Hierarchical Environmental Factors Affecting the Distribution of Abies koreana on the Korean Peninsula


Abstract
1. Introduction
2. Materials and Methods
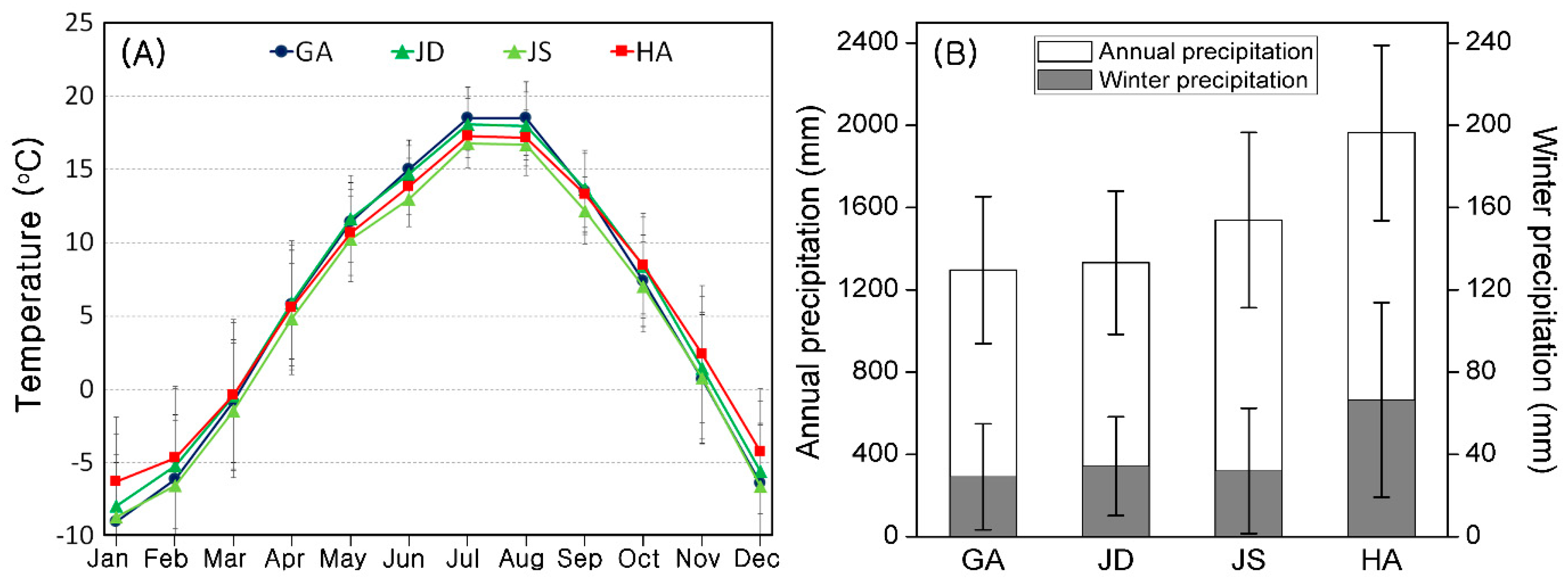
2.1. Study Sites
2.2. Sampling Design and Species Data Collection
2.3. Environmental Data Collection
2.4. Data Analysis
3. Results
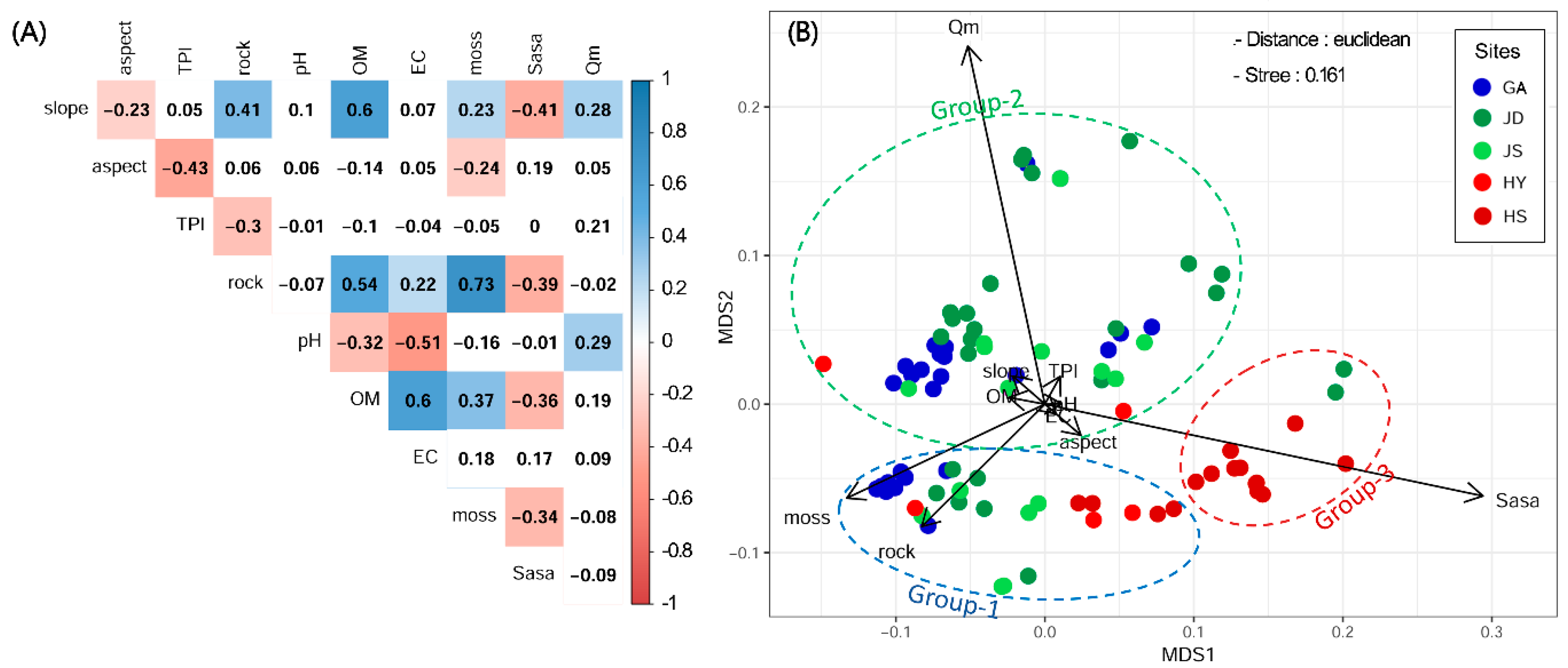
3.1. Comparison of Study Sites
3.2. The Relationship among Environmental Variables
3.3. Korean Fir Seedling: Environmental Factor Relationships
3.4. The Density of Korean Fir: Environmental Factors Relationships
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dale, M.R.T. Spatial Pattern Analysis in Plant Ecology; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Moeslund, J.E.; Arge, L.; Bøcher, P.K.; Dalgaard, T.; Svenning, J.C. Topography as a driver of local terrestrial vascular plant diversity patterns. Nord. J. Bot. 2013, 31, 129–144. [Google Scholar] [CrossRef]
- Lippok, D.; Beck, S.G.; Renison, D.; Hensen, I.; Apaza, A.E.; Schleuning, M. Topography and edge effects are more important than elevation as drivers of vegetation patterns in a neotropical montane forest. J. Veg. Sci. 2014, 25, 724–733. [Google Scholar] [CrossRef]
- Opedal, Ø.H.; Armbruster, W.S.; Graae, B.J. Linking small-scale topography with microclimate, plant species diversity and intra-specific trait variation in an alpine landscape. Plant Ecol. Divers. 2015, 8, 305–315. [Google Scholar] [CrossRef]
- Yasuhiro, K.; Hirofumi, M.; Kihachiro, K. Effects of topographic heterogeneity on tree species richness and stand dynamics in a subtropical forest in Okinawa Island, southern Japan. J. Ecol. 2004, 92, 230–240. [Google Scholar] [CrossRef]
- Shi, H.; Wen, Z.; Guo, M. Leaf Trait Variation with Environmental Factors at Different Spatial Scales: A Multilevel Analysis across a Forest-Steppe Transition. Forests 2018, 9, 122–135. [Google Scholar]
