Saccharomyces paradoxus K66 Killer System Evidences Expanded Assortment of Helper and Satellite Viruses

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Media
2.2. Assay for Killing/Resistance Phenotypes
2.3. Viral dsRNA Isolation From Yeast
2.4. cDNA Synthesis, Amplification, and Cloning
2.5. Sequence Analysis
2.6. Analysis of the Partially Purified K66 Toxin for Thermal and pH Activity
2.7. Screening for Modulators of K66 Activity and Bioinformatic Analysis
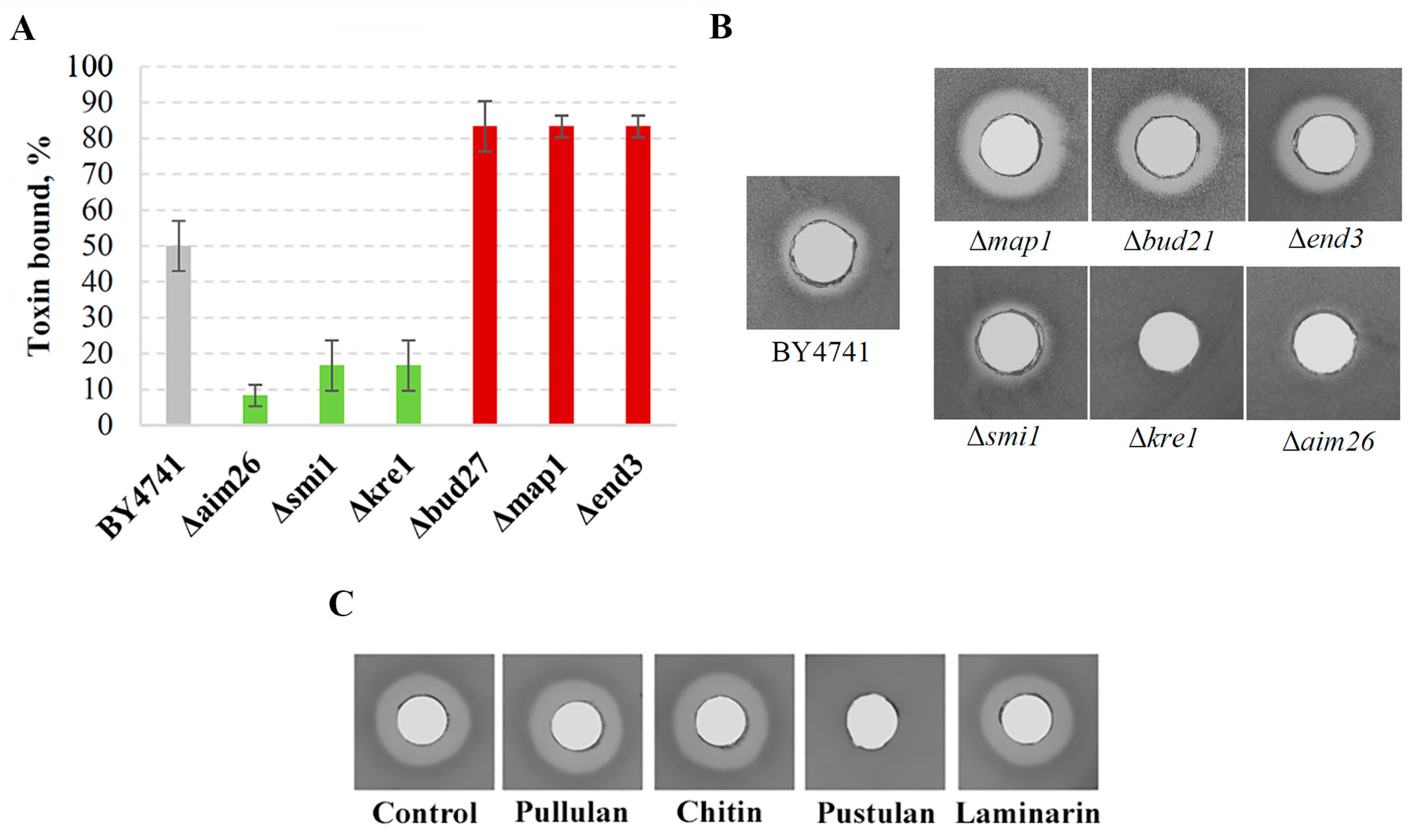
2.8. Evaluation of K66 Toxin Binding to the Yeast Mutants and Different Polysaccharides
2.9. GenBank Accession Numbers
3. Results
3.1. Characterization of the S. paradoxus Killer Strain
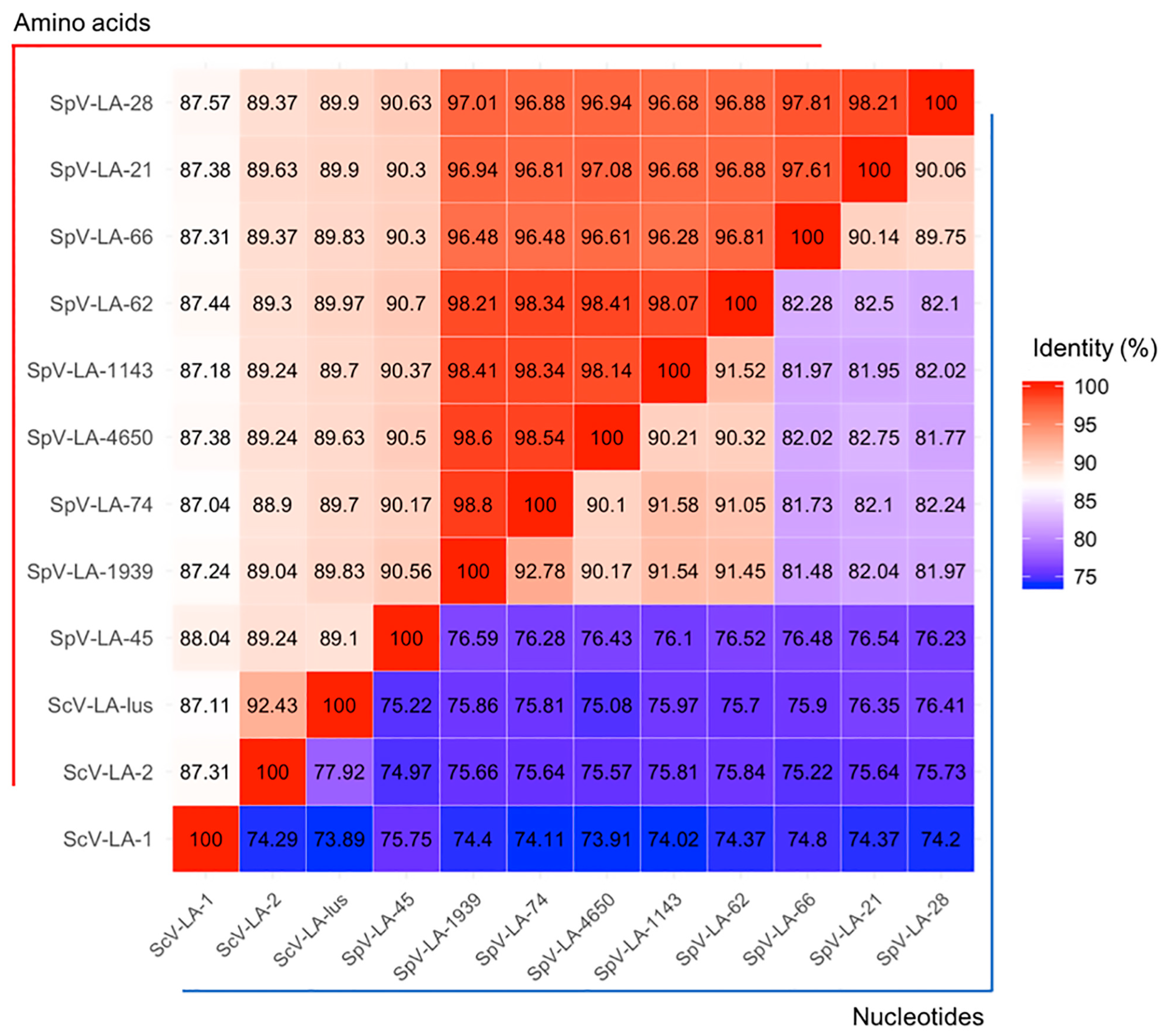
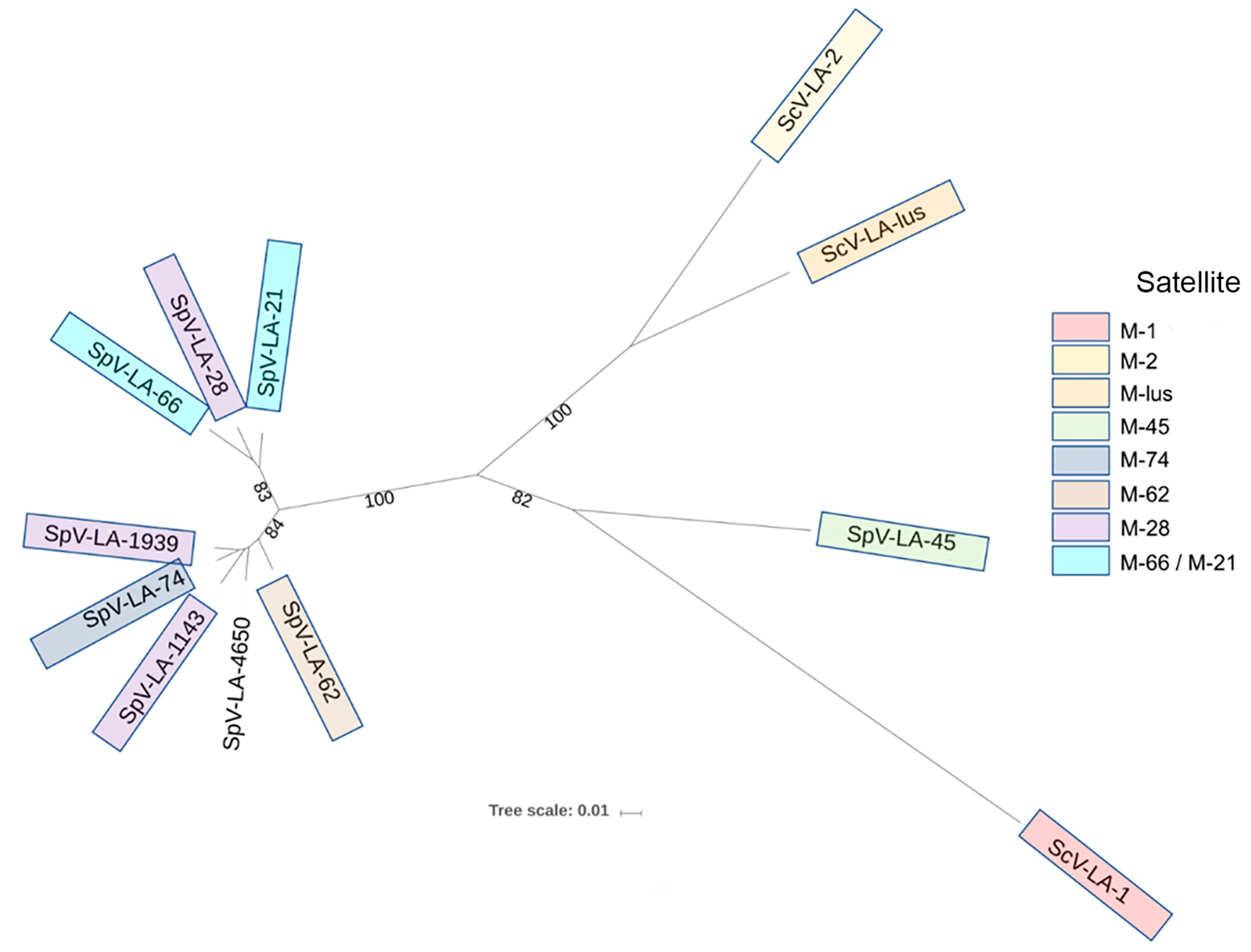
3.2. Double-Stranded RNA Viruses from the S. Paradoxus Strain
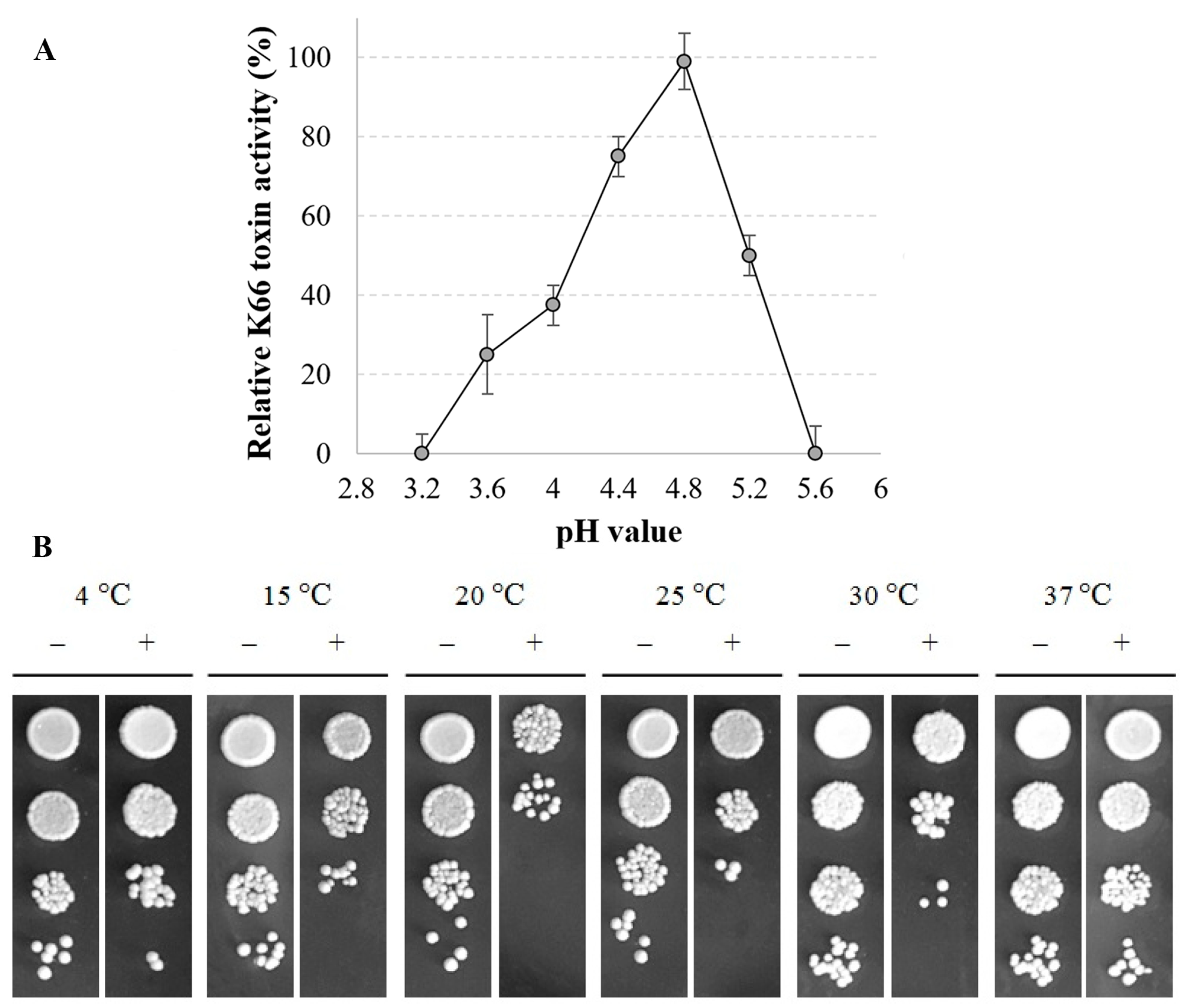
3.3. Effect of pH and Temperature on the Action of the S. Paradoxus K66 Toxin
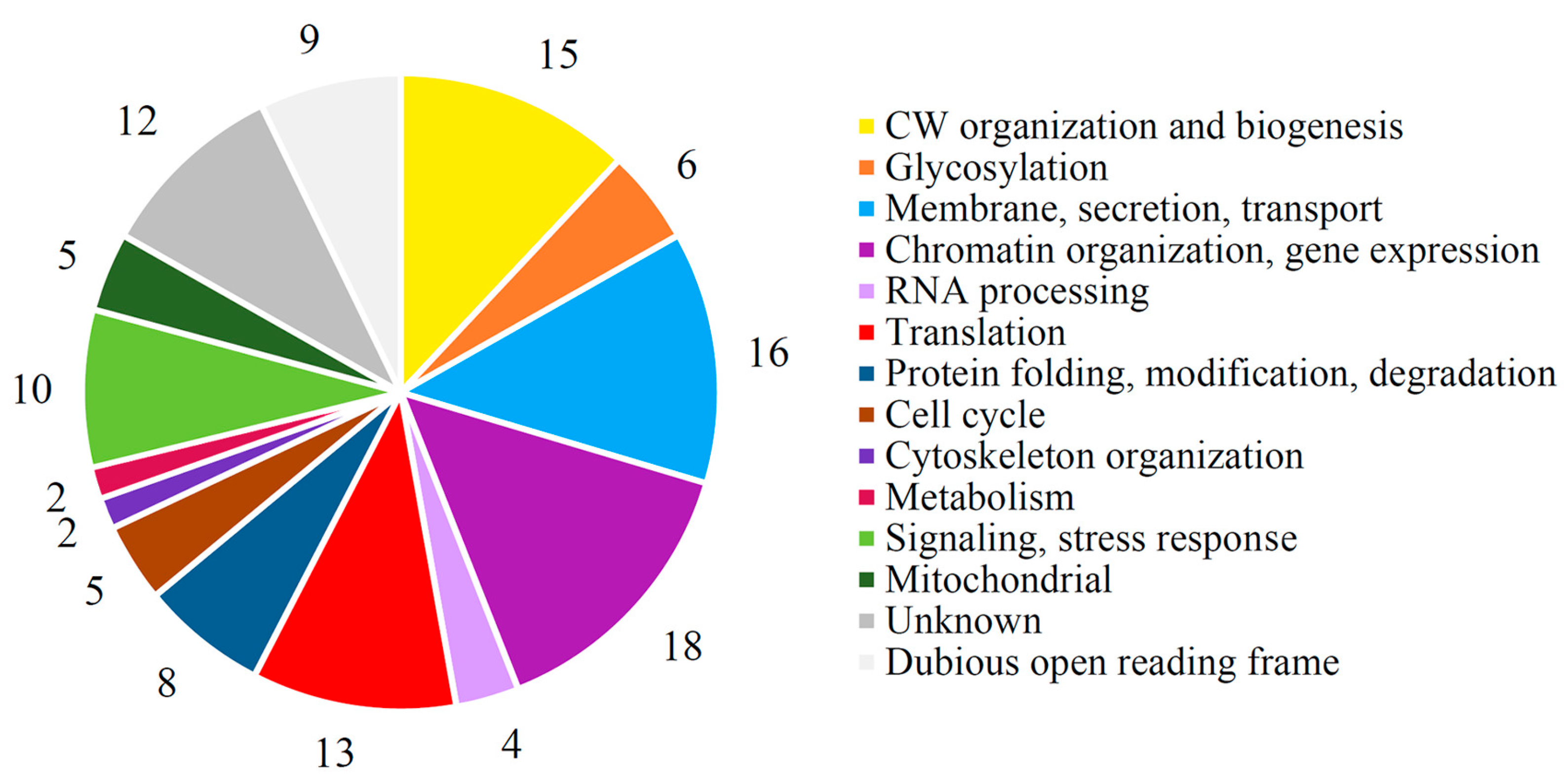
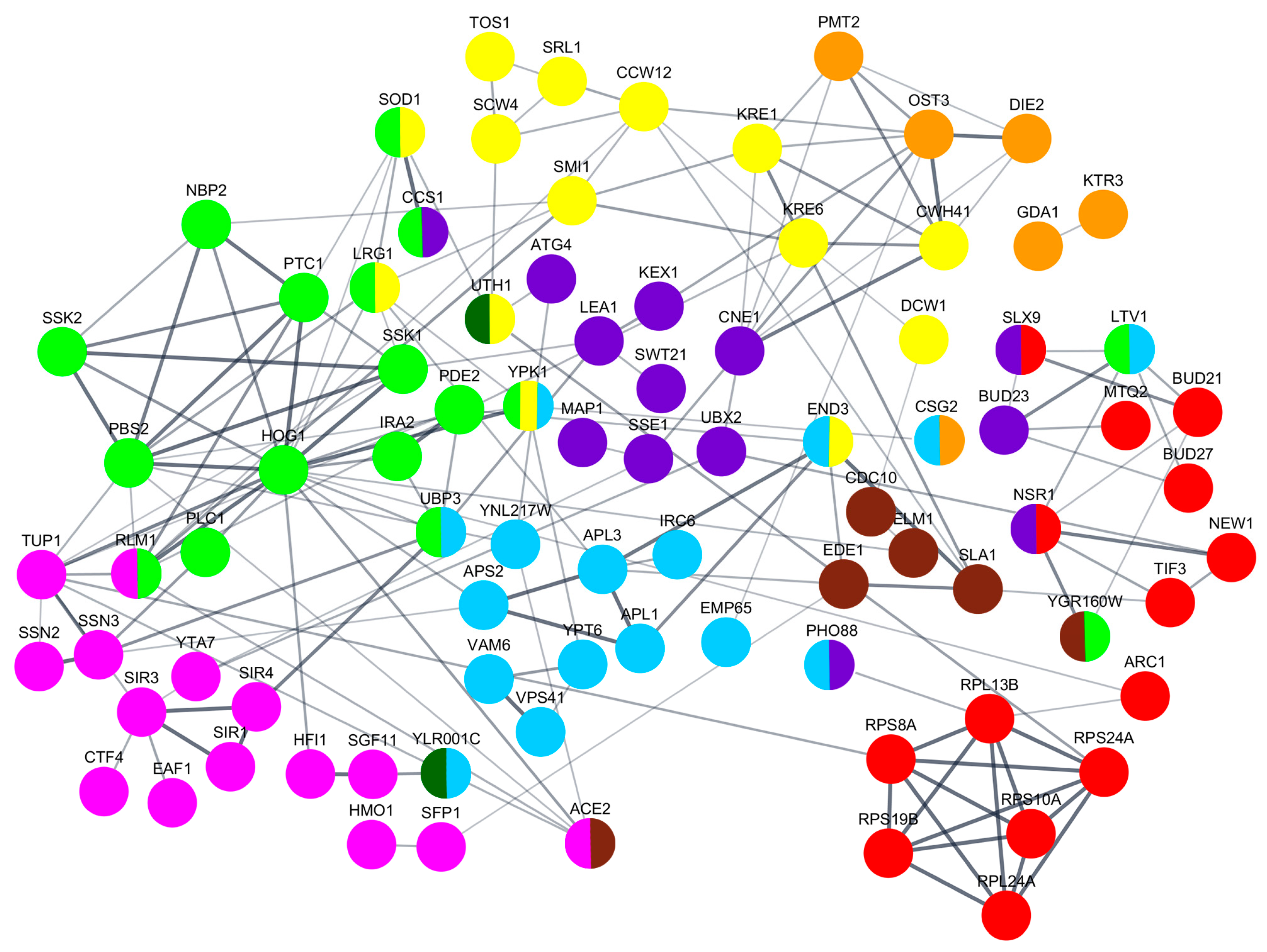
3.4. Genetic Factors Modulating the Functionality of the Viral K66 Toxin
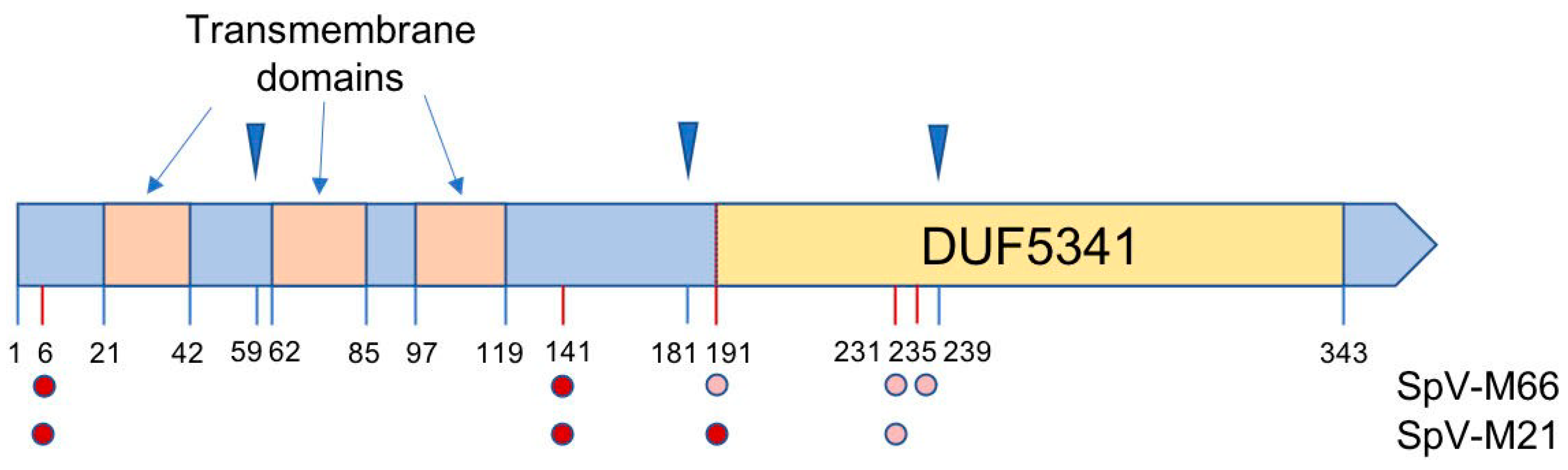
3.5. Targeting of the Viral K66 Killer Protein to the Cell Wall
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Muccilli, S.; Restuccia, C. Bioprotective role of yeasts. Microorganisms 2015, 3, 588–611. [Google Scholar] [CrossRef] [PubMed]
- Yap, N.A.; De Barros Lopes, M.; Langridge, P.; Henschke, P.A. The incidence of killer activity of non-Saccharomyces yeasts towards indigenous yeast species of grape must: Potential application in wine fermentation. J. Appl. Microbiol. 2000, 89, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Kast, A.; Voges, R.; Schroth, M.; Schaffrath, R.; Klassen, R.; Meinhardt, F. Autoselection of cytoplasmic yeast virus like elements encoding toxin/antitoxin systems involves a nuclear barrier for immunity gene expression. PLoS Genet. 2015, 11, e1005005. [Google Scholar] [CrossRef] [PubMed]
- Drinnenberg, I.A.; Fink, G.R.; Bartel, D.P. Compatibility with killer explains the rise of RNAi-deficient fungi. Science 2011, 333, 1592. [Google Scholar] [CrossRef] [PubMed]
- Golubev, V.I. Wine yeast races maintained in the All-Russia Collection of Microorganisms (VKM IBPM RAS). Prikl. Biokhim. Mikrobiol. 2005, 41, 592–595. [Google Scholar] [PubMed]
