Stand Volume Production in the Subsequent Stand during Three Decades Remains Unaffected by Slash and Stump Harvest in Nordic Forests



Abstract
1. Introduction
- Stand volume production and site productivity increases after stump harvest due to increased nutrient availability as a result of soil disturbance but decreases after slash harvest as the nutrient rich needles and branches are removed;
- Survival of planted seedlings increases after stump harvest due to increased nutrient availability and reduced vegetation competition;
- Recruitment of trees through natural regeneration increases after both stump and slash harvest, resulting in an even higher increase after combined stump and slash harvest. This increase is due to greater mineral soil exposure and reduced vegetation competition after stump and slash harvest.
2. Materials and Methods
2.1. Study Sites
2.2. Experimental Design
2.3. Measurements of Stand Characteristics
2.3.1. Seedling Establishment and Growth
2.3.2. Stand Volume Production
2.4. Statistical Analyses
3. Results
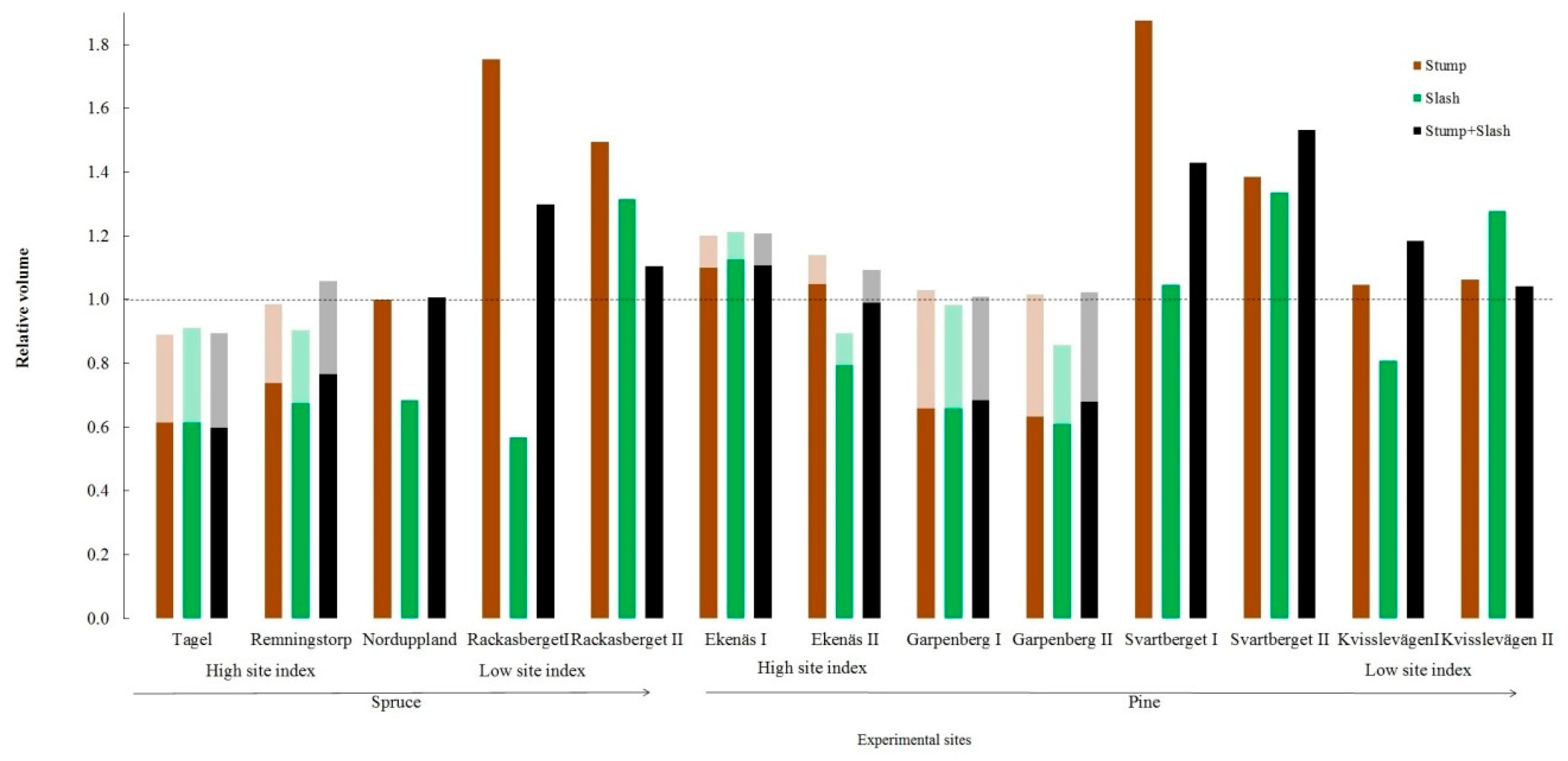
3.1. Stand Volume Production
3.2. Height Growth
3.3. Seedling Survival
3.4. Natural Regeneration
3.5. Species
4. Discussion
4.1. Stand Volume Production
4.2. Height Growth as a Proxy for Site Productivity
4.3. Seedling Survival
4.4. Natural Regeneration
5. Conclusions and Practical Implications
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Edenhofer, O.; Pichs-Madruga, R.; Sokona, Y.; Seyboth, K.; Matschoss, P.; Kadner, S.; Zwickel, T.; Eickemeier, P.; Hansen, G.; Schloemer, S.; et al. Renewable Energy Sources and Climate Change Mitigation, Special Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2011. [Google Scholar]
- Routa, J.; Asikainen, A.; Bjorheden, R.; Laitila, J.; Röser, D. Forest energy procurement: State of the art in Finland and Sweden. WIREs Energy Environ. 2013, 2, 602–613. [Google Scholar] [CrossRef]
- Börjesson, P.; Hansson, J.; Berndes, G. Future demand for forest-based biomass for energy purposes in Sweden. For. Ecol. Manag. 2017, 383, 17–26. [Google Scholar] [CrossRef]
- Walmsley, J.D.; Jones, D.L.; Reynolds, B.; Price, M.H.; Healey, J.R. Whole tree harvesting can reduce second rotation forest productivity. For. Ecol. Manag. 2009, 257, 1104–1111. [Google Scholar] [CrossRef]
- Grelle, A.; Strömgren, M.; Hyvönen, R. Carbon balance of a forest ecosystem after stump harvest. Scand. J. For. Res. 2012, 27, 762–773. [Google Scholar] [CrossRef]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage: Meta analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Zanchi, G.; Pena, N.; Bird, D.N. Is woody bioenergy carbon neutral? A comparative assessment of emissions from consumption of woody bioenergy and fossil fuel. GCB Bioenergy 2012, 4, 761–772. [Google Scholar] [CrossRef]
