First Report on Establishment of Laricobius osakensis (Coleoptera: Derodontidae), a Biological Control Agent for Hemlock Woolly Adelgid, Adelges tsugae (Hemiptera: Adelgidae), in the Eastern U.S.

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sites
2.2. Evaluating Tree Health and HWA Density
2.3. Laricobius Sampling
2.3.1. Beat Sheet Sampling
2.3.2. Branch Clip Sampling
2.4. Laricobius Dispersal
2.5. Statistical Analysis
3. Results
3.1. Evaluating Tree Health Index and HWA Density
3.2. Recovery of L. osakensis
3.3. Dispersal Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Havill, N.P.; Montgomery, M.E.; Yu, G.; Shiyake, S.; Caccone, A. Mitochondrial DNA from hemlock woolly adelgid (Hemiptera: Adelgidae) suggests cryptic speciation and pinpoints the source of the introduction to eastern North America. Ann. Entomol. Soc. Am. 2006, 99, 195–203. [Google Scholar] [CrossRef]
- Orwig, D.A.; Foster, D.R.; Mausel, D.L. Landscape patterns of hemlock decline in New England due to the introduced hemlock woolly adelgid. J. Biogeogr. 2002, 29, 1475–1487. [Google Scholar] [CrossRef]
- Daley, M.J.; Philips, N.G.; Pettijohn, C.; Hadley, J.L. Water use by eastern hemlock (Tsuga canadensis) and black birch (Betula lenta): Implications of effects of the hemlock woolly adelgid. Can. J. For. Res. 2007, 37, 2031–2040. [Google Scholar] [CrossRef]
- Lamb, A.; Montgomery, M.E.; Vieira, L.C.; Shiyake, S.; Salom, S.M. Laricobius osakensis, a hemlock woolly adelgid predator from Japan. In Implementation and Status Biological Control of the Hemlock Woolly Adelgid. FHTET-2011-04; Onken, B., Reardon, R., Eds.; Department of Agriculture, Forest Service Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2011; pp. 90–96. [Google Scholar]
- Clark, R.C.; Brown, N.R. Studies of predators of the balsam woolly aphid, Adelges piceae (Ratz.) (Homoptera: Adelgidae), VII. Laricobius rubidus Lec. (Coleoptera: Derodontidae), a predator of Pineus strobi (Htg.) (Homoptera: Adelgidae). Can. Entomol. 1960, 92, 237–240. [Google Scholar] [CrossRef]
- Zilahi-Balogh, G.M.G.; Humble, L.M.; Lamb, A.B.; Salom, S.M.; Kok, L.T. Seasonal abundance and synchrony between Laricobius nigrinus (Coleoptera: Derodontidae) and its prey, the hemlock woolly adelgid (Hemiptera: Adelgidae). Can. Entomol. 2003, 135, 103–115. [Google Scholar] [CrossRef]
- Mausel, D.L.; Salom, S.M.; Kok, L.T.; Davis, G.A. Establishment of the hemlock woolly adelgid predator, Laricobius nigrinus (Coleoptera: Derodontidae), in the eastern United States. Environ. Entomol. 2010, 39, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Lamb, A.B.; Salom, S.M.; Kok, L.T.; Mausel, D.L. Confined field release of Laricobius nigrinus (Coleoptera: Derodontidae), a predator of the hemlock woolly adelgid, Adelges tsugae (Hemiptera: Adelgidae), in Virginia. Can. J. For. Res. 2006, 36, 369–375. [Google Scholar] [CrossRef]
