Forty-Eight Years of Forest Succession: Tree Species Change across Four Forest Types in Mid-Missouri


Abstract
1. Introduction
2. Materials and Methods
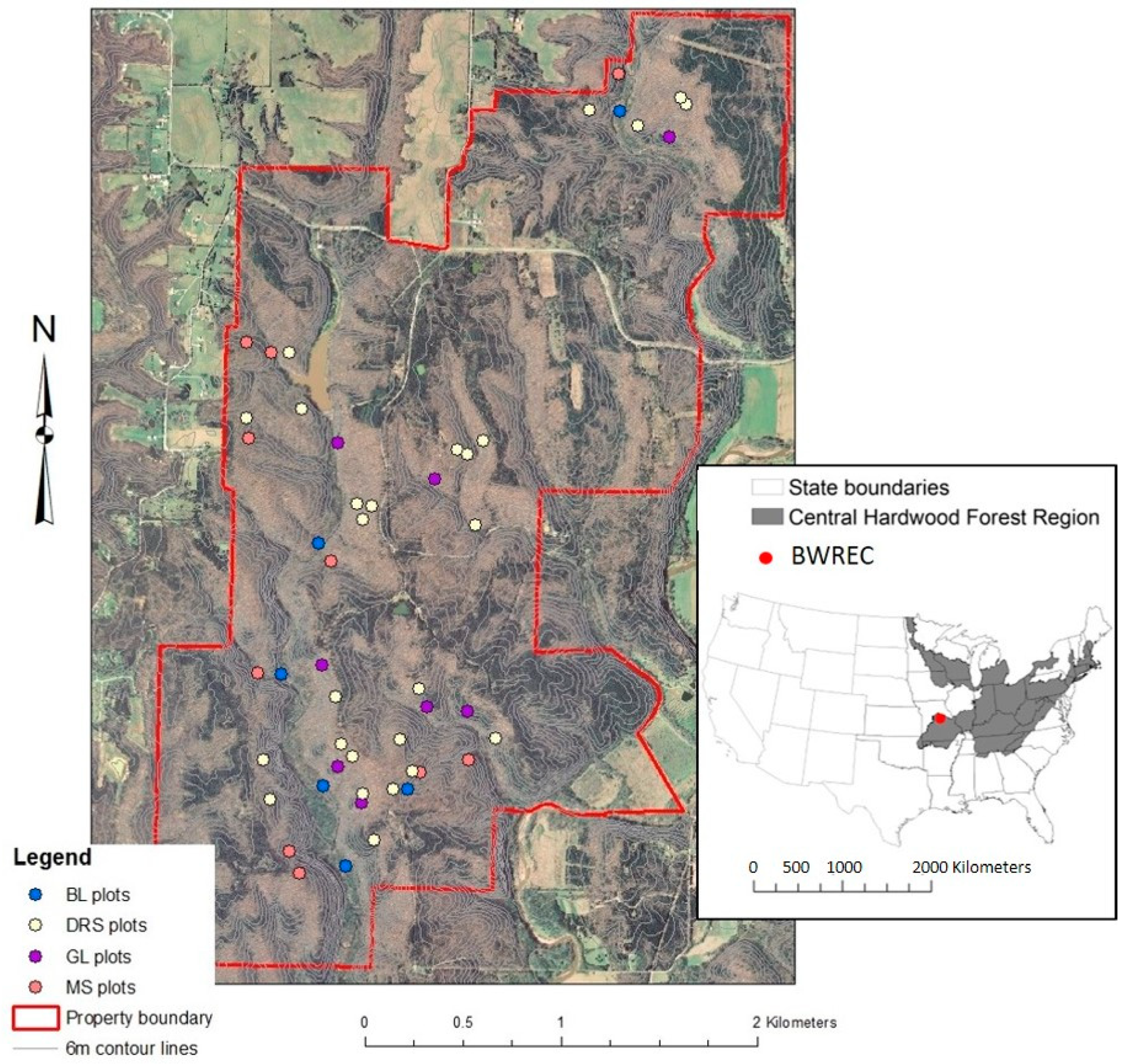
2.1. Study Site
