All Known Human Rhinovirus Species Are Present in Sputum Specimens of Military Recruits During Respiratory Infection

Abstract
:1. Introduction
2. Results and Discussion
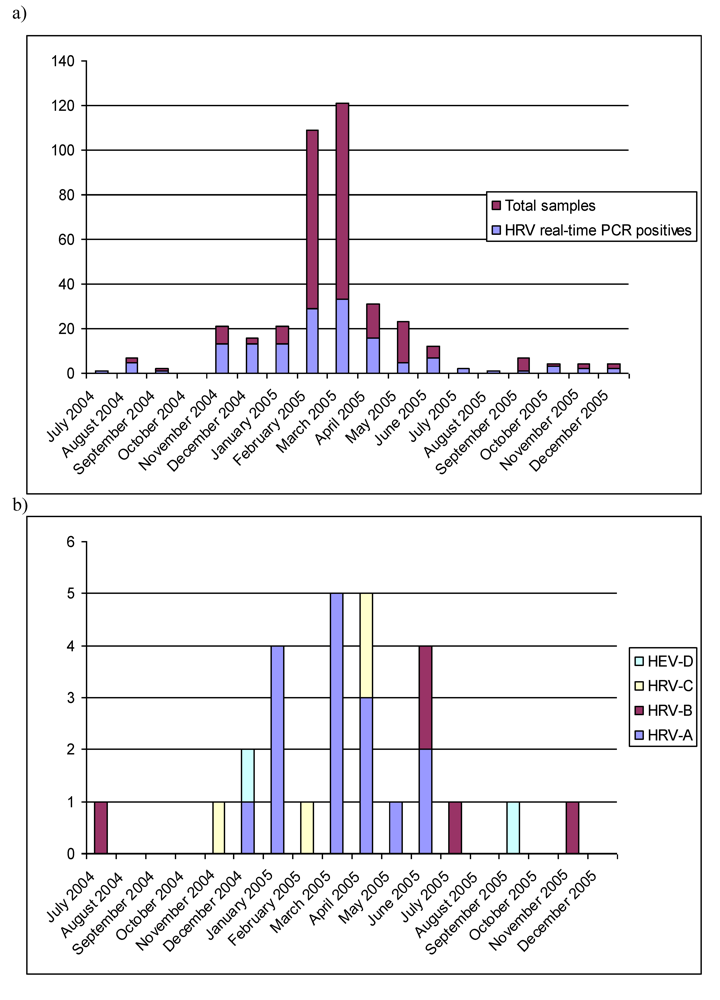
2.1. Real-time RT-PCR
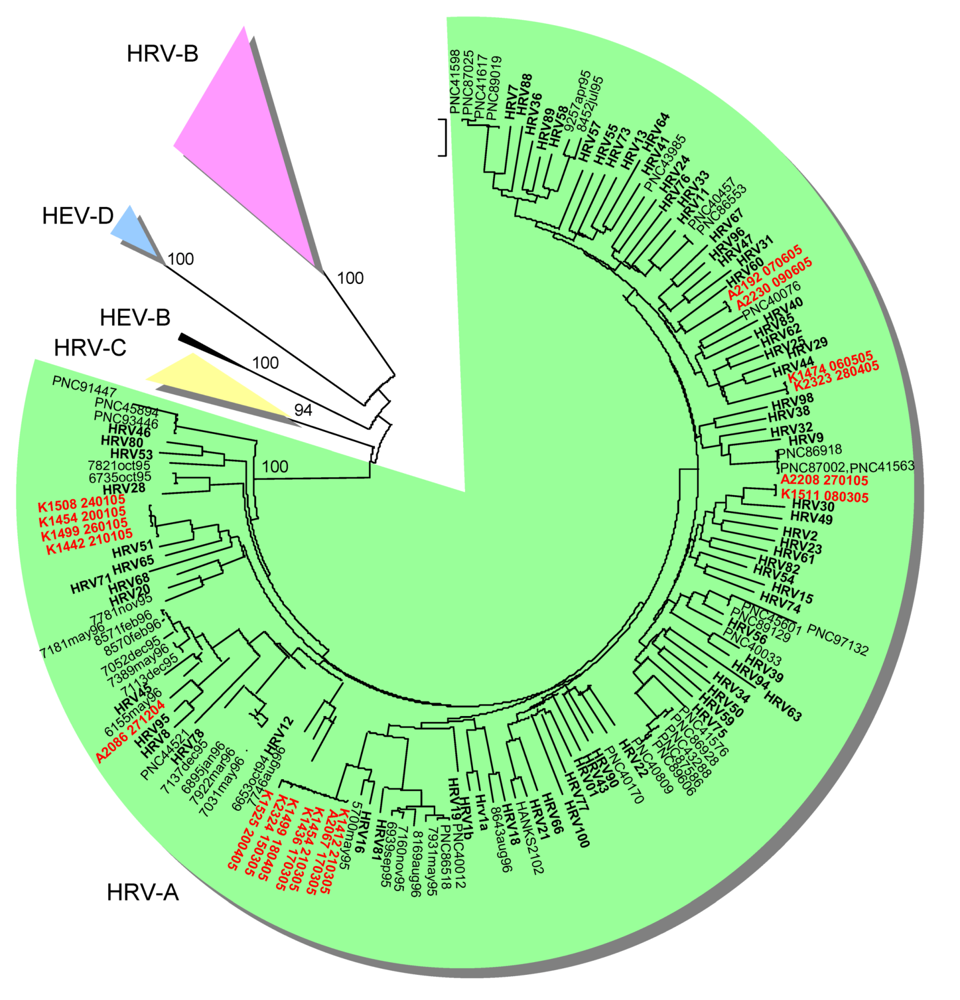
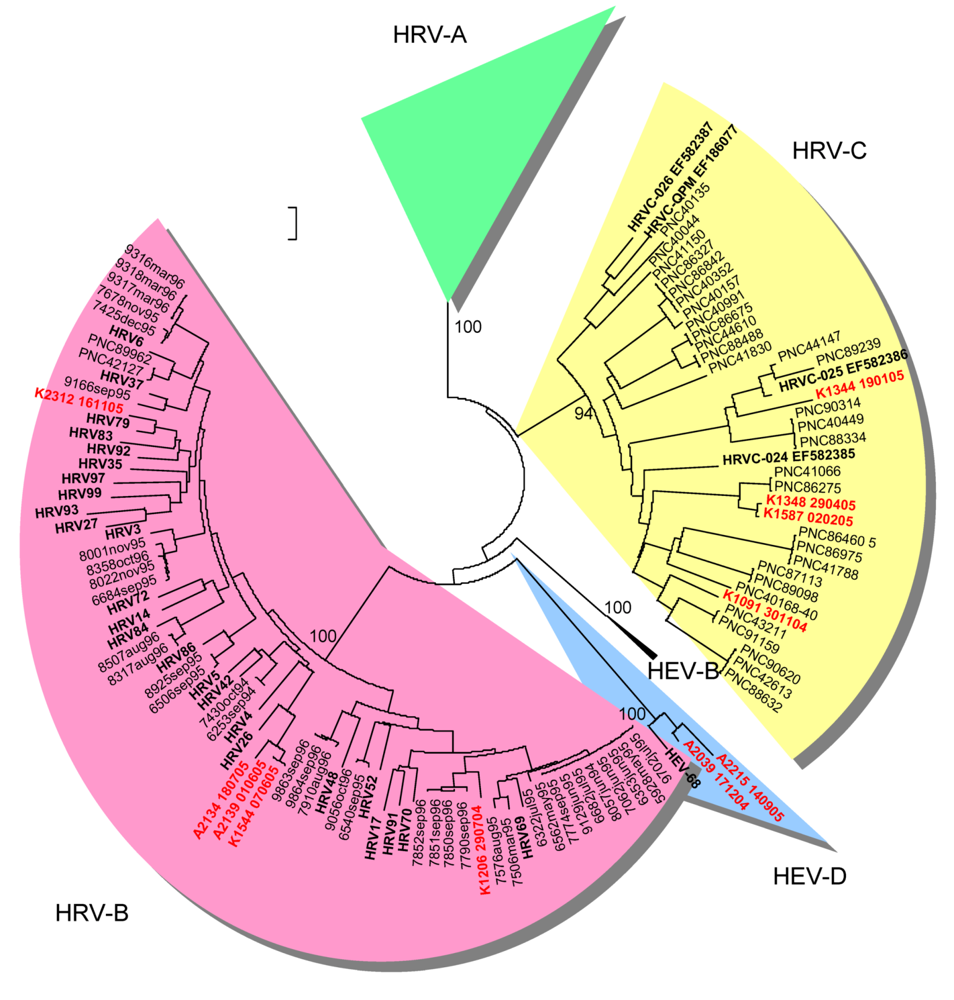
2.2. Genetic typing in the capsid coding region

{kind=link}
{kind=link}
{kind=link}
| Typing in the VP4/VP2 coding region | Typing in the partial VP1 coding region | ||||||
|---|---|---|---|---|---|---|---|
| Collection date | Strain ID | HRV/HEV Real time RT-PCR result | The closest prototype strain or GenBank match* | Genetic similarity | The closest prototype strain or GenBank match | Genetic similarity | |
| 2004-7-29 | 1206 | HRV+/HEV- | HRV91 (HRV-B) | 90.0 % | |||
| 2004-11-30 | 1091 | HRV+/ HEV+ | NAT045 (HRV-C) | 82.5% | untypable^ | ||