- Mayfield, A.E.; Reynolds, B.C.; Coots, C.I.; Havill, N.P.; Brownie, C.; Tait, A.R.; Hanula, J.L.; Joseph, S.V.; Galloway, A.B. Establishment, hybridization and impact of Laricobius predators on insecticide-treated hemlocks: Exploring integrated management of the hemlock woolly adelgid. For. Ecol. Manag. 2015, 335, 1–10. [Google Scholar] [CrossRef]
- Montgomery, M.E.; Shiyake, S.; Havill, N.P.; Leschen, R.A.B. A new species of Laricobius (Coleoptera: Derodontidae from Japan with phylogeny and a key for native and introduced congeners in North America. Ann. Entomol. Soc. Am. 2011, 104, 389–401. [Google Scholar] [CrossRef]
- Vieira, L.C.; Lamb, A.B.; Shiyake, S.; Salom, S.M.; Kok, L.T. Seasonal abundance and synchrony between Laricobius osakensis Montgomery & Shiyake (Coleoptera: Derodontidae) and its prey, Adelges tsugae Annand (Hemiptera: Adelgidae) in Japan. Ann. Entomol. Soc. Am. 2013, 106, 249–257. [Google Scholar]
- Vieira, L.C.; Mcavoy, T.J.; Chantos, J.; Lamb, A.B.; Salom, S.M.; Kok, L.T. Host range of Laricobius osakensis (Coleoptera: Derodontidae), a new biological control agent of hemlock woolly adelgid (Hemiptera: Adelgidae). Environ. Entomol. 2011, 40, 324–332. [Google Scholar] [CrossRef]
- Story, H.M.; Vieira, L.C.; Salom, S.M.; Kok, L.T. Assessing performance and competition among three Laricobius (Coleoptera: Derodontidae) species, predators of hemlock woolly adelgid, Adelges tsugae (Hemiptera: Adelgidae). Environ Entomol. 2012, 41, 896–904. [Google Scholar] [CrossRef]
- Fischer, M.J.; Brewster, C.C.; Havill, N.P.; Salom, S.M.; Kok, L.T. Assessment of the potential for hybridization between Laricobius nigrinus (Coleoptera: Derodontidae) and Laricobius osakensis, predators of the hemlock woolly adelgid (Hemiptera: Adelgidae). Biocontrol Sci. Technol. 2015, 25, 1467–1482. [Google Scholar] [CrossRef]
- Roberts, A.; Lamb, A.; Onken, B.; Salom, S.M. HWA Predator Release and Monitoring Database. Last Modified: May 2018. Available online: http://hiro.ento.vt.edu/pdb/ (accessed on 6 October 2015).
- Mooneyham, K.; Loke, T.; Kok, K.; Salom, S.M. Release and colonization of Laricobius osakensis (Coleoptera: Derodontidae), a predator of the hemlock woolly adelgid, Adelges tsugae. Northeast. Nat. 2016, 23, 141–150. [Google Scholar] [CrossRef]
- USDA Plant Hardiness Zone Map. Agricultural Research Service, U.S. Department of Agriculture. Available online: http://planthardiness.ars.usda.gov (accessed on 3 April 2016).