2.2. Plot Establishment and Vegetation Sampling
2.3. Data Analyses
3. Results
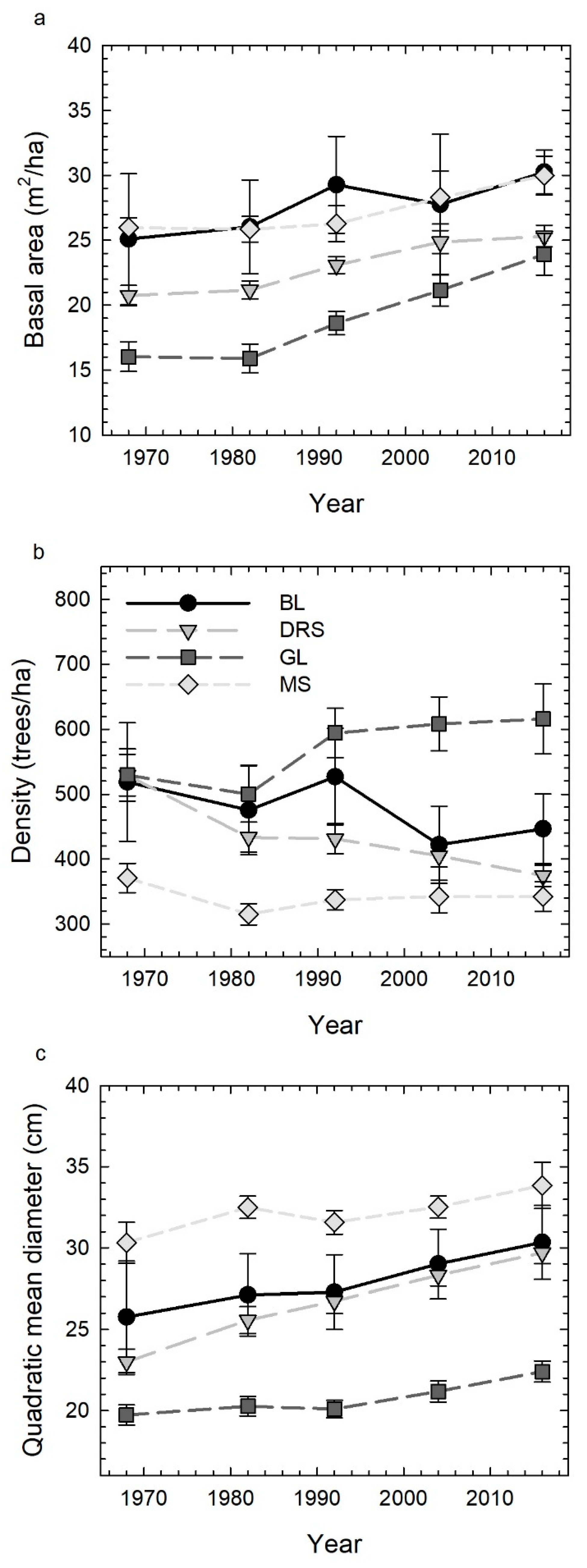
