Identification of Secreted Proteins Involved in Nonspecific dsRNA-Mediated Lutzomyia longipalpis LL5 Cell Antiviral Response

 , ,
, ,
Abstract
1. Introduction
2. Material and Methods
2.1. Cell Growth
2.2. Transfection and Conditioned Medium Collection
2.3. Trichloroacetic Acid Precipitation
2.4. Mass Spectrometry Analysis
2.5. In Silico Analyses
2.6. RNA Extraction and cDNA Synthesis
2.7. qPCR
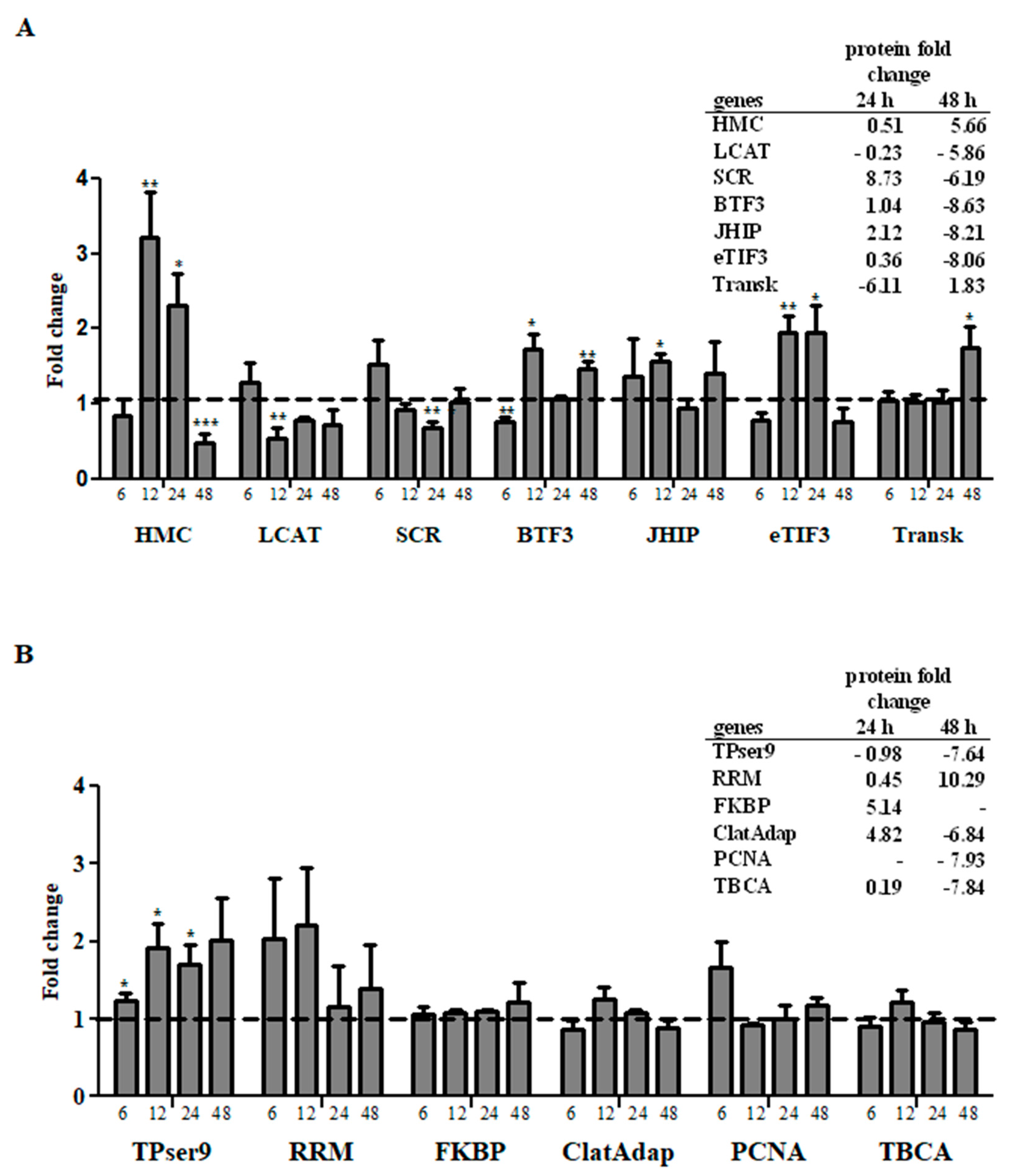
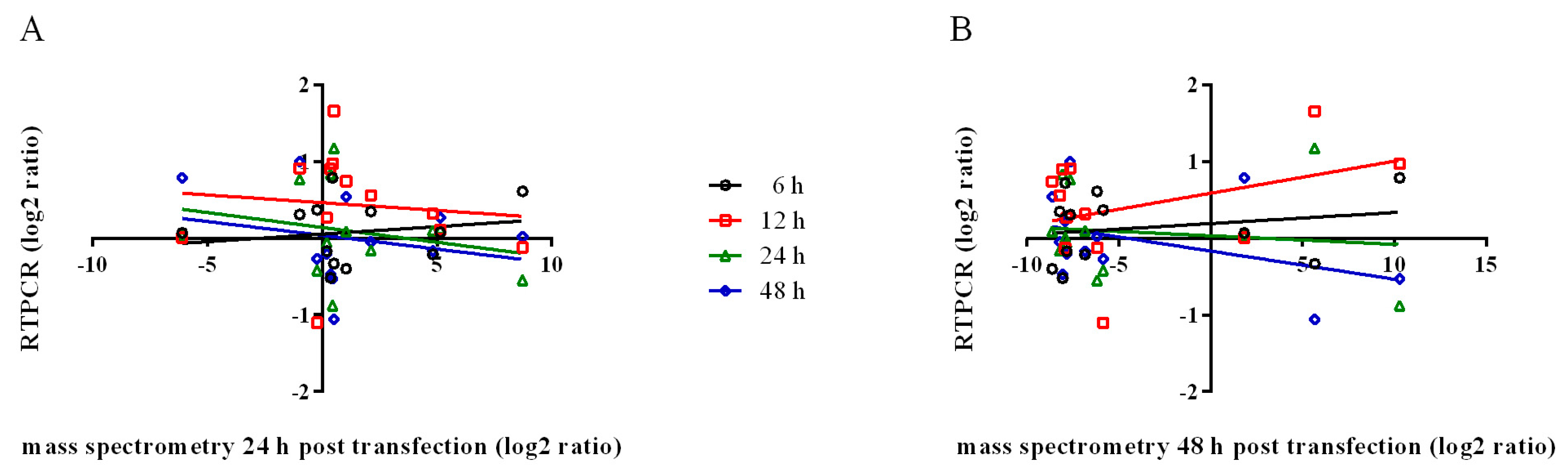
2.8. Comparative Analysis of Transcript and Protein Levels
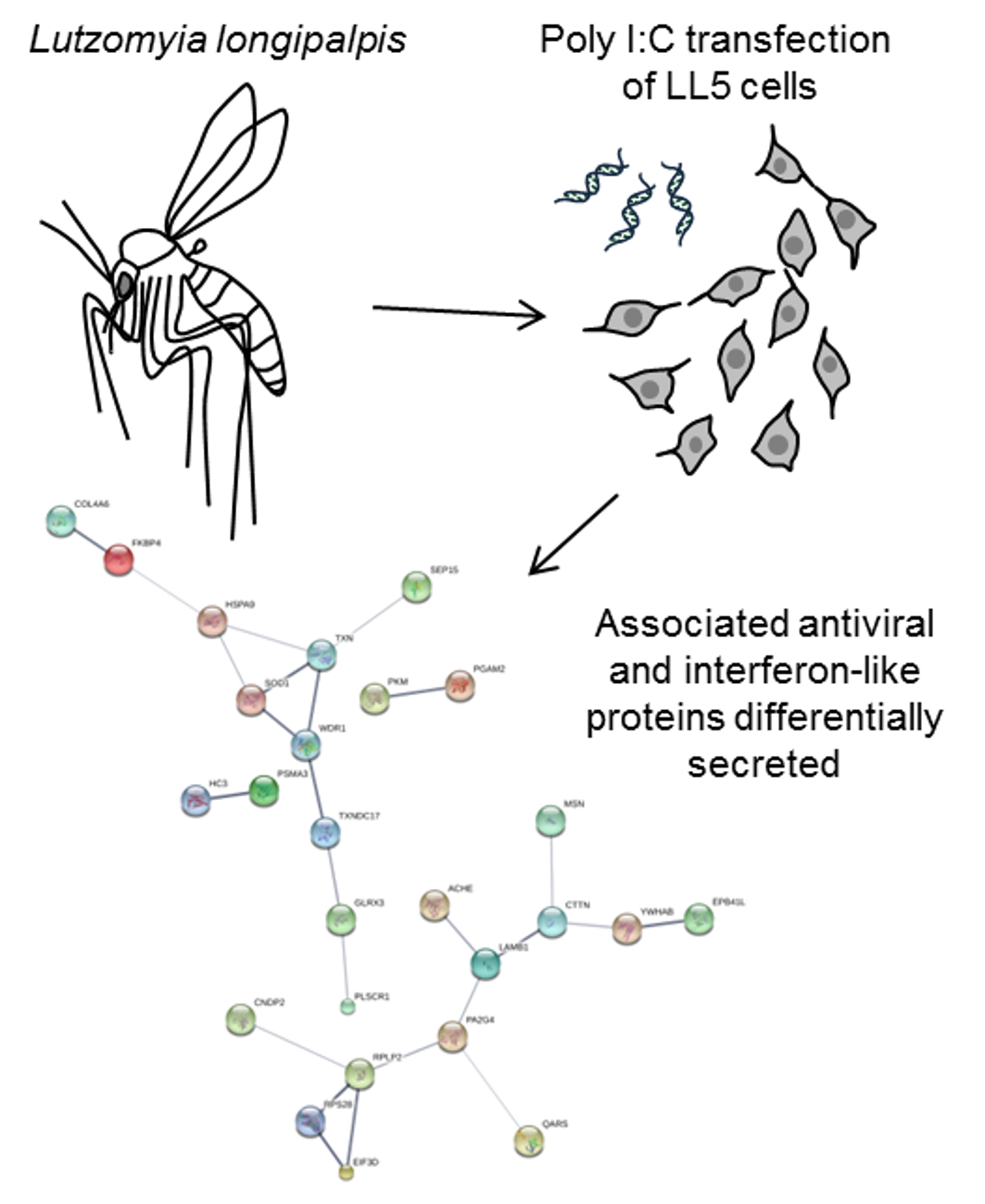
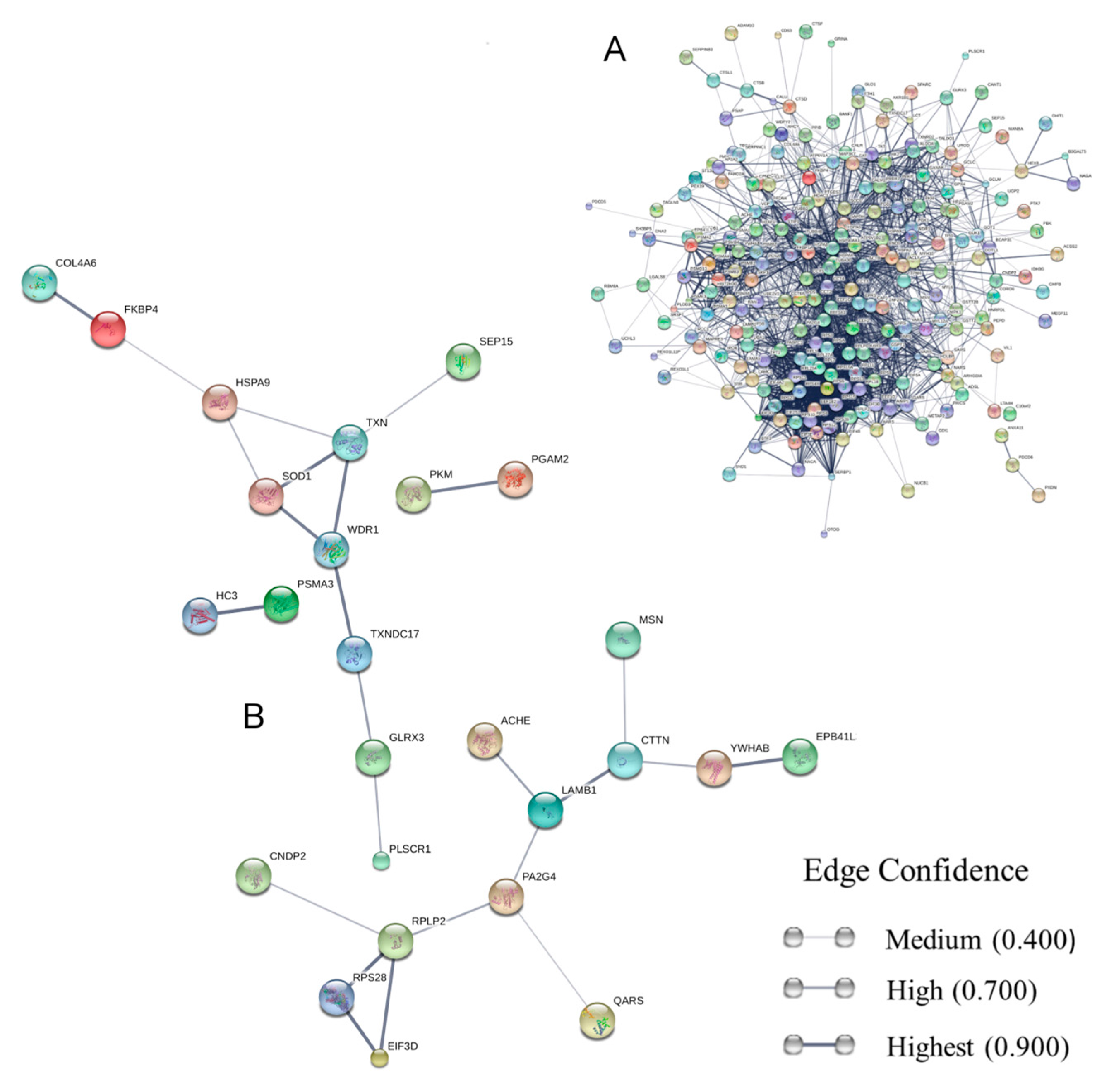
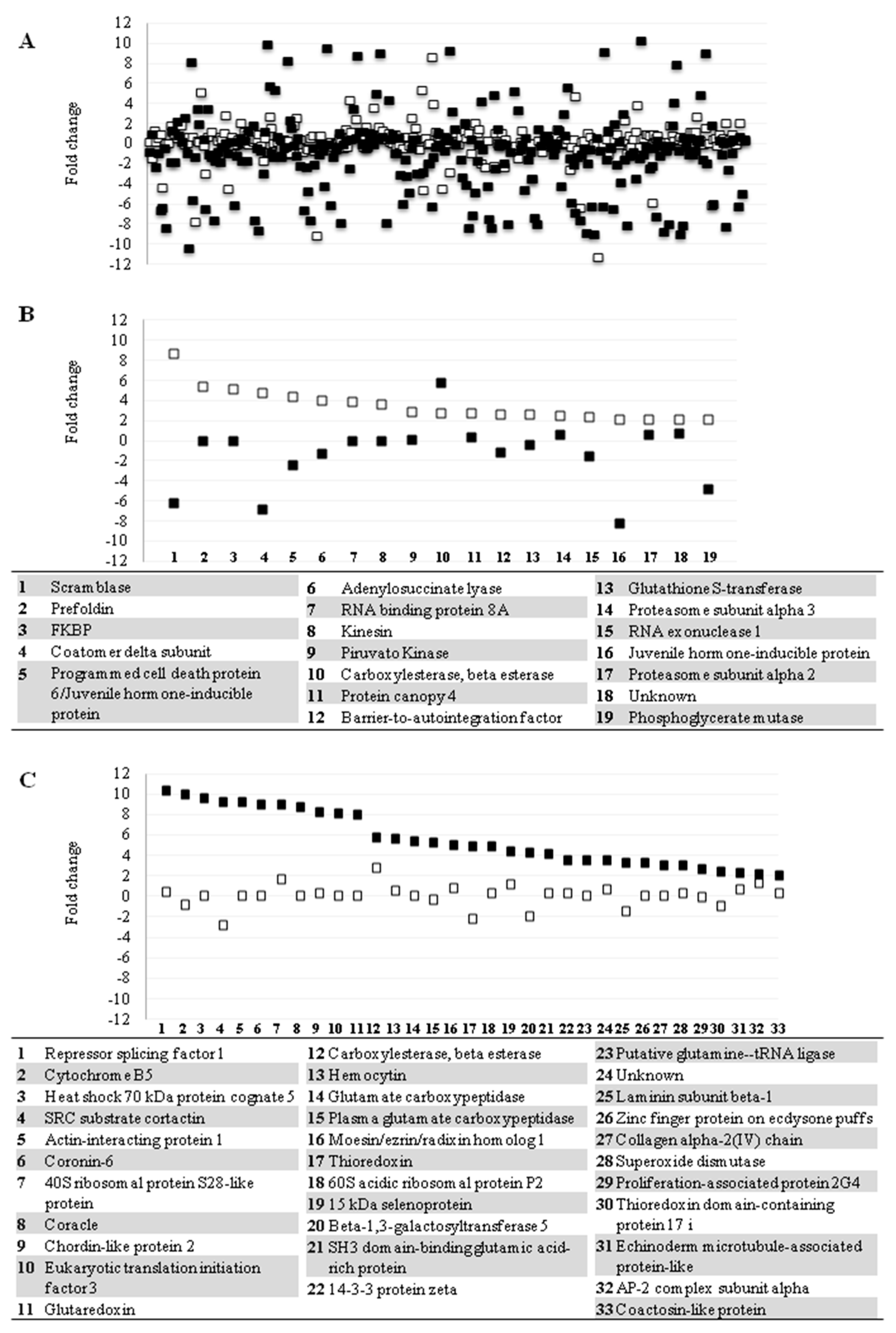
3. Results and Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ready, P.D. Biology of Phlebotomine Sand Flies as Vectors of Disease Agents. Annu. Rev. Entomol. 2013, 58, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Jeffries, C.L.; Walker, T. Biological Control of Mosquito Vectors: Past, Present, and Future. Insects 2016, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.N.; Rodrigues, F.M. Chandipura: A New Arbovirus Isolated in India from Patients with Febrile Illness. Indian J. Med. Res. 1967, 55, 1295–1305. [Google Scholar] [PubMed]
- Dhanda, V.; Rodrigues, F.M.; Ghosh, S.N. Isolation of Chandipura Virus from Sandflies in Aurangabad. Indian J. Med. Res. 1970, 58, 179–180. [Google Scholar] [PubMed]
- Tesh, R.B. The Genus Phlebovirus and Its Vectors. Annu. Rev. Entomol. 1988, 33, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Travassos da Rosa, A.P.; Tesh, R.B.; da Rosa, J.F.T.; Herve, J.P.; Main, A.J., Jr. Carajas and Maraba Viruses, Two New V esiculoviruses Isolated from Phlebotomine Sand Flies in Brazil. Am. J. Trop. Med. Hyg. 1984, 33, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Duboise, S.M. Viremia and Immune Response with Sequential Phlebovirus Infections. Am. J. Trop. Med. Hyg. 1987, 36, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Zapata, S.; Bichaud, L.; Moureau, G.; Lemey, P.; Firth, A.E.; Gritsun, T.S.; Gould, E.A.; de Lamballerie, X.; Depaquit, J.; et al. Ecuador Paraiso Escondido Virus, a New Flavivirus Isolated from New World Sand Flies in Ecuador, Is the First Representative of a Novel Clade in the Genus Flavivirus. J. Virol. 2015, 89, 11773–11785. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Wiley, M.R.; da Rosa, A.P.T.; Guzman, H.; Quiroz, E.; Savji, N.; Carrera, J.P.; Bussetti, A.V.; Ladner, J.T.; Lipkin, W.I.; et al. Characterization of the Punta Toro Species Complex (Genus Phlebovirus, Family Bunyaviridae). J. Gen. Virol. 2015, 96, 2079–2085. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Gao, X.; Gould, E.A. Factors Responsible for the Emergence of Arboviruses; Strategies, Challenges and Limitations for Their Control. Emerg. Microbes Infect. 2015, 4, e18. [Google Scholar] [CrossRef] [PubMed]