- Sumpter, K.L.; McAvoy, T.J.; Brewster, C.C.; Mayfield, A.E.; Salom, S.M. Assessing an integrated biological and chemical control stratefy for managing hemlock woolly adelgid in southern Appalachian forests. For. Ecol. Manag. 2018, 411, 12–19. [Google Scholar] [CrossRef]
- Salom, S.M.; Kok, L.T.; Lamb, A.B.; Jubb, C. Laboratory rearing of Laricobius nigrinus (Coleoptera: Derodontidae): A predator of the hemlock woolly adelgid (Hemiptera: Adelgidae). Psyche 2012, 1–9. [Google Scholar]
- Lamb, A.B.; Salom, S.M.; Kok, L.T. Survival and reproduction of Laricobius nigrinus Fender (Coleoptera: Derodontidae), a predator of hemlock woolly adelgid, Adelges tsugae Annand (Homoptera: Adelgidae), in field cages. Biol. Contr. 2005, 32, 200–207. [Google Scholar] [CrossRef]
- Davis, G.A.; Havill, N.P.; Adelman, Z.N.; Caccone, A.; Kok, L.T.; Salom, S.M. DNA barcodes and molecular diagnostics to distinguish an introduced and native Laricobius (Coleoptera: Derodontidae) species in eastern North America. Biol. Contr. 2011, 58, 53–59. [Google Scholar] [CrossRef]
- Fischer, M.J.; Havill, N.P.; Jubb, C.S.; Prosser, S.W.; Opell, B.D.; Salom, S.M.; Kok, L.T. Contamination delays the release of Laricobius osakensis for biological control of hemlock woolly adelgid: Cryptic diversity in Japanese Laricobius spp. and colony purification techniques. Southeast. Nat. 2014, 13, 178–191. [Google Scholar]
- Rijal, J.P.; Brewster, C.C.; Bergh, J.C. Effects of biotic and abiotic factors on grape root borer (Lepidoptera: Sesiidae) infestations in commercial vineyards in Virginia. Environ. Entomol. 2014, 43, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Freund, R.J.; Wilson, W.J.; Sa, P. Statistical Modeling of a Response Variable. In Regression Analysis; Academic Press: St. Louis, MI, USA, 1998. [Google Scholar]