| 2004-12-9 | 2119 | HRV+/ HEV+ | untypable^ | HEV68 (HEV-D) | 86.3% | ||
| 2004-12-17 | 2039 | HRV+/ HEV+ | HRV87 (HEV-D) | 91.8% | untypable^ | ||
| 2004-12-17 | 1105 | HRV+/ HEV+ | untypable^ | HEV68 (HEV-D) | 86.3% | ||
| 2004-12-27 | 2086 | HRV+/HEV- | HRV95 (HRV-A) | 92.5% | |||
| 2005-1-20 | 1454 | HRV+/HEV- | HRV51 (HRV-A) | 92.5% | |||
| 2005-1-21 | 1442 | HRV+/ HEV+ | HRV51 (HRV-A) | 92.3% | HRV51 (HRV-A) | 88.4% | |
| 2005-1-24 | 1508 | HRV+/ HEV+ | HRV51 (HRV-A) | 93.4% | HRV51 (HRV-A) | 88.4% | |
| 2005-1-26 | 1499 | HRV+/ HEV+ | HRV51 (HRV-A) | 93.1% | HRV51 (HRV-A) | 88.4% | |
| 2005-1-27 | 2208 | HRV+/HEV- | HRV30 (HRV-A) | 93.8% | |||
| 2005-2-2 | 1587 | HRV+/ HEV+ | 06-339 (HRV-C) | 99.0 % | |||
| 2005-2-22 | 2145 | HRV-/ HEV+ | untypable^ | HEV68 (HEV-D) | 86.0% | ||
| 2005-3-8 | 1511 | HRV+/HEV- | HRV30 (HRV-A) | 94.0 % | |||
| 2005-3-15 | 2324 | HRV+/HEV- | HRV16 (HRV-A) | 92.9% | |||