- Saiz, J.C.; Vázquez-Calvo, Á.; Blázquez, A.B.; Merino-Ramos, T.; Escribano-Romero, E.; Martín-Acebes, M.A. Zika Virus: The Latest Newcomer. Front. Microbiol. 2016, 7, 496. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, P.; Damsky, W.E., Jr.; Giordano, R.; Birungi, J.; Munstermann, L.E.; Conn, J.E. Infection of New- and Old-World Aedes Albopictus (Diptera: Culicidae) by the Intracellular Parasite Wolbachia: Implications for Host Mitochondrial DNA Evolution. J. Med. Entomol. 2003, 40, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R.; Ward, R.D.; Shaw, J.J. Experimental Transmission of Leishmania Chagasi, Causative Agent of Neotropical Visceral Leishmaniasis, by the Sandfly Lutzomyia longipalpis. Nature 1977, 266, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Telleria, E.L.; Sant’Anna, M.R.; Ortigão-Farias, J.R.; Pitaluga, A.N.; Dillon, V.M.; Bates, P.A.; Traub-Csekö, Y.M.; Dillon, R.J. Caspar-Like Gene Depletion Reduces Leishmania Infection in Sand Fly Host Lutzomyia longipalpis. J. Biol. Chem. 2012, 287, 12985–12993. [Google Scholar] [CrossRef] [PubMed]
- Telleria, E.L.; Sant’Anna, M.R.V.; Alkurbi, M.O.; Pitaluga, A.N.; Dillon, R.J.; Traub-Csekö, Y.M. Bacterial Feeding, Leishmania Infection and Distinct Infection Routes Induce Differential Defensin Expression in Lutzomyia longipalpis. Parasit Vectors 2013, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Di-Blasi, T.; Lobo, A.R.; Nascimento, L.M.; Córdova-Rojas, J.L.; Pestana, K.; Marín-Villa, M.; Tempone, A.J.; Telleria, E.L.; Ramalho-Ortigao, M.; McMahon-Pratt, D.; et al. The Flagellar Protein Flag1/Smp1 Is a Candidate for Leishmania-Sand Fly Interaction. Vector Borne Zoonotic Dis. 2015, 15, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, T.; Kobayashi, M.; Eshita, Y.; Shirato, K.; Kimura, T.; Ako, Y.; Miyoshi, H.; Takasaki, T.; Kurane, I.; Kariwa, H.; et al. Involvement of the Jnk-Like Protein of the Aedes Albopictus Mosquito Cell Line, C6/36, in Phagocytosis, Endocytosis and Infection of West Nile Virus. Insect Mol. Biol. 2003, 12, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vargas, I.; Travanty, E.A.; Keene, K.M.; Franz, A.W.; Beaty, B.J.; Blair, C.D.; Olson, K.E. RNA Interference, Arthropod-Borne Viruses, and Mosquitoes. Virus Res. 2004, 102, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Rasgon, J.L.; Ren, X.; Petridis, M. Can Anopheles Gambiae Be Infected with Wolbachia Pipientis? Insights from an in Vitro System. Appl. Environ. Microbiol. 2006, 72, 7718–7722. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Modi, G.B. Development of a Continuous Cell Line from the Sand Fly Lutzomyia longipalpis (Diptera: Psychodidae), and Its Susceptibility to Infection with Arboviruses. J. Med. Entomol. 1983, 20, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Tinoco-Nunes, B.; Telleria, E.L.; da Silva-Neves, M.; Marques, C.; Azevedo-Brito, D.A.; Pitaluga, A.N.; Traub-Csekö, Y.M. The Sandfly Lutzomyia longipalpis Ll5 Embryonic Cell Line Has Active Toll and Imd Pathways and Shows Immune Responses to Bacteria, Yeast and Leishmania. Parasit Vectors 2016, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Pitaluga, A.N.; Mason, P.W.; Traub-Cseko, Y.M. Non-Specific Antiviral Response Detected in RNA-Treated Cultured Cells of the Sandfly, Lutzomyia longipalpis. Dev. Comp. Immunol. 2008, 32, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.E.; Garcia-Sastre, A. The Virus Battles: IFN Induction of the Antiviral State and Mechanisms of Viral Evasion. Cytokine Growth Factor Rev. 2001, 12, 143–156. [Google Scholar] [CrossRef]