- McMahon, S.M.; Diez, J.M. Scales of association: Hierarchical linear models and the measurement of ecological systems. Ecol. Lett. 2007, 10, 437–452. [Google Scholar] [CrossRef] [PubMed]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Kéry, M.; Royle, J.A. Applied Hierarchical Modeling in Ecology; Elsevier: Amsterdam, The Netherland, 2016. [Google Scholar]
- Lee, T.B. Illustrated Flora of Korea; Hyangmunsa: Seoul, Korea, 1999. [Google Scholar]
- Koo, K.A.; Park, W.K.; Kong, W.S. Dendrochronological analysis of Abies koreana W. at Mt. Halla, Korea: Effects of climate change on the growths. Korean J. Ecol. 2001, 24, 281–288. [Google Scholar]
- Rochefort, R.M.; Little, R.L.; Woodward, A.; Peterson, D.L. Changes in sub-alpine tree distribution in western North America: A review of climatic and other causal factors. Holocene 1994, 4, 89–100. [Google Scholar] [CrossRef]
- Koo, K.A.; Kong, W.S.; Park, S.U.; Lee, J.H.; Kim, J.; Jung, H. Sensitivity of Korean fir (Abies koreana Wils.), a threatened climate relict species, to increasing temperature at an island subalpine area. Ecol. Model. 2017, 353, 5–16. [Google Scholar] [CrossRef]
- Lim, J.H.; Woo, S.Y.; Kwon, M.J.; Chun, J.H.; Shin, J.H. Photosynthetic Capacity and Water Use Efficiency under Different Temperature Regimes on Healthy and Declining Korean Fir in Mt. Halla. J. Korean For. Sci. 2006, 95, 705–710. [Google Scholar]
- Tsuyama, I.; Higa, M.; Nakao, K.; Matsui, T.; Horikawa, M.; Tanaka, N. How will subalpine conifer distributions be affected by climate change? Impact assessment for spatial conservation planning. Reg. Environ. Chang. 2015, 15, 393–404. [Google Scholar] [CrossRef]
- Hwang, J.E.; Kim, Y.J.; Shin, M.H.; Hyun, H.J.; Bohnert, H.J.; Park, H.C. A comprehensive analysis of the Korean fir (Abies koreana) genes expressed under heat stress using transcriptome analysis. Sci. Rep. 2018, 8, 10233. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.Y.; Lim, J.H.; Lee, D.K. Effects of temperature on photosynthetic rates in Korean fir (Abies koreana) between healthy and dieback population. J. Integr. Plant Boil. 2008, 50, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Gonzalez, P.