| 2005-3-17 | 2067 | HRV+/HEV- | HRV16 (HRV-A) | 92.7% | |||
| 2005-3-17 | 1436 | HRV+/HEV- | HRV16 (HRV-A) | 92.9% | |||
| 2005-3-21 | 1412 | HRV+/HEV- | HRV16 (HRV-A) | 92.6% | |||
| 2005-3-21 | 1454 | HRV+/ HEV+ | HRV16 (HRV-A) | 92.7% | |||
| 2005-4-18 | 1344 | HRV+/HEV- | N38 (HRV-C) | 87.3% | |||
| 2005-4-18 | 1499 | HRV+/HEV- | HRV16 (HRV-A) | 94.6% | |||
| 2005-4-20 | 1525 | HRV+/HEV- | HRV16 (HRV-A) | 91.5% | |||
| 2005-4-28 | 2323 | HRV+/HEV- | HRV29 (HRV-A) | 91.6% | |||
| 2005-4-29 | 1348 | HRV+/ HEV+ | N27 (HRV-C) | 98.2% | |||
| 2005-5-6 | 1474 | HRV+/HEV- | HRV44 (HRV-A) | 91.3% | |||
| 2005-6-1 | 2139 | HRV+/HEV- | HRV26 (HRV-B) | 92.8% | |||
| 2005-6-7 | 2192 | HRV+/ HEV+ | HRV55 (HRV-A) | 85.0 % | |||
| 2005-6-7 | 1544 | HRV+/HEV- | HRV26 (HRV-B) | 92.9% | |||
| 2005-6-9 | 2230 | HRV+/HEV- | HRV57 (HRV-A) | 89.0 % | |||