- Fayzulin, R.; Scholle, F.; Petrakova, O.; Frolov, I.; Mason, P.W. Evaluation of Replicative Capacity and Genetic Stability of West Nile Virus Replicons Using Highly Efficient Packaging Cell Lines. Virology 2006, 351, 196–209. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.K.; Jwa, N.S.; Lebrun, M.H.; Job, D.; Rakwal, R. Plant Secretome: Unlocking Secrets of the Secreted Proteins. Proteomics 2010, 10, 799–827. [Google Scholar] [CrossRef] [PubMed]
- Assil, S.; Webster, B.; Dreux, M. Regulation of the Host Antiviral State by Intercellular Communications. Viruses 2015, 7, 4707–4733. [Google Scholar] [CrossRef] [PubMed]
- Watanabe Costa, R.; da Silveira, J.F.; Bahia, D. Interactions between Trypanosoma Cruzi Secreted Proteins and Host Cell Signaling Pathways. Front. Microbiol. 2016, 7, 388. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.V.; de Godoy, L.M.; Li, G.; Macek, B.; Mortensen, P.; Pesch, R.; Makarov, A.; Lange, O.; Horning, S.; Mann, M. Parts Per Million Mass Accuracy on an Orbitrap Mass Spectrometer Via Lock Mass Injection into a C-Trap. Mol. Cell. Proteom. 2005, 4, 2010–2021. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. Maxquant Enables High Peptide Identification Rates, Individualized P.P.B.-Range Mass Accuracies and Proteome-Wide Protein Quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A Peptide Search Engine Integrated into the Maxquant Environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Luber, C.A.; Cox, J.; Lauterbach, H.; Fancke, B.; Selbach, M.; Tschopp, J.; Akira, S.; Wiegand, M.; Hochrein, H.; O’keeffe, M.; et al. Quantitative Proteomics Reveals Subset-Specific Viral Recognition in Dendritic Cells. Immunity 2010, 32, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Hiller, K.; Grote, A.; Scheer, M.; Münch, R.; Jahn, D. Predisi: Prediction of Signal Peptides and Their Cleavage Positions. Nucleic Acids Res. 2004, 32, W375–W379. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. String V10: Protein-Protein Interaction Networks, Integrated over the Tree of Life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Schefe, J.H.; Lehmann, K.E.; Buschmann, I.R.; Unger, T.; Funke-Kaiser, H. Quantitative Real-Time RT-PCR Data Analysis: Current Concepts and the Novel “Gene Expression’s CT Difference” Formula. J. Mol. Med. 2006, 84, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Meireles, A.C.A.; Amoretty, P.R.; Souza, N.A.; Kyriacou, C.P.; Peixoto, A.A. Rhythmic Expression of the Cycle Gene in a Hematophagous Insect Vector. BMC Mol. Biol. 2006, 7, 38. [Google Scholar]
- Lemaitre, B.; Kromer-Metzger, E.; Michaut, L.; Nicolas, E.; Meister, M.; Georgel, P.; Reichhart, J.M.; Hoffmann, J.A. A Recessive Mutation, Immune Deficiency (IMD), Defines Two Distinct Control Pathways in the Drosophila Host Defense. Proc. Natl. Acad. Sci. USA 1995, 92, 9465–9469. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.A.; Anderson, K.V. Drosophila: The Genetics of Innate Immune Recognition and Response. Annu. Rev. Immunol. 2004, 22, 457–483. [Google Scholar] [CrossRef] [PubMed]
- Blair, C.D. Mosquito Rnai Is the Major Innate Immune Pathway Controlling Arbovirus Infection and Transmission. Future Microbiol. 2011, 6, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Zeidler, M.P.; Bausek, N. The Drosophila Jak-Stat Pathway. JAKSTAT 2013, 2, e25353. [Google Scholar] [PubMed]