- Rodríguez-Cousiño, N.; Gómez, P.; Esteban, R. Variation and distribution of L-A helper totiviruses in Saccharomyces sensu stricto yeasts producing different killer toxins. Toxins (Basel) 2017, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.; Velázquez, R.; López-Piñeiro, A.; Naranjo, B.; Roig, F.; Llorens, C. New insights into the genome organization of yeast killer viruses based on “atypical” killer strains characterized by high-throughput sequencing. Toxins (Basel) 2017, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B. Prions and RNA viruses of Saccharomyces cerevisiae. Annu. Rev. Genet. 1996, 30, 109–139. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.L.; Leu, J.Y.; Chang, T.H. A population study of killer viruses reveals different evolutionary histories of two closely related Saccharomyces sensu stricto yeasts. Mol. Ecol. 2015, 24, 4312–4322. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Caston, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B.; Fujimura, T.; Esteban, R. Viruses and prions of Saccharomyces cerevisiae. Adv. Virus Res. 2013, 86, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B. Double-stranded RNA viruses of Saccharomyces cerevisiae. Microbiol. Rev. 1996, 60, 250–265. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.J.; Tipper, D.J. K28, a unique double-stranded RNA killer virus of Saccharomyces cerevisiae. Mol. Cell. Biol. 1990, 10, 4807–4815. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cousiño, N.; Gómez, P.; Esteban, R. L-A-lus, a new variant of the L-A totivirus found in wine yeasts with klus killer toxin-encoding mlus double-stranded RNA: Possible role of killer toxin-encoding satellite RNAs in the evolution of their helper viruses. Appl. Environ. Microbiol. 2013, 79, 4661–4674. [Google Scholar] [CrossRef] [PubMed]
- Schaffrath, R.; Meinhardt, F.; Klassen, R. Yeast killer toxins: Fundamentals and applications. In Physiology and Genetics, 2nd ed.; Anke, T., Schüffler, A., Eds.; Springer: Cham, Switzerland, 2018; pp. 87–118. ISBN 978-3-319-71739-5. [Google Scholar]
- Icho, T.; Wickner, R.B. The double-stranded RNA genome of yeast virus L-A encodes its own putative RNA polymerase by fusing two open reading frames. J. Biol. Chem. 1989, 264, 6716–6723. [Google Scholar] [PubMed]