- Olsson, B.A.; Bengtsson, J.; Lundkvist, H. Effects of different forest harvest intensities on the pools of exchangeable cations in coniferous forest soils. For. Ecol. Manag. 1996, 84, 135–147. [Google Scholar] [CrossRef]
- Jonsell, M.; Hansson, J.; Wedmo, L. Diversity of saproxylic beetle species in logging residues in Sweden—Comparisons between tree species and diameters. Biol. Conserv. 2007, 138, 89–99. [Google Scholar] [CrossRef]
- Eklöf, K.; Meili, M.; Akerblom, S.; Von Brömssen, C.; Bishop, K. Impact of stump harvest on run-off concentrations of total mercury and methylmercury. For. Ecol. Manag. 2013, 290, 83–94. [Google Scholar] [CrossRef]
- Albaugh, T.J.; Albaugh, J.M.; Foxa, T.R.; Allen, H.L.; Rubilar, R.A.; Trichet, P.; Loustau, D.; Linder, S. Tamm Review: Light use efficiency and carbon storage in nutrient and water experiments on major forest plantation species. For. Ecol. Manag. 2016, 376, 333–342. [Google Scholar] [CrossRef]
- Ingestad, T. Towards optimum nutrition. Ambio 1974, 3, 49–54. [Google Scholar]
- Landsberg, J.J.; Waring, R.H. A generalised model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning. For. Ecol. Manag. 1997, 95, 209–228. [Google Scholar] [CrossRef]
- Tamm, C.O. Nitrogen in Terrestrial Ecosystems, Questions of Productivity, Vegetational Changes, and Ecosystem Stability; Springer: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Boyle, J.R.; Phillips, J.J.; Ek, A.R. Whole-tree harvesting: Nutrient budget evaluation. J. For. 1973, 71, 760–762. [Google Scholar]
- Carey, M.L. Whole-tree harvesting in Sitka spruce, possibilities and implications. Ir. For. 1980, 37, 48–63. [Google Scholar]
- Freedman, B.; Morash, R.; Hanson, A.J. Biomass and nutrient removals by conventional and whole-tree clear-cutting of a red spruce-balsam fir stand in central Nova Scotia. Can. J. For. Res. 1981, 11, 249–257. [Google Scholar] [CrossRef]
- Mälkönen, E. Effect of whole-tree harvesting on soil fertility. Silva Fenn. 1976, 10, 157–164. [Google Scholar] [CrossRef]
- Son, Y.; Gower, S.T. Nitrogen and phosphorus distribution for 5 plantation species in Southwestern Wisconsin. For. Ecol. Manag. 1992, 53, 175–193. [Google Scholar] [CrossRef]
- Hellsten, S.; Helmisaari, H.-S.; Melin, Y.; Skovsgaard, J.P.; Kaakinen, S.; Kukkola, M.; Saarsalmi, A.; Petersson, H.; Akselsson, C. Nutrient concentrations in stumps and coarse roots of Norway spruce, Scots pine and silver birch in Sweden, Finland and Denmark. For. Ecol. Manag. 2013, 290, 40–48. [Google Scholar] [CrossRef]