- Cerutti, H.; Casas-Mollano, J.A. On the Origin and Functions of RNA-Mediated Silencing: From Protists to Man. Curr. Genet. 2006, 50, 81–99. [Google Scholar] [CrossRef] [PubMed]
- Robalino, J.; Bartlett, T.C.; Chapman, R.W.; Gross, P.S.; Browdy, C.L.; Warr, G.W. Double-Stranded RNA and Antiviral Immunity in Marine Shrimp: Inducible Host Mechanisms and Evidence for the Evolution of Viral Counter-Responses. Dev. Comp. Immunol. 2007, 31, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Robalino, J.; Carnegie, R.B.; Nuala, O.L.; Ouvry-Patat, S.A.; de la Vega, E.; Prior, S.; Gross, P.S.; Browdy, C.L.; Chapman, R.W.; Schey, K.L.; et al. Contributions of Functional Genomics and Proteomics to the Study of Immune Responses in the Pacific White Leg Shrimp Litopenaeus Vannamei. Vet. Immunol. Immunopathol. 2009, 128, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Flenniken, M.L.; Andino, R. Non-Specific Dsrna-Mediated Antiviral Response in the Honey Bee. PLoS ONE 2013, 8, e77263. [Google Scholar] [CrossRef] [PubMed]
- Piot, N.; Snoeck, S.; Vanlede, M.; Smagghe, G.; Meeus, I. The Effect of Oral Administration of Dsrna on Viral Replication and Mortality in Bombus Terrestris. Viruses 2015, 7, 3172–3185. [Google Scholar] [CrossRef] [PubMed]
- Brutscher, L.M.; Daughenbaugh, K.F.; Flenniken, M.L. Virus and dsRNA-triggered transcriptional responses reveal key components of honey bee antiviral defense. Sci. Rep. 2017, 7, 6448. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.C.; Tassetto, M.; van Rij, R.P.; Goic, B.; Gausson, V.; Berry, B.; Jacquier, C.; Antoniewski, C.; Andino, R. Antiviral Immunity in Drosophila Requires Systemic RNA Interference Spread. Nature 2009, 458, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA Helicase Rig-I Has an Essential Function in Double-Stranded RNA-Induced Innate Antiviral Responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Matsumoto, K.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Foy, E.; Loo, Y.M.; Gale, M.; Akira, S.; et al. Shared and Unique Functions of the Dexd/H-Box Helicases RIG-I, MDA5, and LGP2 in Antiviral Innate Immunity. J. Immunol. 2005, 175, 2851–2858. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Pathogen Recognition with Toll-Like Receptors. Curr. Opin. Immunol. 2005, 17, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Seth, R.B.; Sun, L.; Ea, C.K.; Chen, Z.J. Identification and Characterization of MAVS, a Mitochondrial Antiviral Signaling Protein That Activates NF-κB and IRF 3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Seya, T. TLR3: Interferon Induction by Double-Stranded RNA Including Poly(I:C). Adv. Drug Deliv. Rev. 2008, 60, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Levine, S.J. Toll-Like Receptor, RIG-I-Like Receptors and the Nlrp3 Inflammasome: Key Modulators of Innate Immune Responses to Double-Stranded RNA Viruses. Cytokine Growth Factor Rev. 2011, 22, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Nickel, W. Unconventional Secretory Routes: Direct Protein Export across the Plasma Membrane of Mammalian Cells. Traffic 2005, 6, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.A.; Akram, W. Cyromazine Resistance in a Field Strain of House Flies, Musca domestica L.: Resistance Risk Assessment and Bio-Chemical Mechanism. Chemosphere 2016, 167, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhao, J.; Stout, J.G.; Luhm, R.A.; Wiedmer, T.; Sims, P.J. Molecular Cloning of Human Plasma Membrane Phospholipid Scramblase. A Protein Mediating Transbilayer Movement of Plasma Membrane Phospholipids. J. Biol. Chem. 1997, 272, 18240–18244. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Zhou, Q.; Zhao, J.; Zhou, A.; Harty, R.N.; Bose, S.; Banerjee, A.; Slee, R.; Guenther, J.; Williams, B.R.; et al. Phospholipid Scramblase 1 Potentiates the Antiviral Activity of Interferon. J. Virol. 2004, 78, 8983–8993. [Google Scholar] [CrossRef] [PubMed]
- Talukder, A.H.; Bao, M.; Kim, T.W.; Facchinetti, V.; Hanabuchi, S.; Bover, L.; Zal, T.; Liu, Y.J. Phospholipid Scramblase 1 Regulates Toll-Like Receptor 9-Mediated Type I Interferon Production in Plasmacytoid Dendritic Cells. Cell Res. 2012, 22, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Kusano, S.; Eizuru, Y. Human Phospholipid Scramblase 1 Interacts with and Regulates Transactivation of Htlv-1 Tax. Virology 2012, 432, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Bartel, S.; Doellinger, J.; Darsow, K.; Bourquain, D.; Buchholz, R.; Nitsche, A.; Lange, H.A. Proteome Analysis of Vaccinia Virus IHD-W-Infected HEK 293 Cells with 2-Dimensional Gel Electrophoresis and MALDI-PSD-TOF MS of on Solid Phase Support N-Terminally Sulfonated Peptides. Virol. J. 2011, 8, 380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Han, Q.; Song, Y.; Chen, Q.; Xia, X. Analysis of Subcellular Prefoldin 1 Redistribution during Rabies Virus Infection. Jundishapur J. Microbiol. 2015, 8, e24757. [Google Scholar] [CrossRef] [PubMed]
- Tsao, M.L.; Chao, C.H.; Yeh, C.T. Interaction of Hepatitis C Virus F Protein with Prefoldin 2 Perturbs Tubulin Cytoskeleton Organization. Biochem. Biophys. Res. Commun. 2006, 348, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Bierer, B.E.; Mattila, P.S.; Standaert, R.F.; Herzenberg, L.A.; Burakoff, S.J.; Crabtree, G.; Schreiber, S.L. Two Distinct Signal Transmission Pathways in T Lymphocytes Are Inhibited by Complexes Formed between an Immunophilin and Either Fk506 or Rapamycin. Proc. Natl. Acad. Sci. USA 1990, 87, 9231–9235. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Farmer, J.D.; Lane, W.S.; Friedman, J.; Weissman, I.; Schreiber, S.L. Calcineurin Is a Common Target of Cyclophilin-Cyclosporin a and FKBP-FK506 Complexes. Cell 1991, 66, 807–815. [Google Scholar] [CrossRef]
- Akiyama, T.; Shiraishi, T.; Qin, J.; Konno, H.; Akiyama, N.; Shinzawa, M.; Miyauchi, M.; Takizawa, N.; Yanai, H.; Ohashi, H.; et al. Mitochondria-Nucleus Shuttling FK506-Binding Protein 51 Interacts with Traf Proteins and Facilitates the RIG-I-Like Receptor-Mediated Expression of Type I IFN. PLoS ONE 2014, 9, e95992. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Goldberg, J. Structure of Coatomer Cage Proteins and the Relationship among COPI, COPII, and Clathrin Vesicle Coats. Cell 2010, 142, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Gabriely, G.; Kama, R.; Gerst, J.E. Involvement of Specific Copi Subunits in Protein Sorting from the Late Endosome to the Vacuole in Yeast. Mol. Cell. Biol. 2007, 27, 526–540. [Google Scholar] [CrossRef] [PubMed]
