How to Calibrate Historical Aerial Photographs: A Change Analysis of Naturally Dynamic Boreal Forest Landscapes


Abstract
1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Aerial Photographs and Their Orientation
2.3. Visual Canopy Cover Interpretation
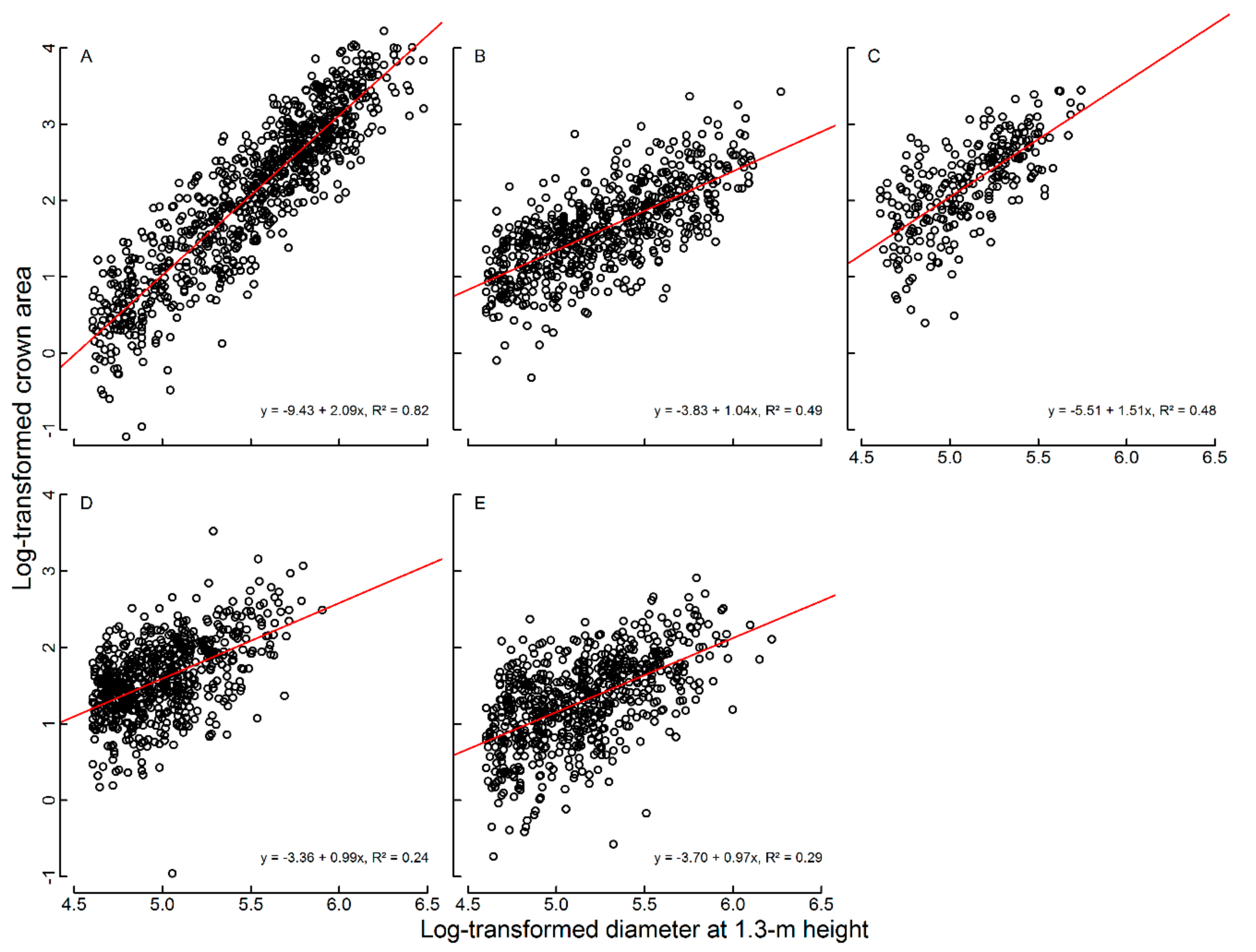
2.4. Field Data for the Calibration of the Canopy Cover Interpretation
2.5. Canopy Cover Reconstruction from the Field Data