| 2005-7-18 | 2134 | HRV+/HEV- | HRV26 (HRV-B) | 89.5% | |||
| 2005-9-14 | 2215 | HRV+/ HEV+ | HEV 68 (HEV-D) | 87.6% | untypable^ | ||
| 2005-11-16 | 2312 | HRV+/HEV- | HRV79 (HRV-B) | 92.0 % | |||
2.3. Epidemiological characteristics
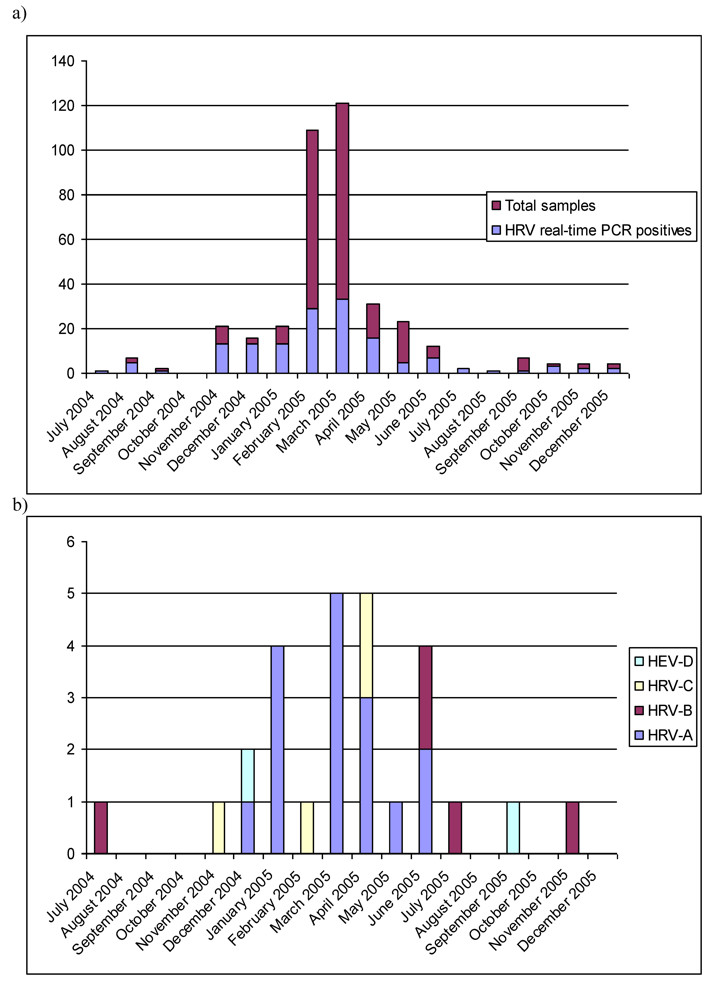
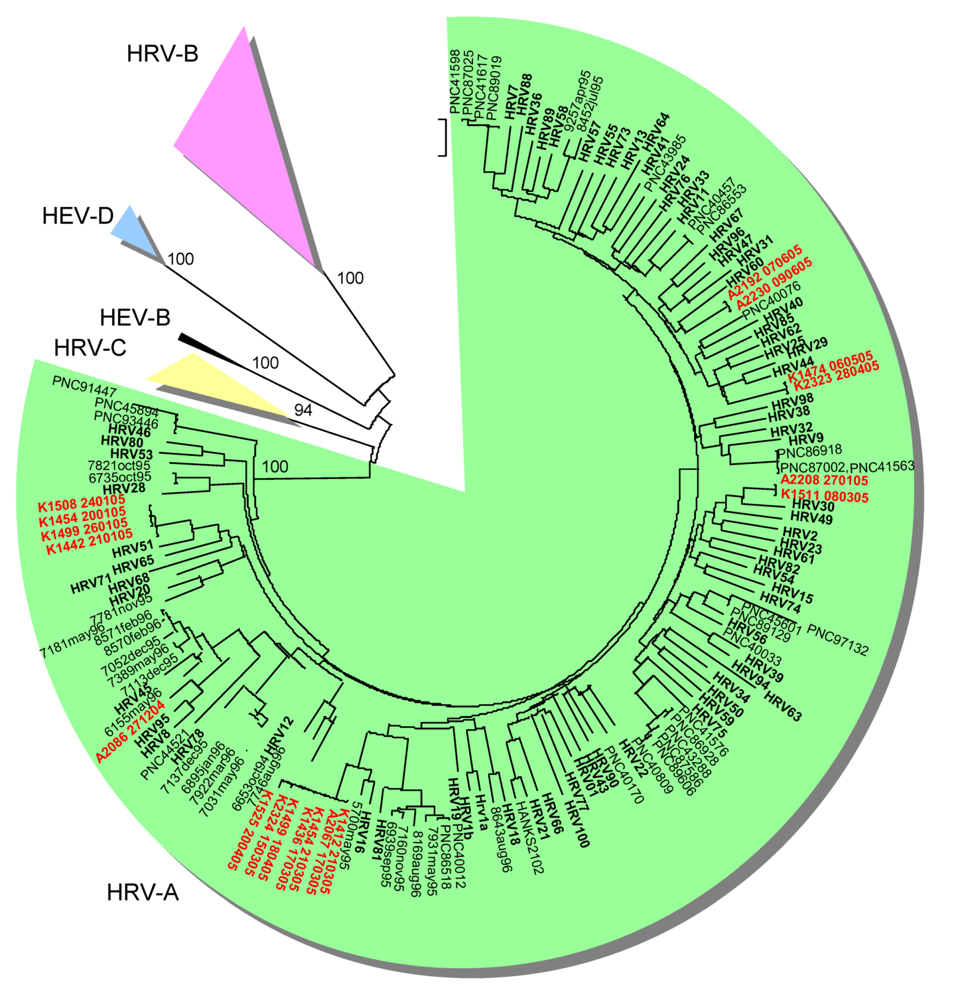
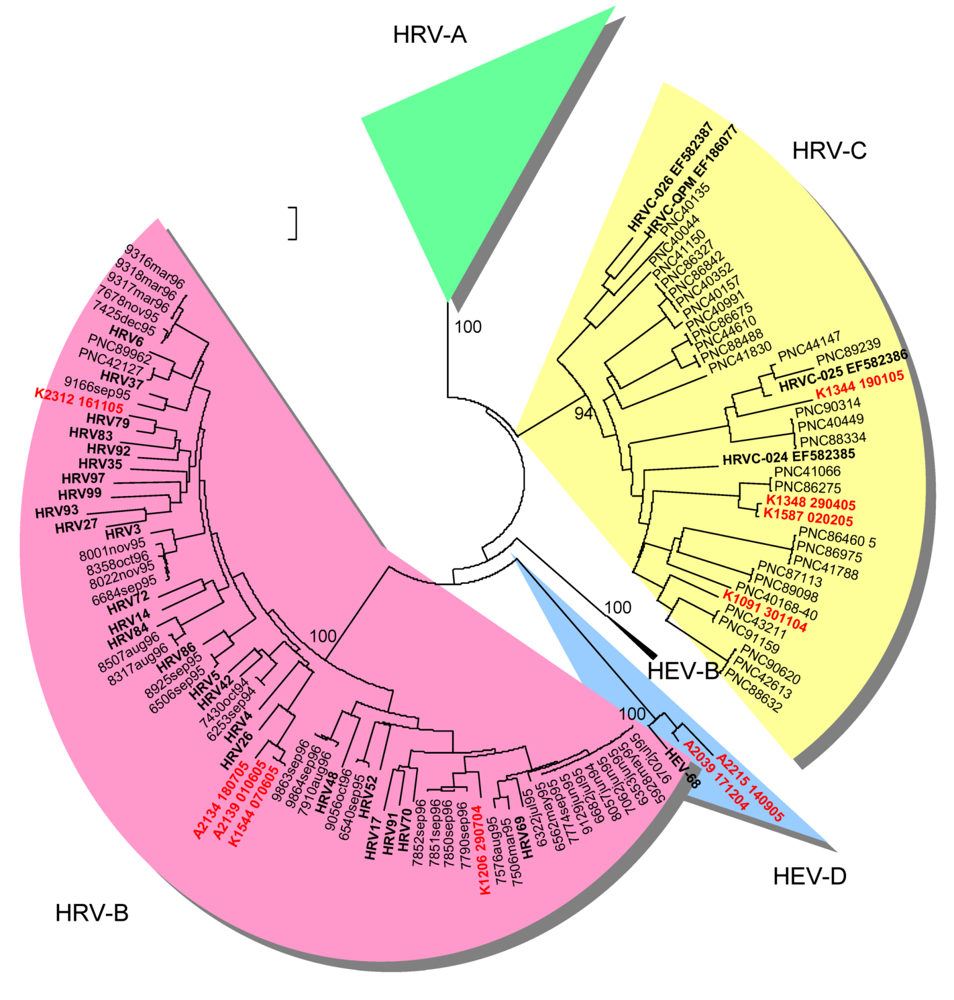
2.4. Limitations of the study and of the methods in the diagnostics of HRV and HEV
3. Experimental Section
3.1. Samples
3.2. Real-time RT-PCR
3.3. RT-PCR and sequencing
4. Conclusions
Acknowledgments
References
- Gern, J.E.; Galagan, D.M.; Jarjour, N.N.; Dick, E.C.; Busse, W.W. Detection of rhinovirus RNA in lower airway cells during experimentally induced infection. Am. J. Respir. Crit. Care Med. 1997, 155, 1159–1161. [Google Scholar] [PubMed]