- Van Lear, D.H.; Kapeluck, P.R. Above- and below-stump biomass and nutrient content of a mature loblolly pine plantation. Can. J. For. 1995, 25, 361–367. [Google Scholar] [CrossRef]
- Bai, S.H.; Blumfield, T.J.; Reverchon, F. The impact of mulch type on soil organic carbon and nitrogen pools in a sloping site. Biol. Fertil. Soils 2014, 50, 37–44. [Google Scholar]
- Emmett, B.A.; Anderson, J.M.; Hornung, M. The controls on dissolved nitrogen losses following two intensities of harvesting in a Sitka spruce forest (N. Wales). For. Ecol. Manag. 1991, 41, 65–80. [Google Scholar] [CrossRef]
- Kataja-aho, S.; Smolander, A.; Fritze, H.; Norrgard, S. Responses of soil carbon and nitrogen transformations to stump removal. Silva Fenn. 2012, 46, 169–179. [Google Scholar] [CrossRef]
- Egnell, G. Is the productivity decline in Norway spruce following whole-tree harvesting in the final felling in boreal Sweden permanent or temporary? For. Ecol. Manag. 2011, 261, 148–153. [Google Scholar] [CrossRef]
- Wall, A.; Hytonen, J. The long-term effects of logging residue removal on forest floor nutrient capital, foliar chemistry and growth of a Norway spruce stand. Biomass Bioenergy 2011, 35, 3328–3334. [Google Scholar] [CrossRef]
- Egnell, G. Effects of slash and stump harvesting after final felling on stand and site productivity in Scots pine and Norway spruce. For. Ecol. Manag. 2016, 371, 42–49. [Google Scholar] [CrossRef]
- Nilsson, U.; Örlander, G. Vegetation management on grass-dominated clearcuts planted with Norway spruce in southern Sweden. Can. J. For. Res. 1999, 29, 1015–1026. [Google Scholar] [CrossRef]
- Örlander, G.; Egnell, G.; Albrektson, A. Long-term effects of site preparation on growth in Scots pine. For. Ecol. Manag. 1996, 86, 27–37. [Google Scholar] [CrossRef]
- Karlsson, K.; Tamminen, P. Long-term effects of stump harvesting on soil properties and tree growth in Scots pine and Norway spruce stands. Scand. J. For. Res. 2013, 8, 550–558. [Google Scholar] [CrossRef]
- Tarvainen, O.; Hekkala, A.-M.; Kubin, E.; Tamminen, P.; Murto, T.; Tolvanen, A. Soil disturbance and early vegetation response to varying intensity of energy wood harvest. For. Ecol. Manag. 2015, 348, 153–163. [Google Scholar] [CrossRef]
- Tamminen, P.; Saarsalmi, A. Effects of whole-tree harvesting on growth of pine and spruce seedlings in southern Finland. Scand. J. For. Res. 2013, 28, 559–565. [Google Scholar] [CrossRef]
- Smolander, A.; Saarsalmi, A.; Tamminen, P. Response of soil nutrient content, organic matter characteristics and growth of pine and spruce seedlings to logging residues. For. Ecol. Manag. 2015, 357, 117–125. [Google Scholar] [CrossRef]