2.6. Bias and Error in the Canopy Cover Interpretation
2.7. Canopy Cover Change Detection
3. Results
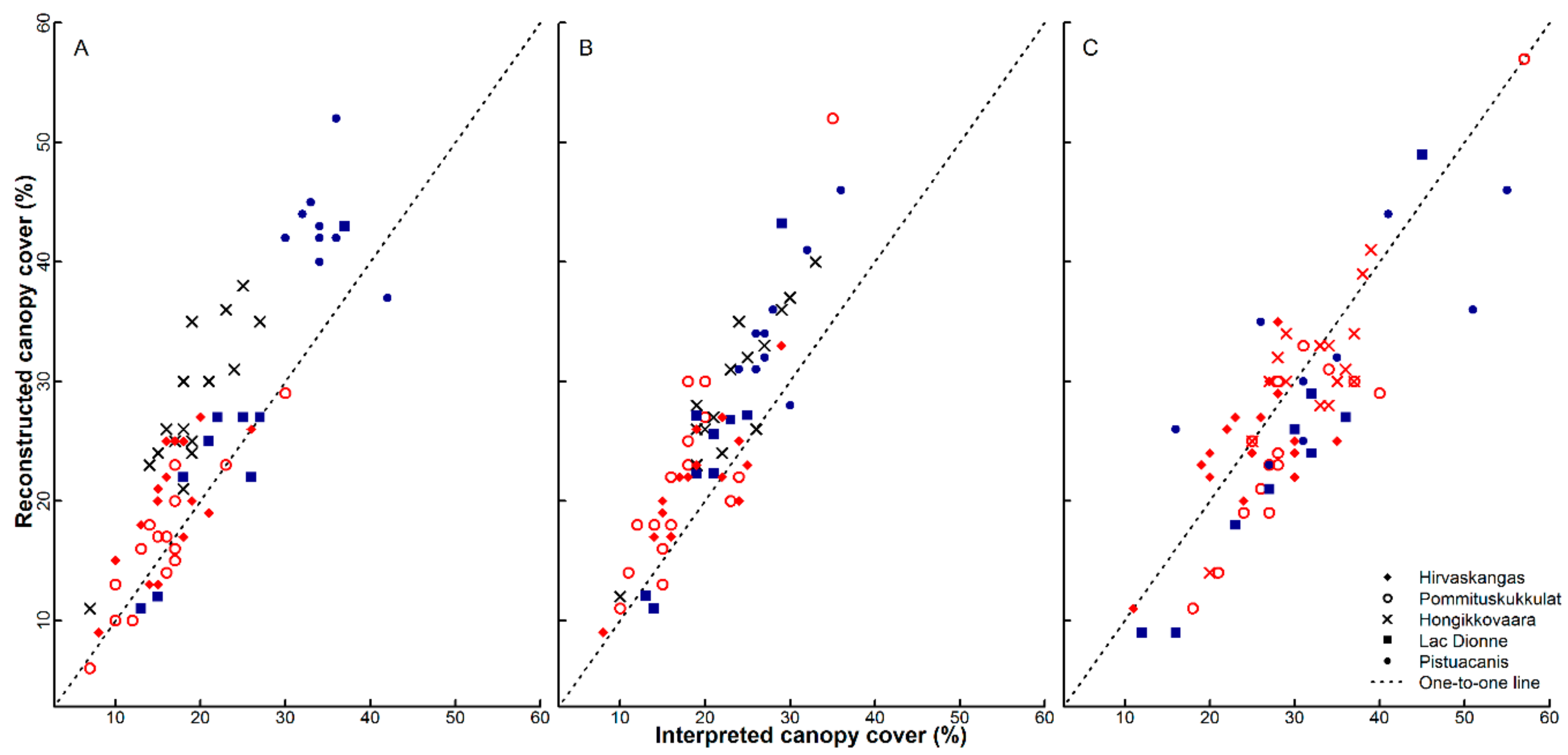
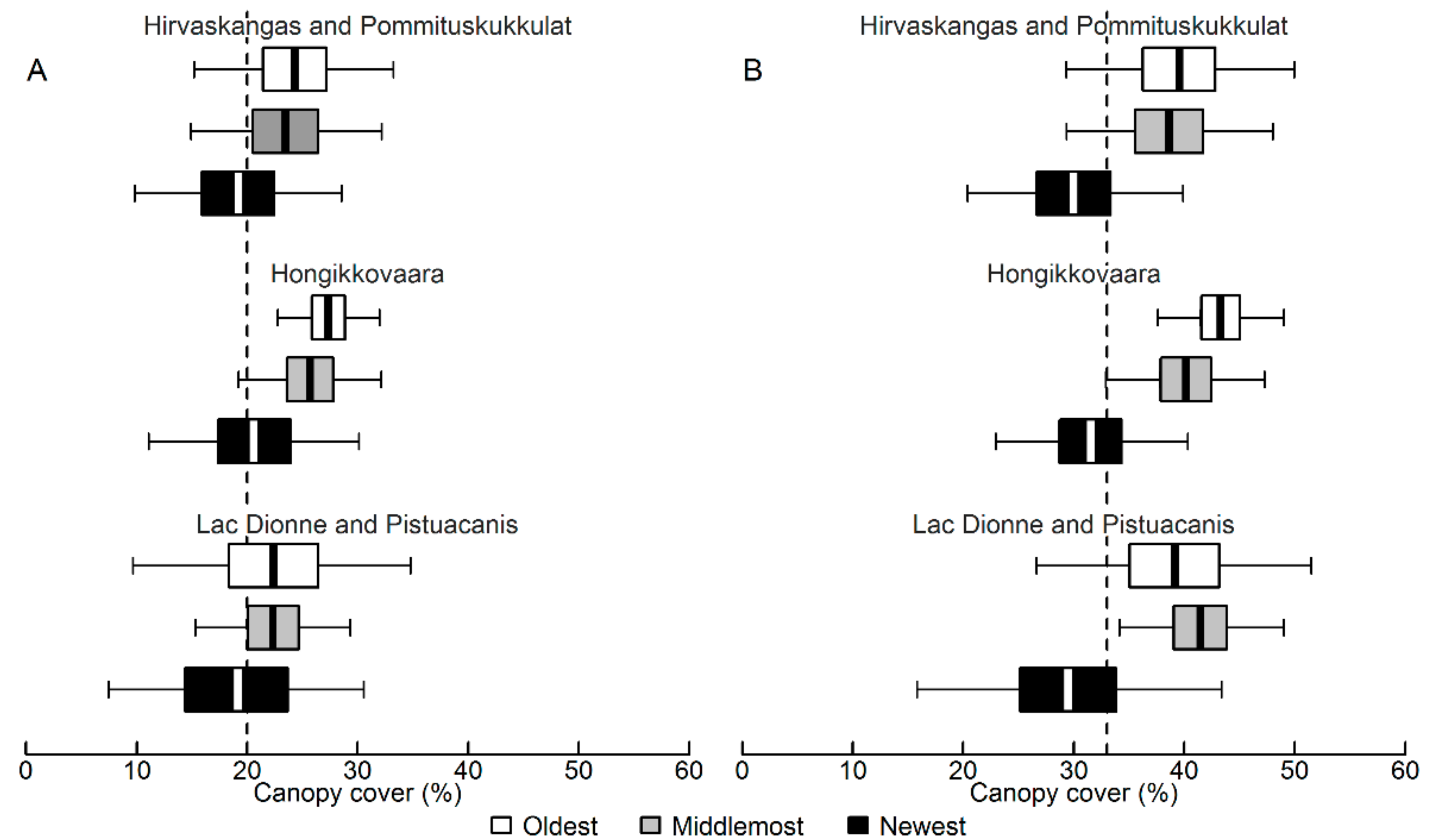
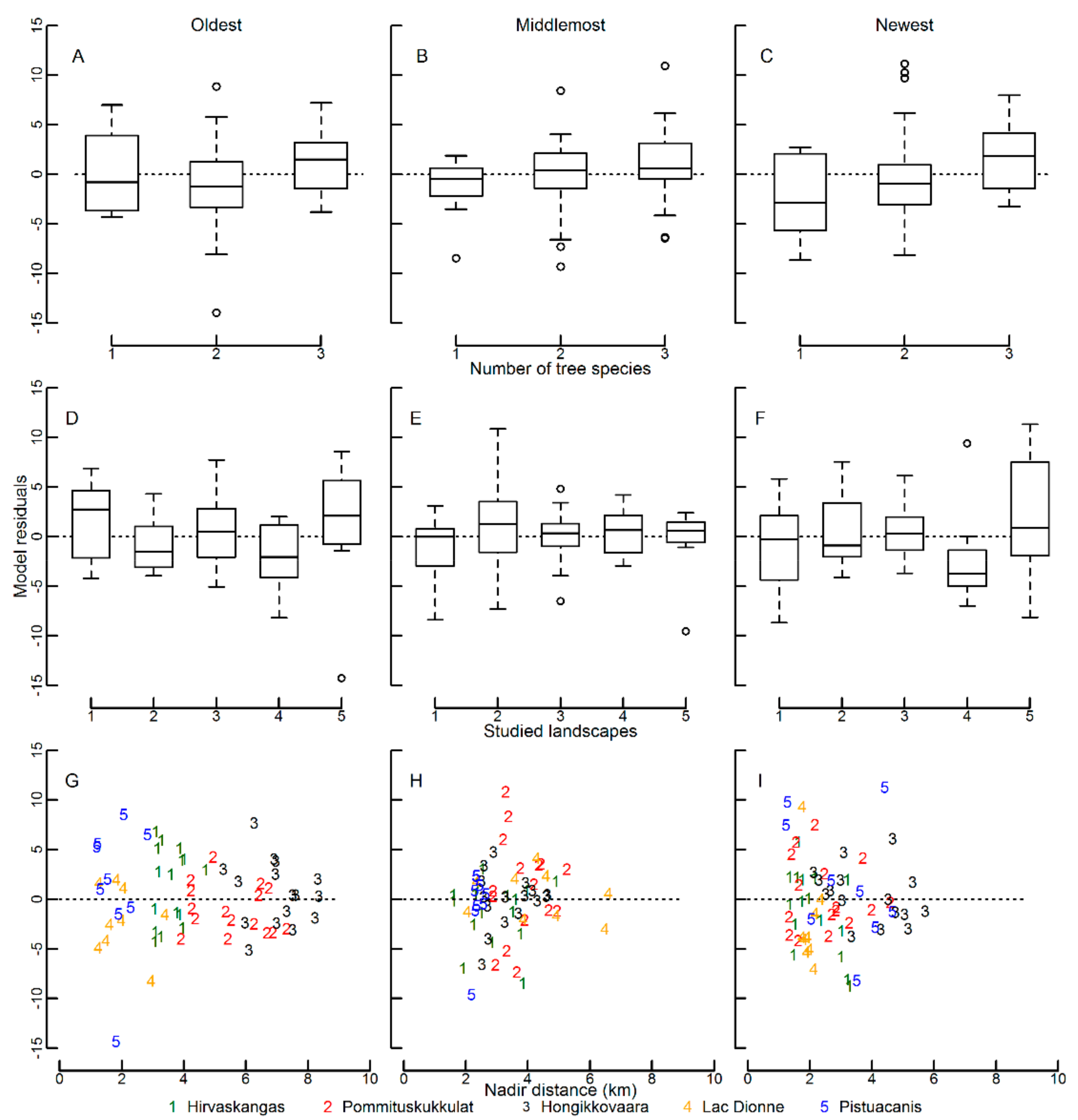
3.1. Bias and Error in the Canopy Cover Interpretation
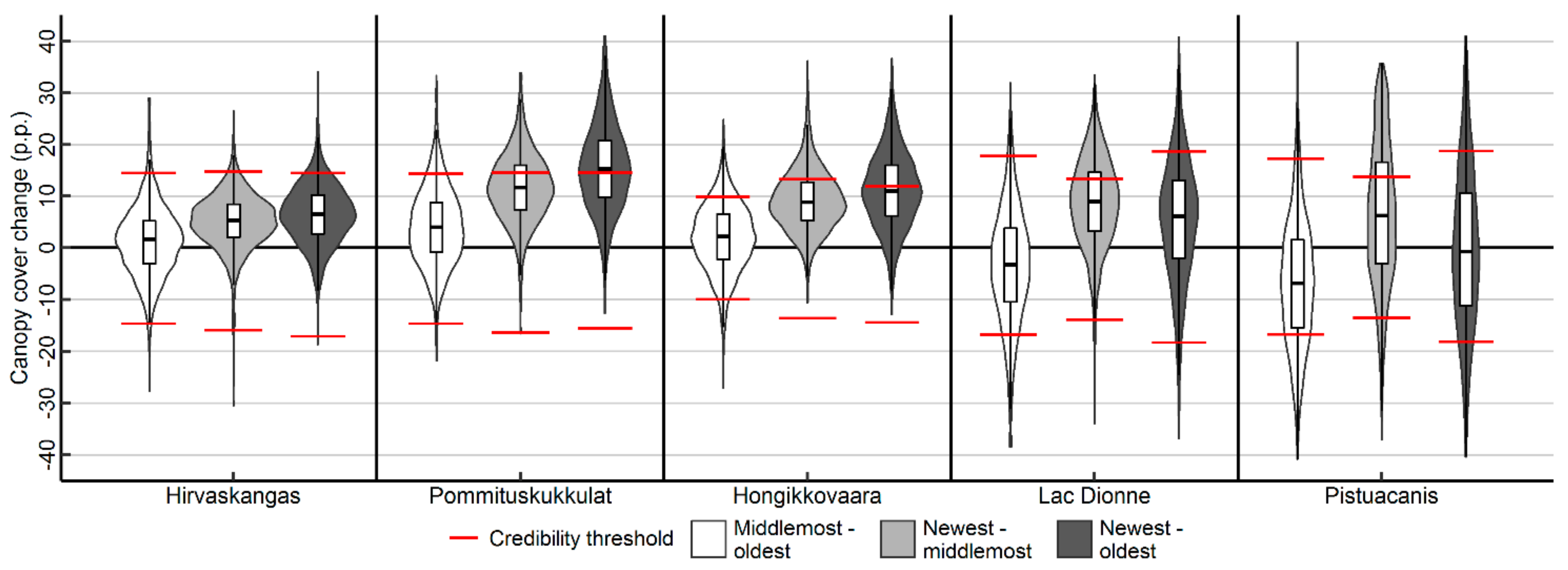
3.2. Canopy Cover Changes and Their Credibilities
4. Discussion
4.1. Bias and Error in the Visual Canopy Cover Interpretation
4.2. Canopy Cover Changes
4.3. Applicability in Other Studies
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictors | Degrees of Freedom | F-Test Score | Significance |
|---|---|---|---|
| Interpreted canopy cover | 1 | 758.54 | <0.00 |
| Aerial photo quality | 2 | 41.47 | <0.00 |
| Studied landscape | 4 | 18.89 | <0.00 |
| Interpreted canopy cover × Aerial photo quality | 2 | 6.05 | 0.00 |
| Interpreted canopy cover × Studied landscape | 4 | 6.98 | <0.00 |
| Aerial photo quality × Studied landscape | 8 | 4.59 | <0.00 |
| Predictors | R2 | RSE | AICc |
|---|---|---|---|
| I_CC | 0.68 | 4.41 | 283 |
| I_CC + P_abies | 0.78 | 3.66 | 265 |
| I_CC + P_sylvestris | 0.74 | 3.93 | 275 |
| I_CC + B_spp. | 0.68 | 4.43 | 285 |
- : canopy cover reconstructed from field measurements at plot in a given landscape or a group of landscapes at a given time point.