- Maroof, D.A. (Ed.) Binary Logistic Regression. In Statistical Methods in Neuropsychology: Common Procedures Made Comprehensible; Springer and Business Media: New York, NY, USA, 2012; pp. 66–67. [Google Scholar]
- Venette, R.C.; Moon, R.D.; Hutchinson, W.D. Strategies and statistics of sampling for rare individuals. Ann. Rev. Entomol. 2002, 47, 143–174. [Google Scholar] [CrossRef] [PubMed]
- McAvoy, T.J.; Régnière, J.; St-Amant, R.; Schneeberger, N.F.; Salom, S.M. Mortality and recovery of hemlock woolly adelgid (Adelges tsugae) in response to winter temperatures and predictions for the future. Forests 2017, 8, 497. [Google Scholar] [CrossRef]
- McClure, M.S. Density-dependent feedback and population cycles in Adelges tsugae (Homoptera: Adelgidae) on Tsuga canadensis. Environ. Entomol. 1991, 20, 258–264. [Google Scholar] [CrossRef]
- Allee, W.C. Animal Aggregations: A Study in General Sociology; University of Chicago Press: Chicago, IL, USA, 1931. [Google Scholar]
- Bajeux, N.; Grongnard, F.; Maileret, L. Introduction Strategies for biological control agents subject to Allee effects. In Proceedings of the 21st International Symposium on Mathematical Theory of Networks and Systems, Goningen, The Netherlands, 7–14 July 2014. [Google Scholar]
- Shea, K.; Possingham, H.P. Optiamal Release strategies for biological control agents: An application of stochastic dynamic programming to population management. J. Appl. Ecol. 2000, 37, 77–86. [Google Scholar] [CrossRef]
- Mausel, D.L.; Salom, S.M.; Kok, L.T.; Fidgen, J.G. Propagation, synchrony, and impact of introduced and native Laricobius spp. (Coleoptera: Derodontidae) on hemlock woolly adelgid in Virginia. Environ. Entomol. 2008, 37, 1498–1507. [Google Scholar] [CrossRef] [PubMed]