- Dinman, J.D.; Icho, T.; Wickner, R.B. A-1 ribosomal frameshift in a double-stranded RNA virus of yeast forms a fag-pol fusion protein. Proc. Natl. Acad. Sci. USA 1991, 88, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Konovalovas, A.; Serviené, E.; Serva, S. Genome sequence of Saccharomyces cerevisiae double-stranded RNA virus L-A-28. Genome Announc. 2016, 4, e00549-16. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cousiño, N.; Esteban, R. Relationships and evolution of double stranded RNA totiviruses of yeasts inferred from analysis of L-A-2 and L-BC variants in wine yeast strain populations. Appl. Environ. Microbiol. 2017, 83, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B. Killer systems in Saccharomyces cerevisiae: Three distinct modes of exclusion of M2 double-stranded RNA by three species of double-stranded RNA, M1, L-A-E, and L-A-HN. Mol. Cell. Biol. 1983, 3, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Luksa, J.; Podoliankaite, M.; Vepstaite, I.; Strazdaite-Zieliene, Z.; Urbonavicius, J.; Serviene, E. Yeast beta-1,6-glucan is a primary target for the Saccharomyces cerevisiae K2 toxin. Eukaryot. Cell 2015, 14, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, K.; Bussey, H. Cell wall receptor for yeast killer toxin: Involvement of (1 leads to 6)-beta-D-glucan. J. Bacteriol. 1983, 154, 161–169. [Google Scholar] [PubMed]
- Breinig, F.; Tipper, D.J.; Schmitt, M.J. Kre1p, the plasma membrane receptor for the yeast K1 viral toxin. Cell 2002, 108, 395–405. [Google Scholar] [CrossRef]
- Orentaite, I.; Poranen, M.M.; Oksanen, H.M.; Daugelavicius, R.; Bamford, D.H. K2 killer toxin-induced physiological changes in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Martinac, B.; Zhu, H.; Kubalski, A.; Zhou, X.L.; Culbertson, M.; Bussey, H.; Kung, C. Yeast K1 killer toxin forms ion channels in sensitive yeast spheroplasts and in artificial liposomes. Proc. Natl. Acad. Sci. USA 1990, 87, 6228–6232. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.J.; Klavehn, P.; Wang, J.; Schonig, I.; Tipper, D.J. Cell cycle studies on the mode of action of yeast K28 killer toxin. Microbiology 1996, 142, 2655–2662. [Google Scholar] [CrossRef] [PubMed]