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Han, A.R.; Kim, H.J.; Jung, J.B.; Park, P.S. Seed germination and initial seedling survival of the subalpine tree species, Picea jezoensis, on different forest floor substrates under elevated temperature. For. Ecol. Manag. 2018, 429, 579–588. [Google Scholar] [CrossRef]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/species/31244/9618913 (accessed on 2 June 2018).
- Song, K.M.; Kang, Y.J.; Hyeon, H.J. Vegetation Structure at the Slope Direction and Characteristic of Seedlings of Abies koreana in Hallasan Mountain. J. Environ. Sci. Int. 2014, 3, 39–46. [Google Scholar] [CrossRef]
- Kim, J.; Park, E. Chemical and Biological Properties of the Genus Abies. In Advances in Plant Phenolics: From Chemistry to Human Health; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2018; pp. 225–236. ISBN 978-0-8412-3295-2. [Google Scholar]
- Korea Institute of Geoscience and Mineral Resources. 2018. Geologic Maps. Available online: https://mgeo.kigam.re.kr/map/geology.jsp (accessed on 27 June 2018).
- Lee, C.S.; Cho, H.J. Structure and Dynamics of Abies koreana Wilson Community in Mt. Gaya. Korean J. Ecol. 1993, 16, 75–91. [Google Scholar]
- Cho, D.S. Community Structure, and Size and Age Distribution of Conifers in Subalpine Korean Fir (Abies koreana) Forest in Mt. Chiri. Korean J. Ecol. 1994, 17, 415–424. [Google Scholar]
- Kim, G.T.; Choo, G.C.; Um, T.W. Studies on the Structure of Abies koreana Community at Subalpine Zone in Hallasan. Korea J. Environ. Ecol. 2007, 21, 161–167. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. R Core Team (2014) nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-137 2014. Available online: http://CRAN.r-project.org/package=nlme (accessed on 2 June 2018).
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; Stevens, M.; Wagner, H. Vegan: Community Ecology Package. R Package Version. 2.5-2. CRAN. Available online: http://CRAN.r-project.org/package=vegan (accessed on 10 June 2018).