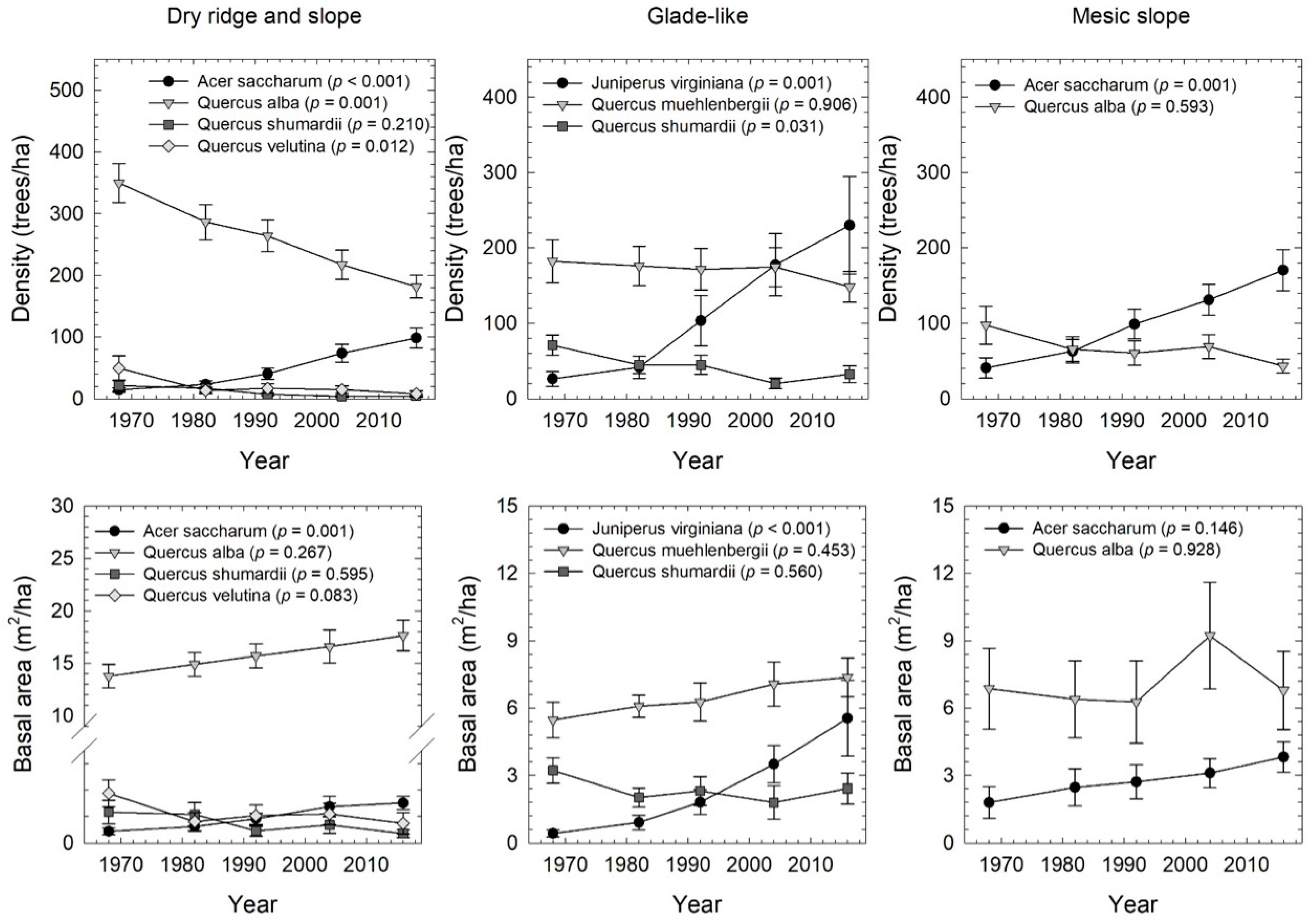
3.1. Forest Structure through Time
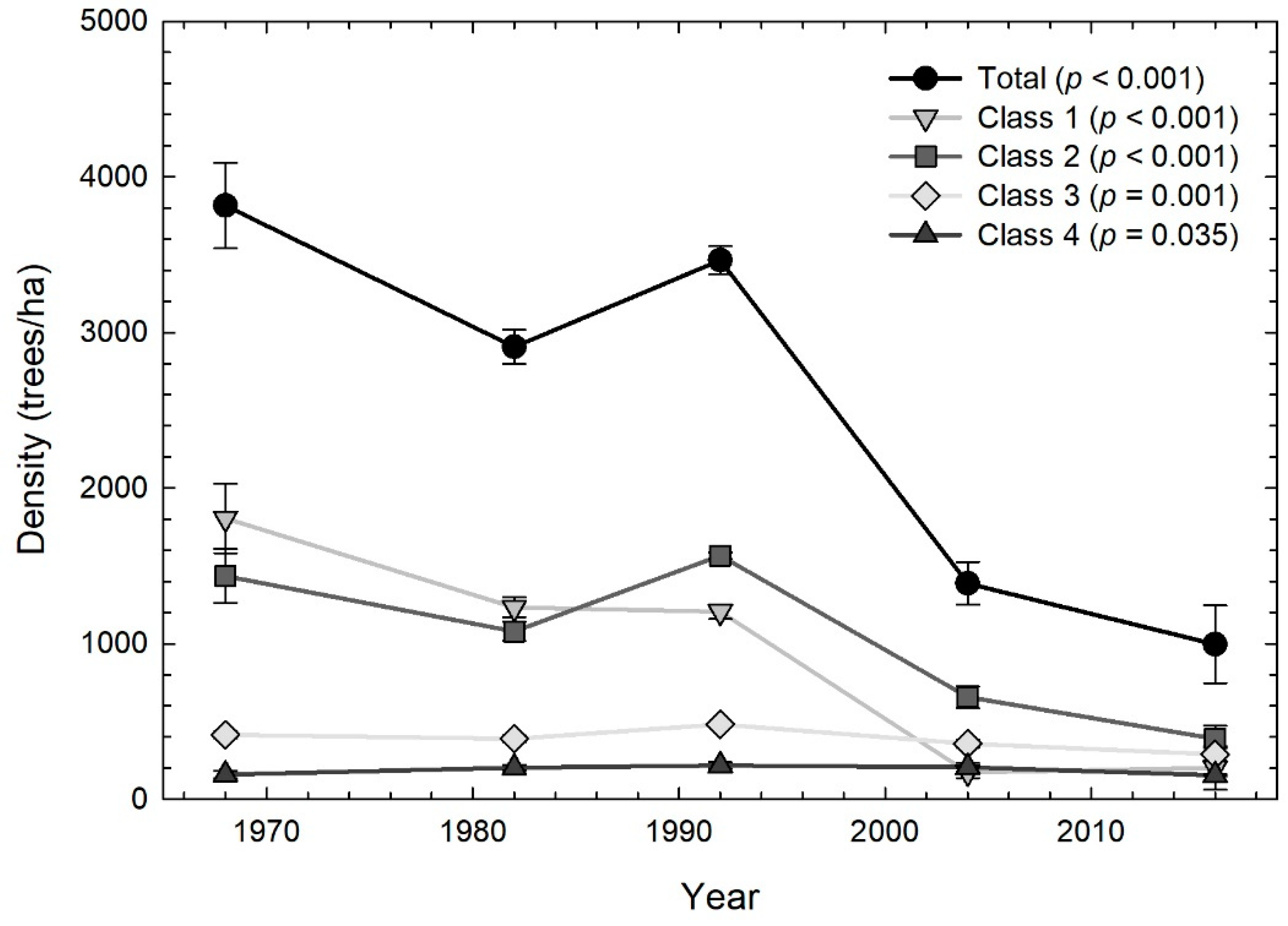