- : corresponding visually interpreted canopy cover
- : the sample size.
References
- Kuuluvainen, T.; Hofgaard, A.; Aakala, T.; Jonsson, B.-G. North Fennoscandian mountain forests: History, composition, disturbance dynamics and the unpredictable future. For. Ecol. Manag. 2017, 385, 140–149. [Google Scholar] [CrossRef]
- Pham, A.T.; de Grandpré, L.; Gauthier, S.; Bergeron, Y. Gap dynamics and replacement patterns in gaps of the northeastern boreal forest of Quebec. Can. J. For. Res. 2004, 34, 353–364. [Google Scholar] [CrossRef]
- Gauthier, S.; Boucher, D.; Morissette, J.; De Grandpré, L. Fifty-seven years of compositional change in the eastern Boreal forest of Canada. J. Veg. Sci. 2010, 21, 772–785. [Google Scholar] [CrossRef]
- Hofgaard, A. 50 years of change in Swedish boreal old-growth Picea abies forest. J. Veg. Sci. 1993, 4, 773–782. [Google Scholar] [CrossRef]
- Fraver, S.; Jonsson, B.G.; Jönsson, M.; Esseen, P.-A. Demographics and disturbance history of a boreal old-growth Picea abies forest. J. Veg. Sci. 2008, 19, 789–798. [Google Scholar] [CrossRef]
- Assal, T.J.; Sibold, J.; Reich, R. Modeling a historical mountain pine beetle outbreak using Landsat MSS and multiple lines of evidence. Remote Sens. Environ. 2014, 155, 275–288. [Google Scholar] [CrossRef]
- Fensham, R.J.; Bray, S.G.; Fairfax, R.J. Evaluation of aerial photography for predicting trends in structural attributes of Australian woodland including comparison with ground-based monitoring data. J. Environ. Manag. 2007, 83, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Nakashizuka, T.; Katsuki, T.; Tanaka, H. Forest canopy structure analyzed by using aerial photographs. Ecol. Res. 1995, 10, 13–18. [Google Scholar] [CrossRef]
- Fensham, R.J.; Fairfax, R.J. Effect of photoscale, interpreter bias and land type on woody crown-cover estimates from aerial photography. Aust. J. Bot. 2007, 55, 457–463. [Google Scholar] [CrossRef]
- Kulha, N.; Pasanen, L.; Holmström, L.; De Grandpré, L.; Kuuluvainen, T.; Aakala, T. At what scales and why does forest structure vary in naturally dynamic boreal forests? An analysis of forest landscapes at two continents. Ecosystems 2018. [Google Scholar] [CrossRef]
- Jackson, M.M.; Topp, E.; Gergel, S.E.; Martin, K.; Pirotti, F.; Sitzia, T. Expansion of subalpine woody vegetation over 40 years on Vancouver Island, British Columbia, Canada. Can. J. For. Res. 2016, 46, 437–443. [Google Scholar] [CrossRef]
- Lydersen, J.M.; Collins, B.M. Change in vegetation patterns over a large forested landscape based on historical and contemporary aerial photography. Ecosystems 2018. [Google Scholar] [CrossRef]
- Korpela, I. Individual Tree Measurements by Means of Digital Aerial Photogrammetry. Silva. Fenn. Monogr. 2004. Available online: https://helda.helsinki.fi/bitstream/handle/10138/20665/individu.pdf?sequence=2 (accessed on 15 July 2018).