- Karlsson, C.; Örlander, G. Soil scarification shortly before a rich seed fall improves seedling establishment in seed tree stands of Pinus sylvestris. Scand. J. For. Res. 2000, 15, 256–266. [Google Scholar] [CrossRef]
- Winsa, H. Influence of rain shelter and site preparation on seedling emergence and Pinus sylvestris L. after direct seeding. Scand. J. For. Res. 1995. [Google Scholar] [CrossRef]
- McInnis, B.G.; Roberts, M.R. The effects of full-tree and tree-length harvests on natural regeneration. North. J. Appl. For. 1994, 11, 131–137. [Google Scholar]
- Karlsson, M.; Nilsson, U.; Örlander, G. Natural regeneration in clear-cuts: Effects of scarification, slash removal and clear-cut age. Scand. J. For. Res. 2002, 17, 131–138. [Google Scholar] [CrossRef]
- Hyvönen, R.; Kaarakka, L.; Leppälammi-Kujansuu, J.; Olsson, B.A.; Palviainen, M.; Vegerfors-Persson, B.; Helmisaari, H.-S. Effects of stump harvesting on soil C and N stocks and vegetation 8–13 years after clear-cutting. For. Ecol. Manag. 2016, 371, 23–32. [Google Scholar] [CrossRef]
- Saksa, T. Regeneration after stump harvesting in southern Finland. For. Ecol. Manag. 2013, 290, 79–82. [Google Scholar] [CrossRef]
- Bergh, J.; Linder, S.; Lundmark, T.; Björn, E. The effect of water and nutrient availability on the productivity of Norway spruce in northern and southern Sweden. For. Ecol. Manag. 1999, 119, 51–62. [Google Scholar] [CrossRef]
- Jarvis, P.; Linder, S. Botany—Constraints to growth of boreal forests. Nature 2000, 405, 904–905. [Google Scholar] [CrossRef]
- Egnell, G.; Leijon, B. Survival and growth of planted seedlings of Pinus sylvestris and Picea abies after different levels of biomass removal in clear-felling. Scand. J. For. Res. 1999, 14, 303–311. [Google Scholar] [CrossRef]
- Hope, G.D. Changes in soil properties, tree growth, and nutrition over a period of 10 years after stump removal and scarification on moderately coarse soils in interior British Columbia. For. Ecol. Manag. 2007, 242, 625–635. [Google Scholar] [CrossRef]
- Kardell, L.; Wärne, C. Stubbar och Ris—Blåbär och Lingon. Utläggning av Skogsenergiförsök 1978–1980; Rapport 21 (in Swedish); SLU, Inst. för Skoglig Landskapsvård: Garpenberg, Sweden, 1981. [Google Scholar]
- Morén, A.-S.; Perttu, K. Regional temperature and radiation indices and their adjustment to horizontal and inclined forest land. Stud. For. Suec. 1994. [Google Scholar]
- Karlsson, G.; Hellsten, S.; Karlsson, P.; Akselsson, C.; Ferm, M. Kvävedepositionen till Sverige. Jämförelse av Depositionsdata från Krondroppsnätet, Luft-och Nederbördskemiska Nätet Samt EMEP, IVL Rapport B2030. Available online: http://www.krondroppsnatet.ivl.se/download/18.488d9cec137bbdebf94800056849/1350483726580/B2030.pdf (accessed on 25 October 2016).