- Reiter, J.; Herker, E.; Madeo, F.; Schmitt, M.J. Viral killer toxins induce caspase-mediated apoptosis in yeast. J. Cell Biol. 2005, 168, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cousiño, N.; Maqueda, M.; Ambrona, J.; Zamora, E.; Esteban, R.; Ramírez, M. A new wine Saccharomyces cerevisiae killer toxin (Klus), encoded by a double-stranded RNA virus, with broad antifungal activity is evolutionarily related to a chromosomal host gene. Appl. Environ. Microbiol. 2011, 77, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Page, N.; Gerard-Vincent, M.; Menard, P.; Beaulieu, M.; Azuma, M.; Dijkgraaf, G.J.P.; Li, H.; Marcoux, J.; Nguyen, T.; Dowse, T.; et al. A Saccharomyces cerevisiae genome-wide mutant screen for altered sensitivity to K1 killer toxin. Genetics 2003, 163, 875–894. [Google Scholar] [PubMed]
- Carroll, S.Y.; Stirling, P.C.; Stimpson, H.E.M.; Giesselmann, E.; Schmitt, M.J.; Drubin, D.G. A yeast killer toxin screen provides insights into a/b toxin entry, trafficking, and killing mechanisms. Dev. Cell 2009, 17, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Serviene, E.; Luksa, J.; Orentaite, I.; Lafontaine, D.L.J.; Urbonavicius, J. Screening the budding yeast genome reveals unique factors affecting K2 toxin susceptibility. PLoS ONE 2012, 7, e50779. [Google Scholar] [CrossRef] [PubMed]
- Masison, D.C.; Blanc, A.; Ribas, J.C.; Carroll, K.; Sonenberg, N.; Wickner, R.B. Decoying the cap- mRNA degradation system by a double-stranded RNA virus and poly(A)- mRNA surveillance by a yeast antiviral system. Mol. Cell. Biol. 1995, 15, 2763–2771. [Google Scholar] [CrossRef] [PubMed]
- Rowley, P.A.; Ho, B.; Bushong, S.; Johnson, A.; Sawyer, S.L. XRN1 is a species-specific virus restriction factor in yeasts. PLoS Pathog. 2016, 12, e1005890. [Google Scholar] [CrossRef] [PubMed]
- Tercero, J.C.; Wickner, R.B. MAK3 encodes an N-acetyltransferase whose modification of the L-A gag NH2 terminus is necessary for virus particle assembly. J. Biol. Chem. 1992, 267, 20277–20281. [Google Scholar] [PubMed]
- Tercero, J.C.; Riles, L.E.; Wickner, R.B. Localized mutagenesis and evidence for post-transcriptional regulation of MAK3. A putative N-acetyltransferase required for double-stranded RNA virus propagation in Saccharomyces cerevisiae. J. Biol. Chem. 1992, 267, 20270–20276. [Google Scholar] [PubMed]
- Ohtake, Y.; Wickner, R.B. Yeast virus propagation depends critically on free 60S ribosomal subunit concentration. Mol. Cell. Biol. 1995, 15, 2772–2781. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.; Schmitt, M.J. Yeast killer toxin K28: Biology and unique strategy of host cell intoxication and killing. Toxins 2017, 9, 333. [Google Scholar] [CrossRef] [PubMed]