- Finch, W.H.; Bolin, J.E.; Kelley, K. Multilevel Modeling Using R; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Washitani, I.; Kabaya, H. Germination responses to temperature responsible for the seedling emergence seasonality of Primula sieboldii E. Morren in its natural habitat. Ecol. Res. 1988, 3, 9–20. [Google Scholar] [CrossRef]
- Cho, H.K.; Takahashi, K.; Shibuya, M.; Saito, H.; Kim, J.J.; Hong, S.G. Effects of cold stratification, temperature, and drought on seed germination characteristics of Abies koreana and Abies sachalinensis. J. Jpn. For. Soc. 2005, 87, 465–470. [Google Scholar] [CrossRef]
- Kim, J.J.; Cho, H.K.; Takahashi, K. Environmental Characteristics of Abies koreana Stand on Mt. Halla. Korean J. Nat. Conserv. 2007, 1, 5–9. [Google Scholar]
- Gratzer, G.; Rai, P.B.; Glatzel, G. The influence of the bamboo Yushania microphylla on regeneration of Abies densa in central Bhutan. Can. J. For. Res. 1999, 29, 1518–1527. [Google Scholar] [CrossRef]
- Park, S.G.; Yi, M.H.; Yoon, J.W.; Sin, H.T. Environmental factors and growth properties of Sasa borealis (Hack.) Makino community and effect its distribution on the development of lower vegetation in Jirisan National Park. Korean J. Environ. Ecol. 2012, 26, 82–90. [Google Scholar]
- Yamamoto, S.I. Gap characteristics and gap regeneration in a subalpine coniferous forest on Mt Ontake, central Honshu. Jpn. Ecol. Res. 1993, 8, 277–285. [Google Scholar] [CrossRef]
- Narukawa, Y.; Yamamoto, S. Effects of dwarf bamboo (Sasa sp.) and forest floor microsites on conifer seedling recruitment in a subalpine forest, Japan. For. Ecol. Manag. 2002, 163, 61–70. [Google Scholar] [CrossRef]
- Takahashi, K. Regeneration and coexistence of two subalpine conifer species in relation to dwarf bamboo in the understorey. J. Veg. Sci. 1997, 8, 529–536. [Google Scholar] [CrossRef]
- Germino, M.J.; Smith, W.K.; Resor, A.C. Conifer seedling distribution and survival in an alpine-tree line ecotone. Plant Ecol. 2002, 162, 157–168. [Google Scholar] [CrossRef]
- Cui, M.; Smith, W.K. Photosynthesis, water relations and mortality in Abies lasiocarpa seedlings during natural establishment. Tree Physiol. 1991, 8, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T. Effect of bryophytes on survival of conifer seedlings in subalpine forests of central Japan. Ecol. Res. 1992, 7, 155–162. [Google Scholar] [CrossRef]
- Karger, D.N.; Kluge, J.; Abrahamczyk, S.; Salazar, L.; Homeier, J.; Lehnert, M.; Amoroso, V.B.; Kessler, M. Bryophyte cover on trees as proxy for air humidity in the tropics. Ecol. Indic. 2012, 20, 277–281. [Google Scholar] [CrossRef]
- Domisch, T.; Martz, F.; Repo, T.; Rautio, P. Winter survival of Scots pine seedlings under different snow conditions. Tree Physiol. 2017, 38, 602–616. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Byun, H.R. Future pattern of Asian drought under global warming scenario. Theor. Appl. Climatol. 2009, 98, 137–150. [Google Scholar] [CrossRef]
- Jang, W.S.; Park, P.S.; Han, A.R.; Kim, K.Y.; Kim, M.P.; Park, H.K. The spatial distribution of Quercus mongolica and its association with other tree species in two Quercus mongolica stands in Mt. Jiri, Korea. J. Ecol. Environ. 2010, 33, 67–77. [Google Scholar] [CrossRef]
- Gosling, S.N.; Dunn, R.; Carrol, F.; Christidis, N.; Fullwood, J.; de Gusmao, D.; Golding, N.; Good, L.; Hall, T.; Kendon, L. Climate: Observations, Projections and Impacts. UK Met Office. Nottingham ePrints: Nottingham, UK, 2011. Available online: http://eprints.nottingham.ac.uk/2040/ (accessed on 12 June 2018).