- Lechner, A.M.; Langford, W.T.; Bekessy, S.A.; Jones, S.D. Are landscape ecologists addressing uncertainty in their remote sensing data? Landsc. Ecol. 2012, 27, 1249–1261. [Google Scholar] [CrossRef]
- Hughes, M.L.; McDowell, P.F.; Marcus, W.A. Accuracy assessment of georectified aerial photographs: Implications for measuring lateral channel movement in a GIS. Geomorphology 2006, 74, 1–16. [Google Scholar] [CrossRef]
- Danby, R.K.; Hik, D.S. Evidence of recent treeline dynamics in southwest Yukon from aerial photographs. Arctic 2007, 60, 411–420. [Google Scholar] [CrossRef]
- Fensham, R.J.; Fairfax, R.J.; Holman, J.E.; Whitehead, P.J. Quantitative assessment of vegetation structural attributes from aerial photography. Int. J. Remote Sens. 2002, 23, 2293–2317. [Google Scholar] [CrossRef]
- St-Onge, B.; Jumelet, J.; Cobello, M.; Véga, C. Measuring individual tree height using a combination of stereophotogrammetry and lidar. Can. J. For. Res. 2004, 34, 2122–2130. [Google Scholar] [CrossRef]
- Hudak, A.T.; Wessman, C.A. Textural Analysis of Historical Aerial Photography to Characterize Woody Plant Encroachment in South African Savanna. Remote Sens. Environ. 1998, 66, 317–330. [Google Scholar] [CrossRef]
- Morgan, J.L.; Gergel, S.E. Automated analysis of aerial photographs and potential for historic forest mapping. Can. J. For. Res. 2013, 43, 699–710. [Google Scholar] [CrossRef]
- D’Aoust, V.; Kneeshaw, D.; Bergeron, Y. Characterization of canopy openness before and after a spruce budworm outbreak in the southern boreal forest. Can. J. For. Res. 2004, 34, 339–352. [Google Scholar] [CrossRef]
- Browning, D.M.; Archer, S.R.; Byrne, A.T. Field validation of 1930s aerial photography: What are we missing? J. Arid Environ. 2009, 73, 844–853. [Google Scholar] [CrossRef]
- Massada, A.B.; Carmel, Y.; Tzur, G.E.; Grünzweig, J.M.; Yakir, D. Assessment of temporal changes in aboveground forest tree biomass using aerial photographs and allometric equations. Can. J. For. Res. 2006, 36, 2585–2594. [Google Scholar] [CrossRef]
- Teillet, P.M.; Guindon, B.; Goodenough, D.G. On the slope-aspect correction of multispectral scanner data. Can. J. Remote Sens. 1982, 8, 84–106. [Google Scholar] [CrossRef]
- Beaty, R.M.; Taylor, A.H. Spatial and temporal variation of fire regimes in a mixed conifer forest landscape, Southern Cascades, California, USA. J. Biogeogr. 2001, 28, 955–966. [Google Scholar] [CrossRef]
- Franco, J.A.; Morgan, J.W. Using historical records, aerial photography and dendroecological methods to determine vegetation changes in grassy woodland since European settlement. Aust. J. Bot. 2007, 55, 1–9. [Google Scholar] [CrossRef]
- Stephens, S.L.; Skinner, C.N.; Gill, S.J. Dendrochronology-based fire history of jeffery pine-mixed conifer forests in the Sierra San Pedro Matir, Mexico. Can. J. For. Res. 2003, 33, 1090–1101. [Google Scholar] [CrossRef]
- Bouchard, M.; Pothier, D. Spatiotemporal variability in tree and stand mortality caused by spruce budworm outbreaks in eastern Quebec. Can. J. For. Res. 2010, 40, 86–94. [Google Scholar] [CrossRef]
- Robitaille, A.; Saucier, J.-P. Paysages Régionaux du Québec Méridional; Les Publications du Québec: Sainte-Foy, QC, Canada, 1998; p. 214. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Aakala, T. Forest fire histories and tree age structures in Värriö and Maltio Strict Nature Reserves, northern Finland. Boreal Environ. Res. 2018, 23, 209–219. [Google Scholar]
- Hennigar, C.R.; MacLean, D.A.; Quiring, D.T.; Kershaw, J.A., Jr. Differences in spruce budworm defoliation among balsam fir and white, red, and black spruce. For. Sci. 2008, 54, 158–166. [Google Scholar] [CrossRef]
- Bognounou, F.; De Grandpré, L.; Pureswaran, D.S.; Kneeshaw, D. Temporal variation in plant neighborhood effects on the defoliation of primary and secondary hosts by an insect pest. Ecosphere 2017, 8, E01759. [Google Scholar] [CrossRef]