- McBride, R.C.; Boucher, N.; Park, D.S.; Turner, P.E.; Townsend, J.P. Yeast response to LA virus indicates coadapted global gene expression during mycoviral infection. FEMS Yeast Res. 2013, 13, 162–179. [Google Scholar] [CrossRef] [PubMed]
- Lukša, J.; Ravoitytė, B.; Konovalovas, A.; Aitmanaitė, L.; Butenko, A.; Yurchenko, V.; Serva, S.; Servienė, E. Different metabolic pathways are involved in response of Saccharomyces cerevisiae to L-A and M viruses. Toxins 2017, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- Göker, M.; Scheuner, C.; Klenk, H.-P.; Stielow, J.B.; Menzel, W. Codivergence of mycoviruses with their hosts. PLoS ONE 2011, 6, e22252. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Discovery of novel dsRNA viral sequences by in silico cloning and implications for viral diversity, host range and evolution. PLoS ONE 2012, 7, e42147. [Google Scholar] [CrossRef] [PubMed]
- Nakayashiki, T.; Kurtzman, C.P.; Edskes, H.K.; Wickner, R.B. Yeast prions [URE3] and [PSI+] are diseases. Proc. Natl. Acad. Sci. USA 2005, 102, 10575–10580. [Google Scholar] [CrossRef] [PubMed]
- Young, T.W.; Yagiu, M. A comparison of the killer character in different yeasts and its classification. Antonie Van Leeuwenhoek 1978, 44, 59–77. [Google Scholar] [CrossRef] [PubMed]
- Naumov, G.I.; Ivannikoiva, I.V.; Naumova, E.S. Molecular polymorphism of viral dsRNA of yeast Saccharomyces paradoxus. Mol. Gen. Mikrobiol. Virusol. 2005, 1, 38–40. [Google Scholar]
- Čitavičius, D.; Inge-Večtomov, S.G. Množestvennye mutanty u drožžej Saccharomyces cerevisiae-I. Polučenie i obščaja kharakteristika. Genetika 1972, 1, 95–102. [Google Scholar]
- Naumova, T.I.; Naumov, G.I. Sravnitel’naja genetika drožžej. Soobščenie XII. Izučenie antagonističeskikh otnošenij u drožžej roda Saccharomyces. Genetika 1973, 9, 85–90. [Google Scholar]
- Somers, J.M.; Bevan, E.A. The inheritance of the killer character in yeast. Genet. Res. 1969, 13, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Vepstaite-Monstavice, I.; Luksa, J.; Staneviciene, R.; Strazdaite-Zieliene, Z.; Yurchenko, V.; Serva, S.; Serviene, E. Distribution of apple and blackcurrant microbiota in Lithuania and the Czech Republic. Microbiol. Res. 2018, 206, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gulbiniene, G.; Kondratiene, L.; Jokantaite, T.; Serviene, E.; Melvydas, V.; Petkuniene, G. Occurrence of killer yeast strains in fruit and berry wine yeast populations. Food Technol. Biotechnol. 2004, 42, 159–163. [Google Scholar]