- Papadopoulos, N.G.; Sanderson, G.; Hunter, J.; Johnston, S.L. Rhinoviruses replicate effectively at lower airway temperatures. J. Med. Virol. 1999, 58, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Schroth, M.K.; Grimm, E.; Frindt, P.; Galagan, D.M.; Konno, S.I.; Love, R.; Gern, J.E. Rhinovirus replication causes RANTES production in primary bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 1999, 20, 1220–1228. [Google Scholar] [PubMed]
- Papadopoulos, N.G.; Bates, P.J.; Bardin, P.G.; Papi, A.; Leir, S.H.; Fraenkel, D.J.; Meyer, J.; Lackie, P.M.; Sanderson, G.; Holgate, S.T.; Johnston, S.L. Rhinoviruses infect the lower airways. J. Infect. Dis. 2000, 181, 1875–1884. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, N.G.; Xepapadaki, P.; Mallia, P.; Brusselle, G.; Watelet, J.B.; Xatzipsalti, M.; Foteinos, G.; van Drunen, C.M.; Fokkens, W.J.; D'Ambrosio, C.; Bonini, S.; Bossios, A.; Lotvall, J.; van Cauwenberge, P.; Holgate, S.T.; Canonica, G.W.; Szczeklik, A.; Rohde, G.; Kimpen, J.; Pitkaranta, A.; Makela, M.; Chanez, P.; Ring, J.; Johnston, S.L. Mechanisms of virus-induced asthma exacerbations: state-of-the-art A GA2LEN and InterAirways document. Allergy 2007, 62, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Hayden, F.G. Rhinovirus and the lower respiratory tract. Rev. Med. Virol. 2004, 14, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Stanway, G.; Brown, F.; Christian, P.; Hovi, T.; Hyypiä, T.; King, A.M.Q.; Knowles, N.J.; Lemon, S.M.; Minor, P.D.; Pallansch, M.A.; Palmenberg, A.C.; Skern, T. Family Picornaviridae. Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; In Virus Taxonomy; Elsevier/Academic Press: London, UK, 2005. [Google Scholar]
- Knowles, N.J. New species in the genus Enterovirus (Picornaviridae)2008. Available online: http://talk.ictvonline.org/media/g/vert01/default.aspx (accessed 3 December 2009).