- Lehtonen, A.; Mäkipää, R.; Heikkinen, J.; Sievänen, R.; Liski, J. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests. For. Ecol. Manag. 2004, 188, 211–224. [Google Scholar] [CrossRef]
- Karlsson, K.; Mossberg, M.; Ulvcrona, T. Fältdatasystem för Skogliga Fältförsök (in Swedish); Swedish University of Agricultural Sciences, Unit for Field-based Forest Research: Umeå, Sweden, 2012. [Google Scholar]
- Brandel, G. Volymfunktioner för Enskilda Träd. Tall, Gran och Björk; Report 26 (in Swedish); SLU, Inst for Skogsproduktion: Garpenberg, Sweden, 1990. [Google Scholar]
- Lundmark, R.; Athanassiadis, D.; Wetterlund, E. Supply assessment of forest biomass—A bottom-up approach for Sweden. Biomass Bioenergy 2015, 75, 213–226. [Google Scholar] [CrossRef]
- Proe, M.F.; Cameron, A.D.; Dutch, J.; Christodoulou, X.C. The effect of whole-tree harvesting on the growth of second rotation Sitka spruce. Forestry 1996, 69, 389–401. [Google Scholar] [CrossRef]
- Egnell, G. A review of Nordic trials studying effects of biomass harvest intensity on subsequent forest production. For. Ecol. Manag. 2017, 383, 27–36. [Google Scholar] [CrossRef]
- Ranius, T.; Hamalainen, A.; Egnell, G.; Olsson, B.; Eklof, K.; Stendahl, J.; Rudolphi, J.; Sténs, A.; Felton, A. The effects of logging residue extraction for energy on ecosystem services and biodiversity: A synthesis. J. Environ. Manag. 2018, 209, 409–425. [Google Scholar] [CrossRef]
- Powers, R.F. Long-term soil productivity: Genesis of the concept and principles behind the program. Can. J. For. Res. 2006, 36, 519–528. [Google Scholar] [CrossRef]
- Fleming, R.L.; Leblanc, J.D.; Hazlett, P.W.; Weldon, T.; Irwin, R.; Mossa, D.S. Effects of biomass harvest intensity and soil disturbance on jack pine stand productivity: 15-year results. Can. J. For. Res. 2014, 44, 1566–1574. [Google Scholar] [CrossRef]
- Morris, D.M.; Kwiaton, M.M.; Duckert, D.R. Black spruce growth response to varying levels of biomass harvest intensity across a range of soil types: 15-year results. Can. J. For. Res. 2014, 44, 313–325. [Google Scholar] [CrossRef]
- Achat, D.L.; Deleuze, C.; Landmann, G.; Pousse, N.; Ranger, J.; Augusto, L. Quantifying consequences of removing harvesting residues on forest soils and tree growth: A meta-analysis. For. Ecol. Manag. 2015, 348, 124–141. [Google Scholar] [CrossRef]
- Palviainen, M.; Finér, L.; Kurka, A.M.; Mannerkoski, H.; Piirainen, S.; Starr, M. Decomposition and nutrient release from logging residues after clear-cutting of mixed boreal forest. Plant Soil 2004, 263, 53–67. [Google Scholar] [CrossRef]
- Hyvönen, R.; Olsson, B.A.; Lundkvist, H.; Staaf, H. Decomposition and nutrient release from Picea abies (L.) Karst. and Pinus sylvestris L. logging residues. For. Ecol. Manag. 2000, 126, 97–112. [Google Scholar] [CrossRef]
- Nurmi, J. Recovery of logging residues for energy from spruce (Pices abies) dominated stands. Biomass Bioenergy 2007, 31, 375–380. [Google Scholar] [CrossRef]
- Peltola, S.; Kilpelainen, H.; Asikainen, A. Recovery rates of logging residue harvesting in Norway spruce (Picea abies (L.) Karsten) dominated stands. Biomass Bioenergy 2011, 35, 1545–1551. [Google Scholar] [CrossRef]
- Thiffault, E.; Bechard, A.; Paré, D.; Allen, D. Recovery rate of harvest residues for bioenergy in boreal and temperate forests: A review. Energ. Environ. 2015, 4, 429–451. [Google Scholar] [CrossRef]