- Razi, M.; Chan, E.Y.; Tooze, S.A. Early Endosomes and Endosomal Coatomer Are Required for Autophagy. J. Cell. Biol. 2009, 185, 305–321. [Google Scholar] [CrossRef] [PubMed]
- Misselwitz, B.; Dilling, S.; Vonaesch, P.; Sacher, R.; Snijder, B.; Schlumberger, M.; Rout, S.; Stark, M.; von Mering, C.; Pelkmans, L.; et al. RNAi Screen of Salmonella Invasion Shows Role of COPI in Membrane Targeting of Cholesterol and Cdc42. Mol. Syst. Biol. 2011, 7, 474. [Google Scholar] [CrossRef] [PubMed]
- Cherry, S.; Kunte, A.; Wang, H.; Coyne, C.; Rawson, R.B.; Perrimon, N. COPI Activity Coupled with Fatty Acid Biosynthesis Is Required for Viral Replication. PLoS Pathog. 2006, 2, e102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lee, S.Y.; Beznoussenko, G.V.; Peters, P.J.; Yang, J.S.; Gilbert, H.Y.; Brass, A.L.; Elledge, S.J.; Isaacs, S.N.; Moss, B.; et al. A Role for the Host Coatomer and Kdel Receptor in Early Vaccinia Biogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 163–168. [Google Scholar] [CrossRef] [PubMed]
- König, R.; Stertz, S.; Zhou, Y.; Inoue, A.; Hoffmann, H.H.; Bhattacharyya, S.; Alamares, J.G.; Tscherne, D.M.; Ortigoza, M.B.; Liang, Y.; et al. Human Host Factors Required for Influenza Virus Replication. Nature 2010, 463, 813–817. [Google Scholar] [CrossRef] [PubMed]
- De Wilde, A.H.; Wannee, K.F.; Scholte, F.E.; Goeman, J.J.; ten Dijke, P.; Snijder, E.J.; Kikkert, M.; van Hemert, M.J. A Kinome-Wide Small Interfering RNA Screen Identifies Proviral and Antiviral Host Factors in Severe Acute Respiratory Syndrome Coronavirus Replication, Including Double-Stranded RNA-Activated Protein Kinase and Early Secretory Pathway Proteins. J. Virol. 2015, 89, 8318–8333. [Google Scholar] [CrossRef] [PubMed]
- Harmon, M.A.; Boehm, M.F.; Heyman, R.A.; Mangelsdorf, D.J. Activation of Mammalian Retinoid X Receptors by the Insect Growth Regulator Methoprene. Proc. Natl. Acad. Sci. USA 1995, 92, 6157–6160. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Jogi, M.; Onomoto, K. Regulation of Antiviral Innate Immune Signaling by Stress-Induced RNA Granules. J. Biochem. 2016, 159, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Vale, R.D. Myosin V Motor Proteins: Marching Stepwise towards a Mechanism. J. Cell Biol. 2003, 163, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Malikov, V.; Da Silva, E.S.; Jovasevic, V.; Bennett, G.; Vieira, D.A.D.S.A.; Schulte, B.; Diaz-Griffero, F.; Walsh, D.; Naghavi, M.H. HIV-1 Capsids Bind and Exploit the Kinesin-1 Adaptor Fez1 for Inward Movement to the Nucleus. Nat. Commun. 2015, 6, 6660. [Google Scholar] [CrossRef] [PubMed]
- Rajasekharan, S.; Kumar, K.; Rana, J.; Gupta, A.; Chaudhary, V.K.; Gupta, S. Host Interactions of Chandipura Virus Matrix Protein. Acta Trop. 2015, 149, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Blissard, G.W.; Theilmann, D.A. Trichoplusia ni Kinesin-1 Associates with Autographa Californica Multiple Nucleopolyhedrovirus Nucleocapsid Proteins and Is Required for Production of Budded Virus. J. Virol. 2016, 90, 3480–3495. [Google Scholar] [CrossRef] [PubMed]