- Han, T.H.; Chung, J.Y.; Hwang, E.S.; Koo, J.W. Detection of human rhinovirus C in children with acute lower respiratory tract infections in South Korea. Arch. Virol. 2009, 154, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Linsuwanon, P.; Payungporn, S.; Samransamruajkit, R.; Posuwan, N.; Makkoch, J.; Theanboonlers, A.; Poovorawan, Y. High prevalence of human rhinovirus C infection in Thai children with acute lower respiratory tract disease. J. Infect. 2009, 59, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Piralla, A.; Rovida, F.; Campanini, G.; Rognoni, V.; Marchi, A.; Locatelli, F.; Gerna, G. Clinical severity and molecular typing of human rhinovirus C strains during a fall outbreak affecting hospitalized patients. J. Clin. Virol. 2009, 45, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Tapparel, C.; L'Huillier, A.G.; Rougemont, A.L.; Beghetti, M.; Barazzone-Argiroffo, C.; Kaiser, L. Pneumonia and pericarditis in a child with HRV-C infection: a case report. J. Clin. Virol. 2009, 45, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Louie, J.K.; Roy-Burman, A.; Guardia-Labar, L.; Boston, E.J.; Kiang, D.; Padilla, T.; Yagi, S.; Messenger, S.; Petru, A.M.; Glaser, C.A.; Schnurr, D.P. Rhinovirus associated with severe lower respiratory tract infections in children. Pediatr. Infect. Dis. J. 2009, 28, 337–339. [Google Scholar] [CrossRef] [PubMed]
- International Committee on Taxonomy of Viruses. 2008. Virus Taxonomy 2008. International Committee on Taxonomy of Viruses. Available online: http://www.ictvonline.org/virusTaxonomy.asp? (accessed 3 December 2009).
- Kistler, A.; Avila, P.C.; Rouskin, S.; Wang, D.; Ward, T.; Yagi, S.; Schnurr, D.; Ganem, D.; DeRisi, J.L.; Boushey, H.A. Pan-viral screening of respiratory tract infections in adults with and without asthma reveals unexpected human coronavirus and human rhinovirus diversity. J. Infect. Dis. 2007, 196, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Louie, J.K.; Hacker, J.K.; Gonzales, R.; Mark, J.; Maselli, J.H.; Yagi, S.; Drew, W.L. Characterization of viral agents causing acute respiratory infection in a San Francisco University Medical Center Clinic during the influenza season. Clin. Infect. Dis. 2005, 41, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Mosser, A.G.; Vrtis, R.; Burchell, L.; Lee, W.M.; Dick, C.R.; Weisshaar, E.; Bock, D.; Swenson, C.A.; Cornwell, R.D.; Meyer, K.C.; Jarjour, N.N.; Busse, W.W.; Gern, J.E. Quantitative and qualitative analysis of rhinovirus infection in bronchial tissues. Am. J. Respir. Crit. Care Med. 2005, 171, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Savolainen-Kopra, C.; Blomqvist, S.; Kilpi, T.; Roivainen, M.; Hovi, T. Novel species of human rhinoviruses in acute otitis media. Pediatr. Infect. Dis. J. 2009, 28, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Nix, W.A.; Oberste, M.S.; Pallansch, M.A. Sensitive, Seminested PCR Amplification of VP1 Sequences for Direct Identification of All Enterovirus Serotypes from Original Clinical Specimens. J. Clin. Microbiol. 2006, 44, 2698–2704. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, C.; Mulders, M.N.; Hovi, T. Phylogenetic analysis of rhinovirus isolates collected during successive epidemic seasons. Virus Res. 2002, 85, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Renwick, N. A recently identified rhinovirus genotype is associated with severe respiratory-tract infection in children in Germany. J. Infect. Dis. 2007, 196, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Briese, T.; Renwick, N.; Venter, M.; Jarman, R.G.; Ghosh, D.; Kondgen, S.; Shrestha, S.K.; Hoegh, A.M.; Casas, I.; Adjogoua, E.V.; Akoua-Koffi, C.; Myint, K.S.; Williams, D.T.; Chidlow, G.; van den Berg, R.; Calvo, C.; Koch, O.; Palacios, G.; Kapoor, V.; Villari, J.; Dominguez, S.R.; Holmes, K.V.; Harnett, G.; Smith, D.; Mackenzie, J.S.; Ellerbrok, H.; Schweiger, B.; Schonning, K.; Chadha, M.S.; Leendertz, F.H.; Mishra, A.C.; Gibbons, R.V.; Holmes, E.C.; Lipkin, W.I. Global distribution of novel rhinovirus genotype. Emerg. Infect. Dis. 2008, 14, 944–947. [Google Scholar] [CrossRef] [PubMed]