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report; Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Streb, P.; Aubert, S.; Bligny, R. High temperature effects on light sensitivity in the two high mountain plant species Soldanella alpina (L.) and Rannunculus glacialis (L.). Plant Biol. 2003, 5, 432–440. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| N | GA | JD | JS | HY | HS | F Value | |
|---|---|---|---|---|---|---|---|
| Living Abies koreana | 102 | 7.2 ± 1.4 c | 12 ± 1.8 bc | 20.8 ± 2.5 a | 12.3 ± 1.1 bc | 15.5 ± 1.5 ab | 8.4 * |
| Dead A. koreana | 84 | 0.2 ± 0.1 b | 4.6 ± 0.9 a | 4.3 ± 0.6 a | 4.61 ± 0.8 a | 2.5 ± 0.8 ab | 10.1 * |
| A. koreana seedling | 84 | 3.6 ± 1.0 | 2.1 ± 0.9 | 4.9 ± 1.7 | 6.0 ± 1.3 | 2.5 ± 1.2 | 1.2 |
| Elevation (m) | 102 | 1354.0 ± 4 e | 1401 ± 19 d | 1457 ± 19 c | 1615 ± 4 b | 1681 ± 5 a | 118 * |
| Aspect (1–4) | 102 | 2.3 ± 0.2 b | 1.9 ± 0.2 bc | 3.6 ± 0.1 a | 1.4 ± 0.1 c | 3.1 ± 0.2 a | 18.4 * |
| Slope (°) | 102 | 31.0 ± 1.9 a | 18.4 ± 0.5 b | 13.5 ± 1.4 c | 8.3 ± 0.7 d | 9.5 ± 0.7 cd | 64.1 * |
| TPI (1–3) | 102 | 2.1 ± 0.2 bc | 2.8 ± 0.1 a | 1.5 ± 0.2 c | 2.3 ± 0.1 ab | 1.9 ± 0.2 bc | 8.2 * |
| Rock cover (0–5) | 102 | 4.0 ± 0.3 a | 1.1 ± 0.2 b | 3.1 ± 0.4 a | 3.7 ± 0.2 a | 1.5 ± 0.4 b | 23.3 * |
| pH | 84 | 4.6 ± 0.1 | 4.6 ± 0 | 4.5 ± 0.1 | 4.3 ± 0.1 | 4.6 ± 0.0 | 1.4 |
| OM (%) | 84 | 36.3 ± 1.6 a | 17.5 ± 1.2 b | 18.4 ± 1 b | 20.1 ± 4.3 b | 14.0 ± 1.6 b | 37.5 * |
| EC (dS/m) | 84 | 0.8 ± 0.1 a | 0.4 ± 0.0 b | 0.5 ± 0.1 b | 1.0 ± 0.2 a | 0.7 ± 0.1 ab | 10.1 * |
| Sasa cover (0–5) | 102 | 0.7 ± 0.3 b | 1.2 ± 0.4 b | 1.1 ± 0.4 b | 3.2 ± 0.5 a | 4.8 ± 0.1 a | 16.9 * |
| Moss cover (0–5) | 102 | 2.3 ± 0.2 ab | 0.8 ± 0.2 c | 1.5 ± 0.3 bc | 2.9 ± 0.2 a | 0.8 ± 0.4 c | 14.6 * |
| Qm cover (0–5) | 102 | 1.8 ± 0.3 a | 1.5 ± 0.2 ab | 1.4 ± 0.3 ab | 0.6 ± 0.2 bc | 0.0 ± 0.0 c | 7.3 * |
| Unconditional model | ||||||
| Fixed Effect | Random Effect | |||||
| Parameter | Estimate | z value | p-value | Variance | AIC | |
| Intercept | 0.16 | 0.415 | 0.67 | 0.39 | 119 | |
| Univariate random coefficient model () | ||||||
| (1) | Intercept | −0.50 | −1.27 | 0.20 | 0.001 | 110 |
| Slope | −0.06 | −0.61 | 0.54 | 0.046 | ||
| (2) | Intercept | −0.17 | −0.55 | 0.58 | 0.036 | 114 |
| Aspect | −0.63 | −1.05 | 0.29 | 1.241 | ||
| (3) | Intercept | 0.15 | 0.37 | 0.71 | 0.386 | 124 |
| TPI | −0.01 | −0.04 | 0.97 | 0.008 | ||
| (4) | Intercept | 0.03 | 0.36 | 0.92 | 0.240 | 111 |
| Rock cover | 0.48 ** | 2.30 | <0.05 | 0.078 | ||
| (5) | Intercept | 0.15 | 0.39 | 0.69 | 0.407 | 123 |
| pH | −0.50 | −0.40 | 0.69 | 1.961 | ||
| (6) | Intercept | 0.16 | 0.45 | 0.65 | 0.256 | 121 |
| OM | 0.03 | 1.13 | 0.26 | 0.001 | ||
| (7) | Intercept | 0.31 | 0.74 | 0.46 | 0.371 | 120 |
| EC | 0.02 | 0.01 | 0.99 | 7.052 | ||