- Diefenbach, R.J.; Davis, A.; Miranda-Saksena, M.; Fernandez, M.A.; Kelly, B.J.; Jones, C.A.; LaVail, J.H.; Xue, J.; Lai, J.; Cunningham, A.L. The Basic Domain of Herpes Simplex Virus 1 pUS9 Recruits Kinesin-1 to Facilitate Egress from Neurons. J. Virol. 2015, 90, 2102–2111. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Shibata, T.; Akashi-Takamura, S.; Kiyokawa, T.; Wakabayashi, Y.; Tanimura, N.; Kobayashi, T.; Matsumoto, F.; Fukui, R.; Kouro, T.; et al. A Protein Associated with Toll-Like Receptor (TLR) 4 (PRAT4A) Is Required for TLR-Dependent Immune Responses. J. Exp. Med. 2007, 204, 2963–2976. [Google Scholar] [CrossRef] [PubMed]
- Wiebe, M.S.; Jamin, A. The Barrier to Autointegration Factor: Interlocking Antiviral Defense with Genome Maintenance. J. Virol. 2016, 90, 3806–3809. [Google Scholar] [CrossRef] [PubMed]
- Kotani, E.; Yamakawa, M.; Iwamoto, S.I.; Tashiro, M.; Mori, H.; Sumida, M.; Matsubara, F.; Taniai, K.; Kadono-Okuda, K.; Kato, Y.; et al. Cloning and Expression of the Gene of Hemocytin, an Insect Humoral Lectin Which Is Homologous with the Mammalian Von Willebrand Factor. Biochim. Biophys. Acta 1995, 1260, 245–258. [Google Scholar] [CrossRef]
- Yamakawa, M.; Tanaka, H. Immune Proteins and Their Gene Expression in the Silkworm, Bombyx Mori. Dev. Comp. Immunol. 1999, 23, 281–289. [Google Scholar] [CrossRef]
- Gonzalez-Dosal, R.; Horan, K.A.; Rahbek, S.H.; Ichijo, H.; Chen, Z.J.; Mieyal, J.J.; Hartmann, R.; Paludan, S.R. Hsv Infection Induces Production of Ros, Which Potentiate Signaling from Pattern Recognition Receptors: Role for S-Glutathionylation of TRAF3 and 6. PLoS Pathog. 2011, 7, e1002250. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, A.; Amaya, M.; Voss, K.; Chung, M.; Benedict, A.; Sampey, G.; Kehn-Hall, K.; Luchini, A.; Liotta, L.; Bailey, C.; et al. Reactive Oxygen Species Activate NF-κB (P65) and P53 and Induce Apoptosis in RVFV Infected Liver Cells. Virology 2014, 449, 270–286. [Google Scholar] [CrossRef] [PubMed]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The NRF2 Cell Defence Pathway: KEAP1-Dependent and -Independent Mechanisms of Regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, R.; Li, M.; Frohlich, J. A Review on Lecithin: Cholesterol Acyltransferase Deficiency. Clin. Biochem. 2015, 48, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huang, P.; Yang, N.; Liu, R.; Liu, X.; Dai, H.; Zhang, L.; Song, F.; Sun, C. Establishment and Application of a High Throughput Screening System Targeting the Interaction between HCV Internal Ribosome Entry Site and Human Eukaryotic Translation Initiation Factor 3. Front. Microbiol. 2017, 8, 977. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.; Wang, W.; Xu, M.; Tu, J. Exploring the Roles of Basal Transcription Factor 3 in Eukaryotic Growth and Development. Biotechnol. Genet. Eng. Rev. 2015, 31, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Kochetov, G.A.; Solovjeva, O.N. Structure and Functioning Mechanism of Transketolase. Biochim. Biophys. Acta 2014, 1844, 1608–1618. [Google Scholar] [CrossRef] [PubMed]
- Farinha, C.M.; Matos, P.; Amaral, M.D. Control of Cystic Fibrosis Transmembrane Conductance Regulator Membrane Trafficking: Not Just from the Endoplasmic Reticulum to the Golgi. FEBS J. 2013, 280, 4396–4406. [Google Scholar] [CrossRef] [PubMed]
- Thio, C.L.P.; Yusof, R.; Ashrafzadeh, A.; Bahari, S.; Abdul-Rahman, P.S.; Karsani, S.A. Differential Analysis of the Secretome of WRL68 Cells Infected with the Chikungunya Virus. PLoS ONE 2015, 10, e0129033. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Sequences |