- Linder, P.; Elfving, B.; Zackrisson, O. Stand structure and successional trends in virgin boreal forest reserves in Sweden. For. Ecol. Manag. 1997, 98, 17–33. [Google Scholar] [CrossRef]
- Hägglund, B.; Lundmark, J.E. Site index estimation by means of site properties—Scots pine and Norway spruce in Sweden. Stud. For. Suec. 1997, 138, 38. [Google Scholar]
- Thiffault, E.; Hannam, K.D.; Pare, D.; Titus, B.D.; Hazlett, P.W.; Maynard, D.G.; Brais, S. Effects of forest biomass harvesting on soil productivity in boreal and temperate forests—A review. Environ. Rev. 2011, 19, 278–309. [Google Scholar] [CrossRef]
- Saarinen, V.M. The effects of slash and stump removal on productivity and quality of forest regeneration operations-preliminary results. Biomass Bioenergy 2006, 30, 349–356. [Google Scholar] [CrossRef]
- SUAS Forest Statistics. Official Statistics of Sweden; Swedish University of Agricultural Sciences: Umeå, Sweden, 2015; ISSN 0280-0543. [Google Scholar]


{kind=link}
| Study Site | Tagel | Remningstorp | Norduppland | Rackasberget | Ekenäs | Garpenberg | Svartberget | Kvisslevägen |
|---|---|---|---|---|---|---|---|---|
| Former Stand | ||||||||
| Species composition (%) a | 0, 90, 10 | 50, 50, 0 | 70, 30, 0 | 0, 100, 0 | 80, 20, 0 | 50, 50, 0 | 10, 90, 0 | 40, 60, 0 |
| Age | 90 | 70 | 95 | 125 | 95 | 100 | 115 | 120 |
| Growing stock (m3 ha−1) | 400 | 280 | 240 | 280 | 220 | 230 | 180 | 210 |
| Site Characteristics | ||||||||
| Latitude (N) | 57°02′ | 58°25′ | 60°25′ | 60°35′ | 58°55′ | 60°20′ | 64°15′ | 62°45′ |
| Longitude (E) | 14°24′ | 16°35′ | 17°35′ | 12°35′ | 13°40′ | 16°15′ | 19°50′ | 15°45′ |
| Altitude (m a.s.l.) | 185 | 140 | 30 | 530 | 50 | 225 | 250 | 410 |
| Temperature sum b | 1324 | 1278 | 1250 | 821 | 1324 | 1092 | 834 | 791 |
| Site index (H100) c | 32 | 30 | 26 | 20 | 26 | 24 | 22 | 20 |
| Nitrogen deposition (kg N ha−1 yr−1) d | 10–12 | 8–10 | 2–4 | 6–8 | 4–6 | 4–6 | <2 | 2–4 |
| Soil texture | Sandy-loamy till | Sandy-loamy till | Loamy till | Sandy-loamy till | Sandy-loamy till | Sandy-loamy till | Sandy-loamy till | Sandy-loamy till |
| Stone content (% of soil volume) | 34–38 | 50–60 | 63–72 | 27–45 | 4–38 | 41–70 | 10–79 | 38–60 |
| Soil moisture | Mesic-moist | Mesic-moist | Mesic | Mesic | Mesic | Mesic | Mesic | Mesic |
| Treatments and Management | ||||||||
| Clearcutting of former stand | 1978 | 1980 | 1978 | 1979 | 1979 | 1978 | 1979 | 1979 |
| Stump and slash harvest | 1979 | 1980 | 1979 | 1980 | 1980 | 1979 | 1980 | 1980 |
| Site preparation and planting year | 1981 | 1981 | 1983 | 1983 | 1981 | 1982 | 1982 | 1983 |
| Age of planted seedlings, years | 5 | 5 | 2 | 1 | 3 | 2 | 1 | 2 |
| Planted species | Norway spruce | Norway spruce | Norway spruce | Norway spruce | Scots pine | Scots pine | Scots pine | Scots pine |
| Supplementary planting | 1982–1985 | 1983–1986 | 1984–1986 | 1986–1988 | 1982 | 1983–1985 | 1983–1985 | 1985–1986 |
| Pre-commercial thinning | 1986 | 1988, 1991, 1993, 1996 | 1987, 1991 | 2004 | 1986, 1992 | 1987, 1990, 1993 | 1987, 1991, 2004 | 1994 |
| Thinning | 2008, 14 | 2005 | ||||||
| Latest revision of inventory | 2014 | 2012 | 2013 | 2013 | 2012 | 2012 | 2013 | 2013 |
| Harvested Biomass (Mg ha−1) e | ||||||||
| Stumps (including roots >5 cm) | 58 | 36 | 30 | 40 | 26 | 28 | 24 | 26 |
| Slash f | 46 | 46 | 38 | 52 | 32 | 40 | 22 | 38 |
| Treatment | Spruce | Pine | All Sites | p-Value | |||