- Fried, H.M.; Fink, G.R. Electron microscopic heteroduplex analysis of “killer” double-stranded RNA species from yeast. Proc. Natl. Acad. Sci. USA 1978, 75, 4224–4228. [Google Scholar] [CrossRef] [PubMed]
- Grybchuk, D.; Akopyants, N.S.; Kostygov, A.Y.; Konovalovas, A.; Lye, L.F.; Dobson, D.E.; Zangger, H.; Fasel, N.; Butenko, A.; Frolov, A.O.; et al. Viral discovery and diversity in trypanosomatid protozoa with a focus on relatives of the human parasite Leishmania. Proc. Natl. Acad. Sci. USA 2018, 115, E506–E515. [Google Scholar] [CrossRef] [PubMed]
- Potgieter, A.C.; Page, N.A.; Liebenberg, J.; Wright, I.M.; Landt, O.; van Dijk, A.A. Improved strategies for sequence-independent amplification and sequencing of viral double-stranded RNA genomes. J. Gen. Virol. 2009, 90, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kall, L.; Krogh, A.; Sonnhammer, E.L.L. Advantages of combined transmembrane topology and signal peptide prediction--the Phobius web server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Ferre, F.; Clote, P. DiANNA: A web server for disulfide connectivity prediction. Nucleic Acids Res. 2005, 33, W230–W232. [Google Scholar] [CrossRef] [PubMed]
- Lukša, J.; Serva, S.; Serviene, E. Saccharomyces cerevisiae K2 toxin requires acidic environment for unidirectional folding into active state. Mycoscience 2016, 57, 51–57. [Google Scholar] [CrossRef]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 2005, 21, 3448–3449. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Kuhn, M.; Simonovic, M.; Roth, A.; Minguez, P.; Doerks, T.; Stark, M.; Muller, J.; Bork, P.; et al. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, D561–D568. [Google Scholar] [CrossRef] [PubMed]
- Novotna, D.; Flegelova, H.; Janderova, B. Different action of killer toxins K1 and K2 on the plasma membrane and the cell wall of Saccharomyces cerevisiae. FEMS Yeast Res. 2004, 4, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Naumov, G.I.; Ivannikova, I.V.; Chernov, I.I.; Naumova, E.S. Genetic polymorphism of double stranded RNA of Saccharomyces plasmids. Mikrobiologiia 2009, 78, 242–247. [Google Scholar] [PubMed]