- McErlean, P.; Shackelton, L.A.; Andrews, E.; Webster, D.R.; Lambert, S.B.; Nissen, M.D.; Sloots, T.P.; Mackay, I.M. Distinguishing molecular features and clinical characteristics of a putative new rhinovirus species, human rhinovirus C (HRV C). PLoS ONE 2008, 3, e1847. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Yip, C.C.; Tsoi, H.W.; Lee, R.A.; So, L.Y.; Lau, Y.L.; Chan, K.H.; Woo, P.C.; Yuen, K.Y. Clinical features and complete genome characterization of a distinct human rhinovirus (HRV) genetic cluster, probably representing a previously undetected HRV species, HRV-C, associated with acute respiratory illness in children. J. Clin. Microbiol. 2007, 45, 3655–3664. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, S.; Savolainen, C.; Raman, L.; Roivainen, M.; Hovi, T. Human rhinovirus 87 and enterovirus 68 represent a unique serotype with rhinovirus and enterovirus features. J. Clin. Microbiol. 2002, 40, 4218–4223. [Google Scholar] [CrossRef] [PubMed]
- Ishiko, H.; Miura, R.; Shimada, Y.; Hayashi, A.; Nakajima, H.; Yamazaki, S.; Takeda, N. Human rhinovirus 87 identified as human enterovirus 68 by VP4-based molecular diagnosis. Intervirology 2002, 45, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Juvonen, R.; Bloigu, A.; Peitso, A.; Silvennoinen-Kassinen, S.; Saikku, P.; Leinonen, M.; Harju, T. Risk factors for acute respiratory tract illness in military conscripts. Respirology 2008, 13, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Holloway, B.; Dare, R.K.; Kuypers, J.; Yagi, S.; Williams, J.V.; Hall, C.B.; Erdman, D.D. Real-time reverse transcription-PCR assay for comprehensive detection of human rhinoviruses. J. Clin. Microbiol. 2008, 46, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Savolainen-Kopra, C.; Blomqvist, S.; Kaijalainen, S.; Jounio, U.; Juvonen, R.; Peitso, A.; Saukkoriipi, A.; Vainio, O.; Hovi, T.; Roivainen, M. All Known Human Rhinovirus Species Are Present in Sputum Specimens of Military Recruits During Respiratory Infection. Viruses 2009, 1, 1178-1189. https://doi.org/10.3390/v1031178
Savolainen-Kopra C, Blomqvist S, Kaijalainen S, Jounio U, Juvonen R, Peitso A, Saukkoriipi A, Vainio O, Hovi T, Roivainen M. All Known Human Rhinovirus Species Are Present in Sputum Specimens of Military Recruits During Respiratory Infection. Viruses. 2009; 1(3):1178-1189. https://doi.org/10.3390/v1031178
Chicago/Turabian StyleSavolainen-Kopra, Carita, Soile Blomqvist, Svetlana Kaijalainen, Ulla Jounio, Raija Juvonen, Ari Peitso, Annika Saukkoriipi, Olli Vainio, Tapani Hovi, and Merja Roivainen. 2009. "All Known Human Rhinovirus Species Are Present in Sputum Specimens of Military Recruits During Respiratory Infection" Viruses 1, no. 3: 1178-1189. https://doi.org/10.3390/v1031178
APA StyleSavolainen-Kopra, C., Blomqvist, S., Kaijalainen, S., Jounio, U., Juvonen, R., Peitso, A., Saukkoriipi, A., Vainio, O., Hovi, T., & Roivainen, M. (2009). All Known Human Rhinovirus Species Are Present in Sputum Specimens of Military Recruits During Respiratory Infection. Viruses, 1(3), 1178-1189. https://doi.org/10.3390/v1031178