|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | ||
| Stem-only (control) | 158 | 56 | 180 | 19 | 172 | 23 | |
| Stump | 161 | 44 | 211 | 16 | 192 | 20 | |
| Slash | 134 | 53 | 184 | 16 | 165 | 23 | |
| Stump + Slash | 159 | 48 | 208 | 15 | 189 | 21 | |
| Post-hoc test | |||||||
| Stem-only vs. Stump | 0.06 | ||||||
| Stem-only vs. Slash | 0.68 | ||||||
| Stem-only vs. Stump + Slash | 0.06 | ||||||
| Stump vs. Slash | 0.02 | ||||||
| Stump vs. Stump + Slash | 0.99 | ||||||
| Slash vs. Stump + Slash | 0.36 | ||||||
| Treatment | Spruce | Pine | All Sites | p-Value | |||
|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | ||
| Stem-only (control) | 3.24 | 0,90 | 2.95 | 0.49 | 3.06 | 0.36 | |
| Stump | 3.35 | 0.65 | 3.12 | 0.49 | 3.22 | 0.28 | |
| Slash | 3.26 | 0.88 | 2.90 | 0.43 | 3.07 | 0.43 | |
| Stump + Slash | 3.77 | 1.06 | 2.90 | 0.48 | 3.17 | 0.52 | |
| Post-hoc test | |||||||
| Stem-only vs. Stump | 0.052 | ||||||
| Stem-only vs. Slash | 1.00 | ||||||
| Stem-only vs. Stump + Slash | 0.89 | ||||||
| Stump vs. Slash | 0.98 | ||||||
| Stump vs. Stump + Slash | 0.99 | ||||||
| Slash vs. Stump + Slash | 0.88 | ||||||
| Treatment | Spruce | Pine | All Sites | p-Value | |||
|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | ||
| Stem-only (control) | 49 | 11 | 79 | 3 | 64 | 7 | |
| Stump | 54 | 10 | 81 | 4 | 67 | 6 | |
| Slash | 53 | 9 | 85 | 3 | 69 | 6 | |
| Stump + Slash | 56 | 8 | 78 | 6 | 68 | 6 | |
| Post-hoc test | |||||||
| Stem-only vs. Stump | 0.39 | ||||||
| Stem-only vs. Slash | 0.06 | ||||||
| Stem-only vs. Stump + Slash | 0.38 | ||||||
| Stump vs. Slash | 0.95 | ||||||
| Stump vs. Stump + Slash | 1.00 | ||||||
| Slash vs. Stump + Slash | 0.97 | ||||||
| Treatment | Spruce | Pine | All sites | p-Value | |||
|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | ||
| Stem-only (control) | 27,135 | 8533 | 17,444 | 3496 | 22,290 | 4627 | |
| Stump | 21,608 | 5074 | 22,838 | 4166 | 22,223 | 3175 | |
| Slash | 32,664 | 5715 | 31,024 | 6680 | 31,844 | 4252 | |
| Stump + Slash | 26,653 | 4740 | 34,372 | 6056 | 30,513 | 3846 | |
| Post-hoc test | |||||||
| Stem-only vs. Stump | 0.99 | ||||||
| Stem-only vs. Slash | 0.051 | ||||||
| Stem-only vs. Stump + Slash | 0.10 | ||||||
| Stump vs. Slash | 0.09 | ||||||
| Stump vs. Stump + Slash | 0.19 | ||||||
| Slash vs. Stump + Slash | 0.98 | ||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurevics, A.; Peichl, M.; Egnell, G. Stand Volume Production in the Subsequent Stand during Three Decades Remains Unaffected by Slash and Stump Harvest in Nordic Forests. Forests 2018, 9, 770. https://doi.org/10.3390/f9120770
Jurevics A, Peichl M, Egnell G. Stand Volume Production in the Subsequent Stand during Three Decades Remains Unaffected by Slash and Stump Harvest in Nordic Forests. Forests. 2018; 9(12):770. https://doi.org/10.3390/f9120770
Chicago/Turabian StyleJurevics, Arnis, Matthias Peichl, and Gustaf Egnell. 2018. "Stand Volume Production in the Subsequent Stand during Three Decades Remains Unaffected by Slash and Stump Harvest in Nordic Forests" Forests 9, no. 12: 770. https://doi.org/10.3390/f9120770
APA StyleJurevics, A., Peichl, M., & Egnell, G. (2018). Stand Volume Production in the Subsequent Stand during Three Decades Remains Unaffected by Slash and Stump Harvest in Nordic Forests. Forests, 9(12), 770. https://doi.org/10.3390/f9120770