| (8) | Intercept | −0.13 | 0.39 | 0.72 | 0.195 | 104 |
| Moss cover | 1.01 ** | 2.27 | <0.05 | 0.514 | ||
| (9) | Intercept | 0.31 | 0.50 | 0.62 | 0.889 | 109 |
| Sasa cover | −0.56 ** | −2.57 | <0.05 | 0.002 | ||
| Final hierarchical logistic regression model () | ||||||
| Intercept | 0.02 * | 0.06 | 0.07 | 0.036 | 97 | |
| Moss cover | 0.76 * | 1.82 | 0.06 | 0.406 | ||
| Sasa cover | −0.68 ** | −2.53 | <0.05 | - | ||
| Pw | 0.08 ** | 1.96 | <0.05 | - | ||
| Unconditional model ( | ||||||
| Fixed Effect | Random Effect | |||||
| Parameter | Estimate | z value | p-value | Variance | AIC | |
| Intercept | 2.59 *** | 16.1 | <0.001 | 0.13 | 757 | |
| Univariate Random coefficient model () | ||||||
| (1) | Intercept | 2.34 *** | 7.35 | <0.001 | 0.39 | 729 |
| Slope | 0.006 | 0.42 | 0.67 | 0.001 | ||
| (2) | Intercept | 2.56 *** | 9.57 | <0.001 | 0.27 | 734 |
| Aspect | −0.17 * | −2.05 | <0.05 | 0.02 | ||
| (3) | Intercept | 2.54 *** | 13.92 | <0.001 | 0.13 | 754 |
| TPI | −0.11 * | −2.54 | <0.05 | 0.003 | ||
| (4) | Intercept | 2.28 *** | 5.57 | <0.001 | 0.65 | 712 |
| Rock cover | 0.07 | 0.57 | 0.56 | 0.07 | ||
| (5) | Intercept | 2.55 *** | 12.47 | <0.001 | 0.16 | 738 |
| pH | 0.03 | 0.08 | 0.95 | 0.62 | ||
| (6) | Intercept | 2.34 *** | 6.32 | <0.001 | 0.53 | 699 |
| OM | 0.02 | 0.87 | 0.38 | 0.002 | ||
| (7) | Intercept | 2.78 *** | 9.21 | <0.001 | 0.33 | 727 |
| EC | −0.27 | −0.58 | 0.56 | 0.79 | ||
| (8) | Intercept | 2.47 *** | 10.04 | <0.001 | 0.07 | 702 |
| Qm cover | −0.17• | −1.89 | 0.06 | 0.03 | ||
| Final hierarchical Poisson regression model | ||||||
| Intercept | 2.66 *** | 46.79 | <0.001 | 0.006 | ||
| Aspect | −0.14 ** | −3.25 | <0.01 | - | ||
| TPI | −0.11 * | −2.03 | <0.05 | - | 688 | |
| Qm cover | −0.17• | −1.78 | 0.08 | 0.022 | ||
| Tsu | −0.30 *** | −5.13 | <0.001 | - | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.S.; Shin, H.S.; Choi, C.-h.; Lee, J.; Kim, J. Hierarchical Environmental Factors Affecting the Distribution of Abies koreana on the Korean Peninsula. Forests 2018, 9, 777. https://doi.org/10.3390/f9120777
Park JS, Shin HS, Choi C-h, Lee J, Kim J. Hierarchical Environmental Factors Affecting the Distribution of Abies koreana on the Korean Peninsula. Forests. 2018; 9(12):777. https://doi.org/10.3390/f9120777
Chicago/Turabian StylePark, Jeong Soo, Hak Sub Shin, Chul-hyun Choi, Junghyo Lee, and Jinhee Kim. 2018. "Hierarchical Environmental Factors Affecting the Distribution of Abies koreana on the Korean Peninsula" Forests 9, no. 12: 777. https://doi.org/10.3390/f9120777
APA StylePark, J. S., Shin, H. S., Choi, C.-h., Lee, J., & Kim, J. (2018). Hierarchical Environmental Factors Affecting the Distribution of Abies koreana on the Korean Peninsula. Forests, 9(12), 777. https://doi.org/10.3390/f9120777