- Bouchard, M.; Pothier, D.; Gauthier, S. Fire return intervals and tree species succession in the North Shore region of eastern Quebec. Can. J. For. Res. 2008, 38, 1621–1633. [Google Scholar] [CrossRef]
- Korpela, I. Geometrically accurate time series of archived aerial images and airborne lidar data in a forest environment. Silva. Fenn. 2006, 40, 109–126. [Google Scholar] [CrossRef]
- Aakala, T.; Shimatani, K.; Abe, T.; Kubota, Y.; Kuuluvainen, T. Crown asymmetry in high latitude forests: Disentangling the directional effects of tree competition and solar radiation. Oikos 2016, 125, 1035–1043. [Google Scholar] [CrossRef]
- Aakala, T. Coarse woody debris in late-successional Picea abies forests in northern Europe: Variability in quantities and models of decay class dynamics. For. Ecol. Manag. 2010, 260, 770–779. [Google Scholar] [CrossRef]
- Aakala, T.; Berninger, F.; Starr, M. The roles of competition and climate in tree growth variation in northern boreal old-growth forests. J. Veg. Sci. 2018, in press. [Google Scholar]
- Krusic, P.J., Jr.; Hornbeck, J.W. Preserving decayed wood samples for tree-ring measurement. Tree-Ring Bull. 1989, 49, 23–27. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 51–67. [Google Scholar]
- Ilvessalo, Y. Metsänarvioiminen; WSOY: Porvoo, Finland, 1965; p. 400. (In Finnish) [Google Scholar]
- Li, R.; Weiskittel, A.R. Estimating and predicting bark thickness for seven conifer species in the Acadian region of North America using a mixed-effects modeling approach: A comparison of model forms and subsampling strategies. Eur. J. For. Res. 2011, 130, 219–233. [Google Scholar] [CrossRef]
- Ilvessalo, Y. On the correlation between crown diameter and the stem of the trees. Commun. Ins. For. Fenn. 1950, 38, 1–26. [Google Scholar]
- Bivand, R.; Rundel, C. Rgeos: Interface to Geometry Engine—Open Source (Geos), R Package Version 0.3-22; 2016. Available online: https://CRAN.R-project.org/package=rgeos (accessed on 13 March 2018).
- Yamaguchi, D.K. A simple method for cross-dating increment cores from living trees. Can. J. For. Res. 1991, 21, 414–416. [Google Scholar] [CrossRef]
- Kadmon, R.; Harari-Kremer, R.H. Studying long-term vegetation dynamics using digital processing of historical aerial photographs. Remote Sens. Environ. 1999, 68, 164–176. [Google Scholar] [CrossRef]
- Garbarino, M.; Lingua, E.; Subirà, M.M.; Motta, R. The larch wood pasture: Structure and dynamics of a cultural landscape. Eur. J. For. Res. 2011, 130, 491–502. [Google Scholar] [CrossRef]
- Zhou, W.Z.; Cadenasso, A.L. Effects of patch characteristics and within patch heterogeneity on the accuracy of urban land cover estimates from visual interpretation. Landsc. Ecol. 2012, 27, 1291–1305. [Google Scholar] [CrossRef]
- Caron, M.-N.; Kneeshaw, D.D.; De Grandpré, L.; Kauhanen, H.; Kuuluvainen, T. Canopy gap characteristics and disturbance dynamics in old-growth Picea abies stands in northern Fennoscandia: Is the forest in quasi-equilibrium? Ann. Bot. Fennici 2009, 46, 251–262. [Google Scholar] [CrossRef][Green Version]
- Weed, A.S.; Ayres, M.P.; Hicke, J.A. Consequences of climate change for biotic disturbances in North American forests. Ecol. Monogr. 2013, 83, 441–470. [Google Scholar] [CrossRef]
- Aakala, T. Temporal variability of deadwood volume and quality in boreal old-growth forests. Silva. Fenn. 2011, 45, 969–981. [Google Scholar] [CrossRef]
- Estes, L.; Elsen, P.R.; Treuer, T.; Ahmed, L.; Caylor, K.; Chang, J.; Choi, J.J.; Ellis, E.C. The spatial and temporal domains of modern ecology. Nat. Ecol. Evol. 2018, 2, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Song, D.-X.; Huang, C.; Sexton, J.O.; Channan, S.; Feng, M.; Townshend, J.R. Use of Landsat and Corona data for mapping forest cover change from the mid-1960s to 2000s: Case studies from the Eastern United States and Central Brazil. ISPRS J. Photogramm. Remote Sens. 2015, 103, 81–92. [Google Scholar] [CrossRef]