- Wantuch, H.A.; Kuhar, T.P.; Salom, S.M. Phenology of the pine bark adelgid, Pineus strobi (Hemiptera: Adelgidae), in white pine forests of southwestern Virginia. Environ. Entomol. 2017, 46, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.A.; Salom, S.M.; Brewster, C.C.; Onken, B.P.; Kok, L.T. Spatiotemporal distribution of the hemlock woolly adelgid predator Laricobius nigrinus after release in eastern forests. Agric. For. Entomol. 2012, 14, 2031–2040. [Google Scholar] [CrossRef]
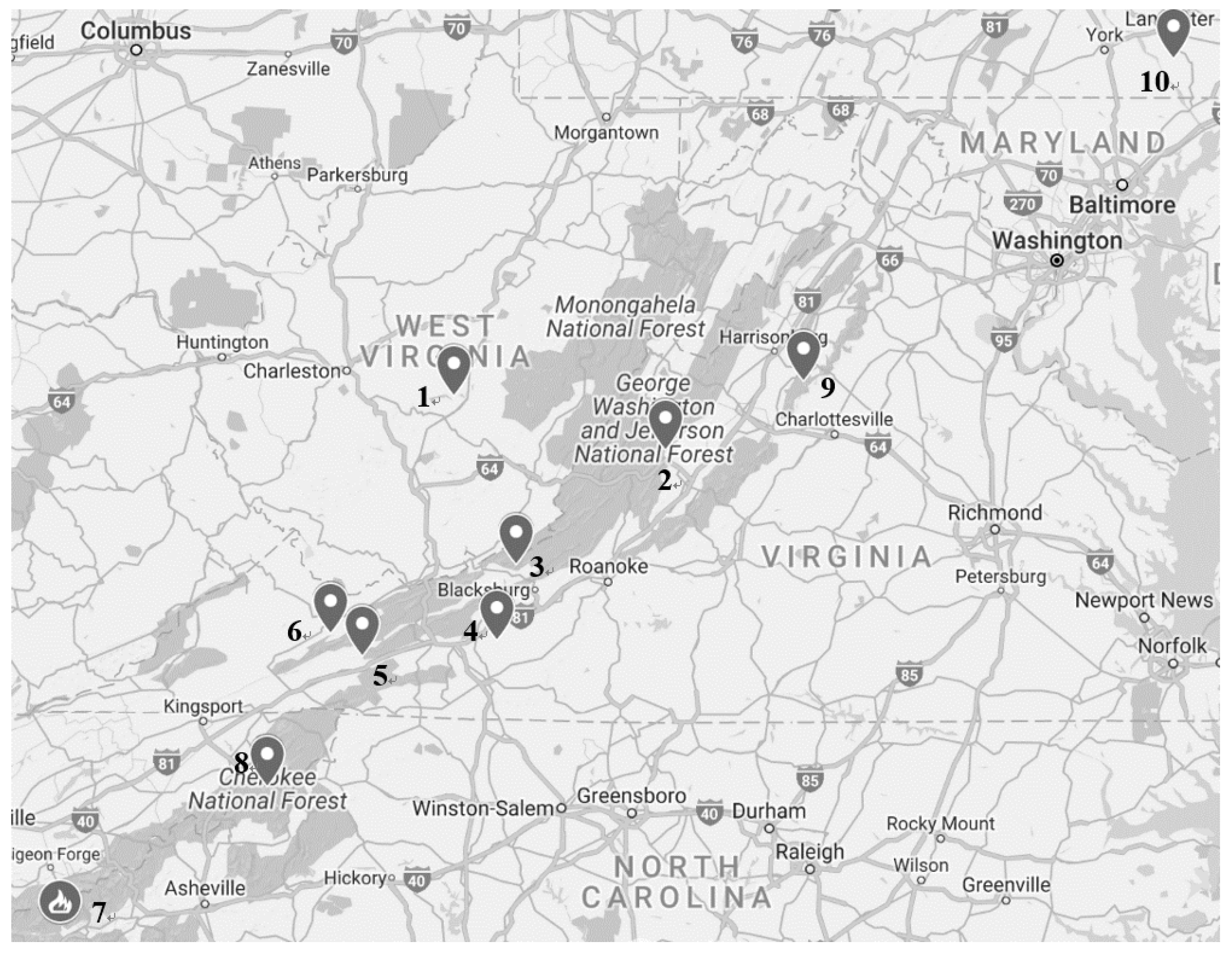

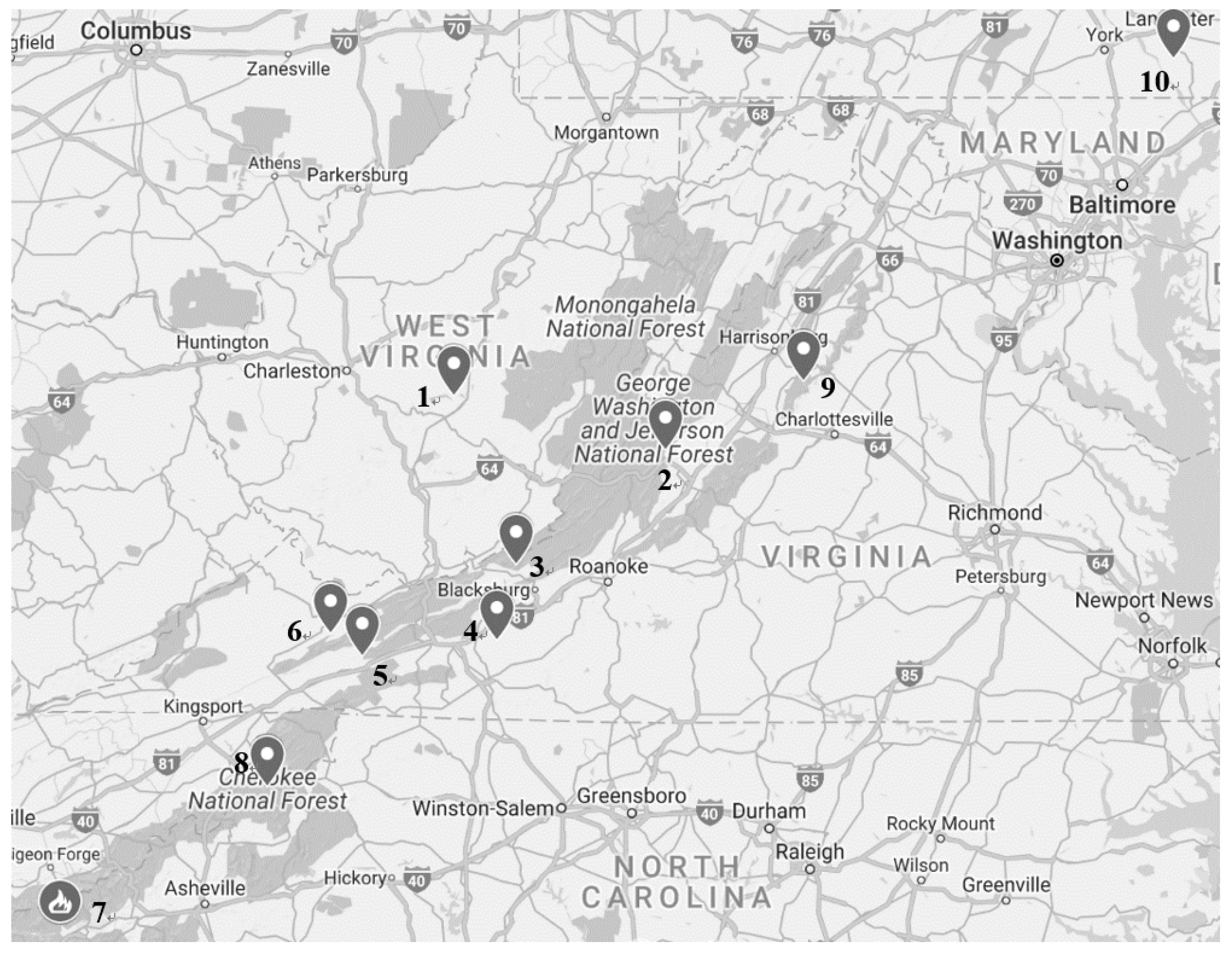{kind=link}
| Site | Year | Average % Live Crown Ratio | Average % Live Branches | Average % Tips Alive | Average % New Growth | Average % Crown Density | Tree Health Index (THI) 1,2 |
|---|---|---|---|---|---|---|---|
| Mountain Lake (VA) | 2015 | 88.8 ± 10.3 | 78.8 ± 16.5 | 78.8 ± 16.5 | 39.4 ± 14.8 | 87.5 ± 2.9 | 74.7 ± 20.3 AB |
| Carnifex, WV | 2015 | 90.0 ± 10.0 | 85.0 ± 8.7 | 86.7 ± 15.3 | 86.7 ± 15.3 | 90.0 ± 10.0 | 87.7 ± 2.2 A |
| 2016 | 95.0 ± 7.1 | 90.0 ± 14.1 | 90.0 ± 14.1 | 86.7 ± 15.3 | 95.0 ± 7.1 | 91.3 ± 3.6 a | |
| Saltville (VA) | 2015 | 63.8 ± 11.9 | 65.0 ± 12.0 | 66.9 ± 11.6 | 48.8 ± 3.5 | 50.0 ± 16.0 | 58.9 ± 8.8 B |
| 2016 | 60 ± 10.7 | 66.25 ± 13.0 | 68.75 ± 15.5 | 37.5 ± 3.5 | 50 ± 19.1 | 56.5 ± 12.9 b | |
| Goshen (VA) | 2015 | 55.6 ± 15.9 | 62.5 ± 16.9 | 66.3 ± 16.0 | 56.3 ± 30.7 | 55.6 ± 12.7 | 59.3 ± 4.9 B |
| 2016 | 72.0 ± 10.9 | 66.0 ± 8.9 | 72.0 ± 4.5 | 20.0 ± 0.0 | 68.0 ± 8.4 | 59.6 ± 22.3 b | |
| Hungry Mother State Park (VA) (1st/2nd) | 2015 | 64.3 ± 11.3 | 61.4 ± 10.7 | 55.7 ± 7.9 | 41.4 ± 10.7 | 54.3 ± 18.1 | 55.4 ± 8.8 B |
| 2016 | 65.7 ± 18.1 | 62.9 ± 17.0 | 57.1 ± 7.6 | 10.0 ± 0.0 | 57.1 ± 24.3 | 50.6 ± 23.0 b | |