|---|---|---|
| Hemocytin | HMC-F | 5′AAGGATGCCTGCAATAGGTG3′ |
| HMC-R | 5′CCTCCGGAACACAGACAAAT3′ | |
| Lecithin-cholesterol acyltransferase | LCAT-F | 5′AGCCGTGAAGGTCTTTGCCATT3′ |
| LCAT-R | 5′TGCCAGGGATGGGGATGTAATTTG3′ | |
| Scramblase | SCR-F | 5′CCATGCGCCCATTTGACATGAA3′ |
| SCR-R | 5′GATTGACAGCGCAATGGACGATAG3′ | |
| Basic transcription factor 3 | BTF3-F | 5′AGTGCATAAGCAGGCAACACCA3′ |
| BTF3-R | 5′TACGTCGCCACCGAAATGAGTT3′ | |
| Juvenile hormone-inducible protein | JHIP-F | 5′CCTTGCTGAGCTCCTTGAGAAACT3′ |
| JHIP-R | 5′TACACATGGCCATTCCCATCTTCC3′ | |
| Eukaryotic translation initiation factor 3 | eTIF3-F | 5′TCGATAGGCATCTCACGTTTCCAC3′ |
| eTIF3-R | 5′TCTTCCCCACTGTATCCAGGATGT3′ | |
| Transmembrane protease serine 9-like | TPser9-F | 5′TCCACAATCCGGATGCAGACATAG3′ |
| TPser9-R | 5′CCAACCCGATTGACGATCTCAGAA3′ | |
| Transketolase | Transk-F | 5′TGTTAGCTGCGAACGTGCTGTA3′ |
| Transk-R | 5′GTATTTGGTCGGGATGTGCGAATG3′ | |
| Repressor splicing factor | RRM-F | 5′AATTTGGGAAGCTCAATA3′ |
| RRM-R | 5′GAGGATGTCGCAAGCCTTCT3′ | |
| forskolin-binding protein | FKBP-F | 5′TGAGTTTGAACGTGCCCAGGAT3′ |
| FKBP-R | 5′CTCCTTGATGTACTTGGCACCCTT3′ | |
| Coatomer delta subunit | ClatAdp-F | 5′TAGCCGATGAGAAGTTCGGGAAGA3′ |
| ClatAdp-R | 5′CTTATCGACATTGGGGTGCGTTTG3′ | |
| Proliferating cell nuclear antigen | PCNA-F | 5′CATGAATCTCGACCAGGAGCACTT3′ |
| PCNA-R | 5′TCACGGCAAATGCGTGCAAA3′ | |
| Tubulin-specific chaperone A | TBCA-F | 5′CGTACGAAAAGGAAGCAGATCAGCA3′ |
| TBCA-R | 5′CTTCCTTCCGGATCACGTGTTCAT3′ | |
| Ribosomal protein 49 | rp49-F | 5′GACCGATATGCCAAGCTAAAGCA3′ |
| rp49-R | 5′GGGGAGCATGTGGCGTGTCTT3′ |
| Samples | 6 h mRNA | 12 h mRNA | 24 h mRNA | 48 h mRNA | ||||
|---|---|---|---|---|---|---|---|---|
| r | r2 | r | r2 | r | r2 | r | r2 | |
| 24 h secreted protein | 0.1742 | 0.0304 | −0.1070 | 0.0115 | −0.2399 | 0.0576 | −0.2265 | 0.0513 |
| 48 h secreted protein | 0.2024 | 0.0410 | 0.3754 | 0.1409 | −0.1197 | 0.0143 | −0.4018 | 0.1615 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins-da-Silva, A.; Telleria, E.L.; Batista, M.; Marchini, F.K.; Traub-Csekö, Y.M.; Tempone, A.J. Identification of Secreted Proteins Involved in Nonspecific dsRNA-Mediated Lutzomyia longipalpis LL5 Cell Antiviral Response. Viruses 2018, 10, 43. https://doi.org/10.3390/v10010043
Martins-da-Silva A, Telleria EL, Batista M, Marchini FK, Traub-Csekö YM, Tempone AJ. Identification of Secreted Proteins Involved in Nonspecific dsRNA-Mediated Lutzomyia longipalpis LL5 Cell Antiviral Response. Viruses. 2018; 10(1):43. https://doi.org/10.3390/v10010043
Chicago/Turabian StyleMartins-da-Silva, Andrea, Erich Loza Telleria, Michel Batista, Fabricio Klerynton Marchini, Yara Maria Traub-Csekö, and Antonio Jorge Tempone. 2018. "Identification of Secreted Proteins Involved in Nonspecific dsRNA-Mediated Lutzomyia longipalpis LL5 Cell Antiviral Response" Viruses 10, no. 1: 43. https://doi.org/10.3390/v10010043
APA StyleMartins-da-Silva, A., Telleria, E. L., Batista, M., Marchini, F. K., Traub-Csekö, Y. M., & Tempone, A. J. (2018). Identification of Secreted Proteins Involved in Nonspecific dsRNA-Mediated Lutzomyia longipalpis LL5 Cell Antiviral Response. Viruses, 10(1), 43. https://doi.org/10.3390/v10010043