- Laliberte, A.S.; Rango, A.; Havstad, K.M.; Paris, J.F.; Beck, R.F.; McNeely, R.; Gonzalez, A.L. Object-oriented image analysis for mapping shrub encroachment from 1937 to 2003 in southern New Mexico. Remote Sens. Environ. 2004, 93, 198–210. [Google Scholar] [CrossRef]
- Næsset, E. Predicting forest stand characteristics with airborne scanning laser using a practical two-stage procedure and field data. Remote Sens. Environ. 2002, 80, 88–99. [Google Scholar] [CrossRef]
- Aakala, T.; Kuuluvainen, T.; Wallenius, T.; Kauhanen, H. Contrasting patterns of tree mortality in late-successional Picea abies stands in two areas in northern Fennoscandia. J. Veg. Sci. 2009, 20, 1016–1026. [Google Scholar] [CrossRef]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Rubin, D.B. Bayesian Data Analysis, 2nd ed.; Chapman & Hall/CRC: London, UK, 2004; p. 668. [Google Scholar]




| Oldest Photographs | Middlemost Photographs | Newest Photographs | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Year | Scale | Pixel Size (m) | Year | Scale | Pixel Size (m) | Year | Scale | Pixel Size (m) | |
| Hirvaskangas | 1959 | 1:30,000 | 0.6 | 1991 | 1:31,000 | 0.4 | 2011 | 1:20,000 | 0.5 |
| Pommituskukkulat | 1959 | 1:30,000 | 0.9 | 1988 | 1:30,000 | 0.4 | 2011 | 1:20,000 | 0.5 |
| Hongikkovaara | 1972 | 1:60,000 | 0.9 | 1988 | 1:30,000 | 0.4 | 2010 | 1:20,000 | 0.5 |
| Lac Dionne | 1965 | 1:16,000 | 0.2 | 1987 | 1:15,000 | 0.2 | 2011 | 1:11,000 | 0.3 |
| Pistuacanis | 1965 | 1:16,000 | 0.2 | 1987 | 1:15,000 | 0.2 | 2011 | 1:11,000 | 0.3 |
| Hirvaskangas and Pommituskukkulat | Hongikkovaara | Lac Dionne and Pistuacanis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | α | b | RSE | R2 | α | b | RSE | R2 | α | b | RSE | |
| Oldest | 0.61 | 3.14 | 0.93 | 3.51 | 0.75 | 4.19 | 1.24 | 3.41 | 0.79 | −3.84 | 1.31 | 5.70 |
| Middlemost | 0.68 | 0.92 | 1.15 | 4.34 | 0.83 | 2.87 | 1.13 | 2.76 | 0.89 | −7.28 | 1.49 | 3.13 |
| Newest | 0.78 | 5.20 | b1 0.84 b2 −7.59 | 3.66 | 0.78 | 5.20 | b1 0.84 b2 −7.59 | 3.66 | 0.70 | 2.74 | 0.81 | 6.14 |
| Hirvaskangas | Pommitus-kukkulat | Hongikkovaara | Lac Dionne | Pistuacanis | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Range | Mean | SD | Range | Mean | SD | Range | Mean | SD | Range | Mean | SD | Range | |
| Oldest photograph | 18 | 4 | 0–35 | 16 | 4 | 0–29 | 20 | 6 | 0–43 | 21 | 5 | 0–69 | 37 | 10 | 0–62 |
| Middlemost photograph | 20 | 4 | 0–43 | 20 | 5 | 0–40 | 22 | 5 | 0–41 | 21 | 7 | 0–51 | 30 | 9 | 0–59 |
| Newest photograph | 26 | 6 | 0–59 | 33 | 8 | 0–63 | 32 | 6 | 0–60 | 26 | 13 | 0–54 | 37 | 12 | 0–70 |
| Middlemost–oldest | 1 | 7 | −28–39 | 4 | 8 | −22–34 | 2 | 7 | −27–25 | −3 | 11 | −39–32 | −7 | 13 | −42–40 |
| Newest–middlemost | 5 | 5 | −31–27 | 12 | 7 | −17–34 | 9 | 6 | −11–36 | 9 | 8 | −34–33 | 7 | 13 | −37–36 |
| Newest–oldest | 7 | 6 | −18–34 | 16 | 9 | −13–41 | 11 | 7 | −13–37 | 6 | 12 | −37–41 | 0 | 15 | −40–41 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulha, N.; Pasanen, L.; Aakala, T. How to Calibrate Historical Aerial Photographs: A Change Analysis of Naturally Dynamic Boreal Forest Landscapes. Forests 2018, 9, 631. https://doi.org/10.3390/f9100631
Kulha N, Pasanen L, Aakala T. How to Calibrate Historical Aerial Photographs: A Change Analysis of Naturally Dynamic Boreal Forest Landscapes. Forests. 2018; 9(10):631. https://doi.org/10.3390/f9100631
Chicago/Turabian StyleKulha, Niko, Leena Pasanen, and Tuomas Aakala. 2018. "How to Calibrate Historical Aerial Photographs: A Change Analysis of Naturally Dynamic Boreal Forest Landscapes" Forests 9, no. 10: 631. https://doi.org/10.3390/f9100631
APA StyleKulha, N., Pasanen, L., & Aakala, T. (2018). How to Calibrate Historical Aerial Photographs: A Change Analysis of Naturally Dynamic Boreal Forest Landscapes. Forests, 9(10), 631. https://doi.org/10.3390/f9100631