| Powhatan Boy Scout Camp (VA) | 2015 | 22.9 ± 12.2 | 36.4 ± 17.0 | 33.6 ± 19.1 | 3.9 ± 2.8 | 17.0 ± 9.5 | 22.8 ± 13.2 C |
| 2016 | 35.0 ± 22.0 | 41.3 ± 18.7 | 40.0 ± 15.6 | 7.5 ± 10.3 | 31.0 ± 16.4 | 31.0 ± 13.7 c | |
| Shenandoah National Park (VA) | 2015 | 84.0 ± 15.2 | 77.0 ± 8.4 | 72.0 ± 13.0 | 52.0 ± 17.9 | 64.0 ± 11.4 | 69.8 ± 12.3 AB |
| 2016 | 64.0 ± 20.7 | 74.0 ± 13.4 | 62.0 ± 4.5 | 20.0 ± 0.0 | 54.0 ± 24.1 | 54.8 ± 20.7 b |
| Site | Year of Release | Number Released | Plant Hardiness Zone 2 | Field Season | HWA Density (adelgid/cm) | Laricobius Adults (Percent Estimated L. osakensis) 3 | Laricobius osakensis Larvae | Laricobius nigrinus Larvae | Laricobius rubidus Larvae |
|---|---|---|---|---|---|---|---|---|---|
| Mountain Lake (VA) | 2012 | 500 | 5b | 2015–2017 | ~0 | 0 | 0 | 0 | 0 |
| Carnifex (WV) | 2012 | 500 | 6a | 2015–2017 | ~0 | 0 | 0 | 0 | 0 |
| Saltville (VA) | 2013 | 2000 | 5b | 2015–2017 | ~0 | 0 | 0 | 0 | 0 |
| Goshen (VA) | 2013 | 2000 | 6b | 2015–2016 | 1.4 ± 1.0 | 0 | 0 | 0 | 7 |
| 2016–2017 | 1.1 ± 0.6 | 0 | 21 | 2 | 38 | ||||
| Hungry Mother (VA) 4 | 2014/2015 | 617/300 | 6b | 2015–2016 | 1.7 ± 0.8 | 3 (80%) | 8 | 2 | 0 |
| 2016–2017 | 1.6 ± 0.8 | 12 (13%) | 12 | 62 | 16 | ||||
| Powhatan Boy Scout Camp (VA) | 2014 | 1000 | 6b | 2015–2016 | 1.0 ± 0.4 | 0 | 2 | 0 | 26 |
| 2016–2017 | 1.2 ± 0.6 | 3 (19%) | 11 | 39 | 8 | ||||
| Shenandoah National Park (VA) 1 | 2015 | 500 | 6b | 2015–2016 | 1.9 ± 0.7 | 0 | 2 | 3 | 3 |
| 2016–2017 | 1.8 ± 0.5 | 1 | 0 | 0 | 0 | ||||
| Great Smoky Mountain National Park (TN) | 2014 | 921 | 6b | 2015–2016 | 1.3 ± 0.3 | 0 | 2 | 0 | 0 |
| Trouts Run (PA) 1 | 2015 | 1021 | 6a | 2016–2017 | 1.6 ± 0.4 | 41 (100%) | 46 | 0 | 0 |
| South Cherokee National Forest (TN) | 2014 | 781 | 7a | 2015–2016 | 1.9 ± 0.6 | 0 | 0 | 0 | 5 |
