https://doi.org/10.3390/v10050242