- Dignard, D.; Whiteway, M.; Germain, D.; Tessier, D.; Thomas, D.Y. Expression in yeast of a cDNA copy of the K2 killer toxin gene. Production 1991, 7, 127–136. [Google Scholar] [CrossRef]
- Frank, A.C.; Wolfe, K.H. Evolutionary capture of viral and plasmid DNA by yeast nuclear chromosomes. Eukaryot. Cell 2009, 8, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, Y.; Jiang, D.; Li, G.; Xie, J.; Cheng, J.; Peng, Y.; Ghabrial, S.A.; Yi, X. Widespread horizontal gene transfer from double-stranded RNA viruses to eukaryotic nuclear genomes. J. Virol. 2010, 84, 11876–11887. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, F.; Klassen, R. Yeast killer toxins: Fundamentals and applications. In Physiology and Genetics. The Mycota (a Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Anke, T., Weber, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 15, pp. 107–130. ISBN 978-3-642-00286-1. [Google Scholar]
- Sun, H.; Ma, H.; Hao, M.; Pretorius, I.; Chen, S. Identification of yeast population dynamics of spontaneous fermentation in Beijing wine region, China. Ann. Mikrobiol. 2009, 59, 69–76. [Google Scholar] [CrossRef]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The microbial ecology of wine grape berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Mannazzu, I.; Clementi, F.; Ciani, M. Strategies and criteria for the isolation and selection of autochthonous starters. In Biodiversity and Biotechnology of Wine Yeasts; Ciani, M., Ed.; Research Signpost: Kerala, India, 2002; pp. 19–35. ISBN 978-8-177-36120. [Google Scholar]
- Podoliankaite, M.; Luksa, J.; Vysniauskis, G.; Sereikaite, J.; Melvydas, V.; Serva, S.; Serviene, E. High-yield expression in Escherichia coli, purification and application of budding yeast K2 killer protein. Mol. Biotechnol. 2014, 56, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Lebionka, A.; Servienë, E.; Melvydas, V. Isolation and purification of yeast Saccharomyces cerevisiae K2 killer toxin. Biologija 2002, 4, 2–4. [Google Scholar]
- Liu, G.L.; Chi, Z.; Wang, G.Y.; Wang, Z.P.; Li, Y.; Chi, Z.M. Yeast killer toxins, molecular mechanisms of their action and their applications. Crit. Rev. Biotechnol. 2015, 35, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; et al. Population genomics of domestic and wild yeasts. Nature 2009, 458, 337–341. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Strain (Killer Type) | Killing Phenotype of S. Paradoxus AML-15-66 | |||||||