| 2016–2017 | 1.8 ± 0.6 | 0 | 0 | 0 | 0 |
| Factors | Spearman ρ 1 | p-Value |
|---|---|---|
| Laricobius spp. larvae and adults & HWA density | 0.48 | 0.042 |
| L. osakensis larvae & HWA density | 0.29 | 0.247 |
| L. osakensis larvae & THI | −0.54 | 0.045 |
| L. osakensis larvae & Plant hardiness zone | 0.21 | 0.412 |
| L. osakensis larvae & Number of beetles released | 0.19 | 0.447 |
| HWA density & Plant hardiness zone | 0.83 | 0.001 |
| HWA density & THI | −0.50 | 0.066 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toland, A.; Brewster, C.; Mooneyham, K.; Salom, S. First Report on Establishment of Laricobius osakensis (Coleoptera: Derodontidae), a Biological Control Agent for Hemlock Woolly Adelgid, Adelges tsugae (Hemiptera: Adelgidae), in the Eastern U.S. Forests 2018, 9, 496. https://doi.org/10.3390/f9080496
Toland A, Brewster C, Mooneyham K, Salom S. First Report on Establishment of Laricobius osakensis (Coleoptera: Derodontidae), a Biological Control Agent for Hemlock Woolly Adelgid, Adelges tsugae (Hemiptera: Adelgidae), in the Eastern U.S. Forests. 2018; 9(8):496. https://doi.org/10.3390/f9080496
Chicago/Turabian StyleToland, Ashley, Carlyle Brewster, Katlin Mooneyham, and Scott Salom. 2018. "First Report on Establishment of Laricobius osakensis (Coleoptera: Derodontidae), a Biological Control Agent for Hemlock Woolly Adelgid, Adelges tsugae (Hemiptera: Adelgidae), in the Eastern U.S." Forests 9, no. 8: 496. https://doi.org/10.3390/f9080496
APA StyleToland, A., Brewster, C., Mooneyham, K., & Salom, S. (2018). First Report on Establishment of Laricobius osakensis (Coleoptera: Derodontidae), a Biological Control Agent for Hemlock Woolly Adelgid, Adelges tsugae (Hemiptera: Adelgidae), in the Eastern U.S. Forests, 9(8), 496. https://doi.org/10.3390/f9080496