|---|---|---|---|---|---|---|---|---|
| pH | ||||||||
| 3.2 | 3.6 | 4.0 | 4.4 | 4.8 | 5.2 | 5.6 | 6.0 | |
| S. cerevisiae ἀ 1 (K0) | - | +/- | + | ++ | +++ | ++/- | - | - |
| S. cerevisiae BY4741 (K0) | - | +/- | + | ++ | +++/- | + | - | - |
| S. cerevisiae K7 (K1) | - | - | +/- | + | + | +/- | - | - |
| S. cerevisiae K7 [L-M-] (K0) | - | - | +/- | + | ++/- | + | - | - |
| S. cerevisiae M437 (K2) | - | - | +/- | +/- | +/- | +/- | - | - |
| S. cerevisiae M437 [L-M-] (K0) | - | - | - | +/- | + | +/- | - | - |
| S. cerevisiae CRB-15-4 (Klus) | - | - | - | +/- | + | + | +/- | - |
| S. cerevisiae CRB-15-4 [L-M-] (K0) | - | - | - | +/- | + | + | +/- | - |
| S. cerevisiae MS300 (K28) | - | - | + | + | ++ | + | - | - |
| S. cerevisiae MS300 [L-M-] (K0) | - | - | + | ++/- | ++ | ++/- | +/- | - |
| S. paradoxus AML-15-66 (K66) | - | - | - | - | - | - | - | - |
| S. paradoxus AML-15-66 [L-M-] (K0) | - | - | +/- | + | ++/- | +/- | - | - |
| S. paradoxus T.21.4 (K21) | - | - | - | - | - | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vepštaitė-Monstavičė, I.; Lukša, J.; Konovalovas, A.; Ežerskytė, D.; Stanevičienė, R.; Strazdaitė-Žielienė, Ž.; Serva, S.; Servienė, E. Saccharomyces paradoxus K66 Killer System Evidences Expanded Assortment of Helper and Satellite Viruses. Viruses 2018, 10, 564. https://doi.org/10.3390/v10100564
Vepštaitė-Monstavičė I, Lukša J, Konovalovas A, Ežerskytė D, Stanevičienė R, Strazdaitė-Žielienė Ž, Serva S, Servienė E. Saccharomyces paradoxus K66 Killer System Evidences Expanded Assortment of Helper and Satellite Viruses. Viruses. 2018; 10(10):564. https://doi.org/10.3390/v10100564
Chicago/Turabian StyleVepštaitė-Monstavičė, Iglė, Juliana Lukša, Aleksandras Konovalovas, Dovilė Ežerskytė, Ramunė Stanevičienė, Živilė Strazdaitė-Žielienė, Saulius Serva, and Elena Servienė. 2018. "Saccharomyces paradoxus K66 Killer System Evidences Expanded Assortment of Helper and Satellite Viruses" Viruses 10, no. 10: 564. https://doi.org/10.3390/v10100564
APA StyleVepštaitė-Monstavičė, I., Lukša, J., Konovalovas, A., Ežerskytė, D., Stanevičienė, R., Strazdaitė-Žielienė, Ž., Serva, S., & Servienė, E. (2018). Saccharomyces paradoxus K66 Killer System Evidences Expanded Assortment of Helper and Satellite Viruses. Viruses, 10(10), 564. https://doi.org/10.3390/v10100564