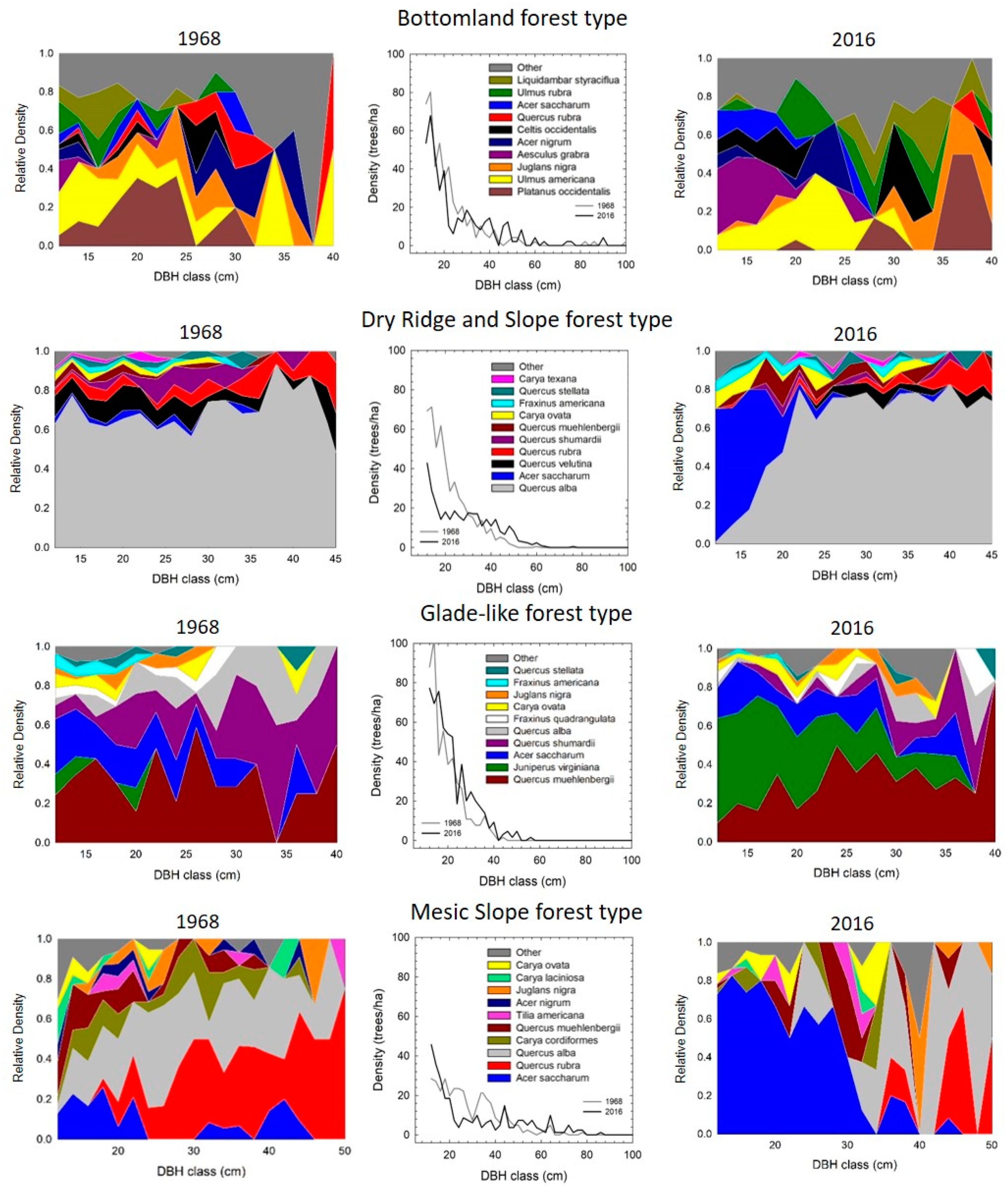
3.2. Forest Composition by Size Class
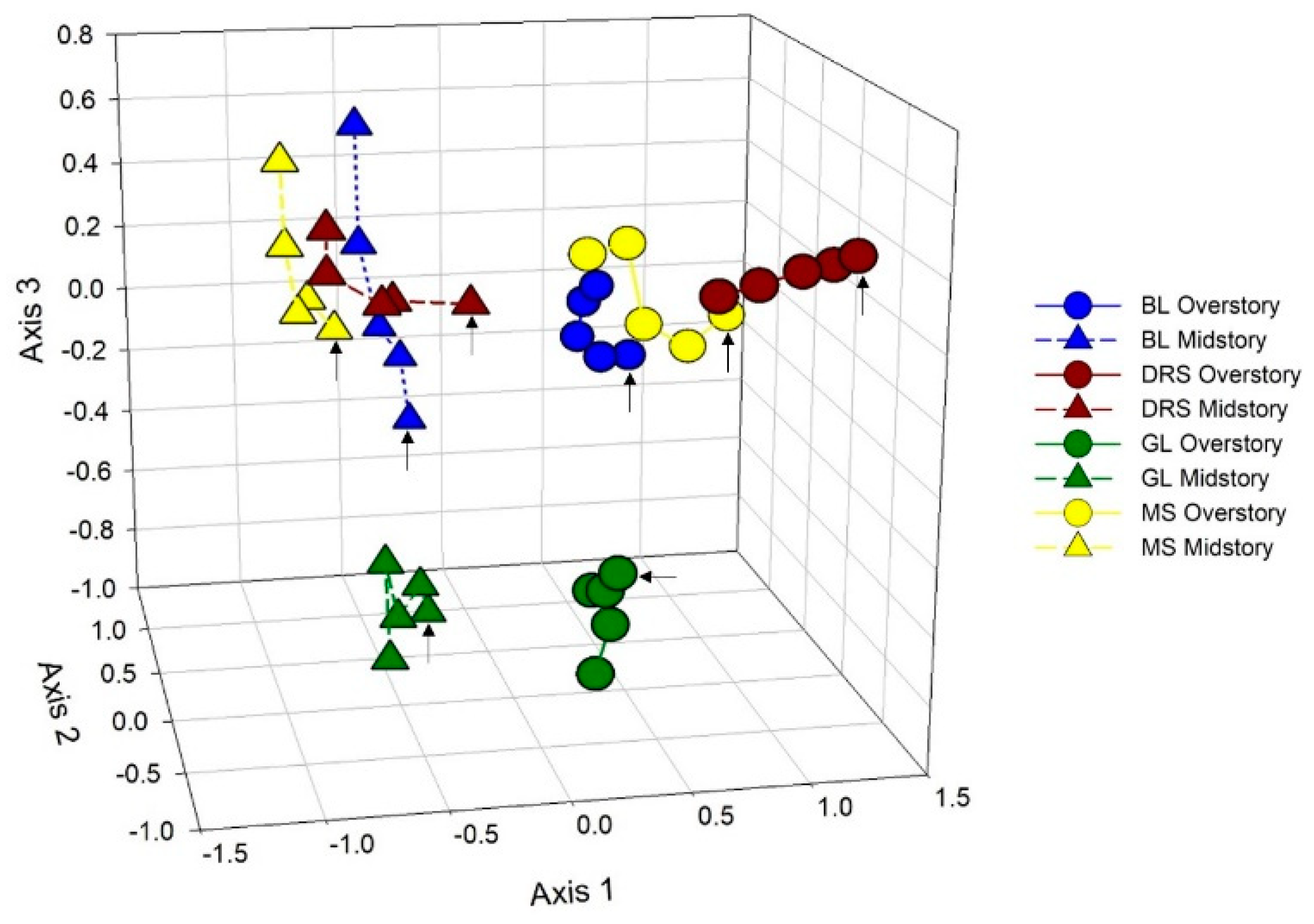
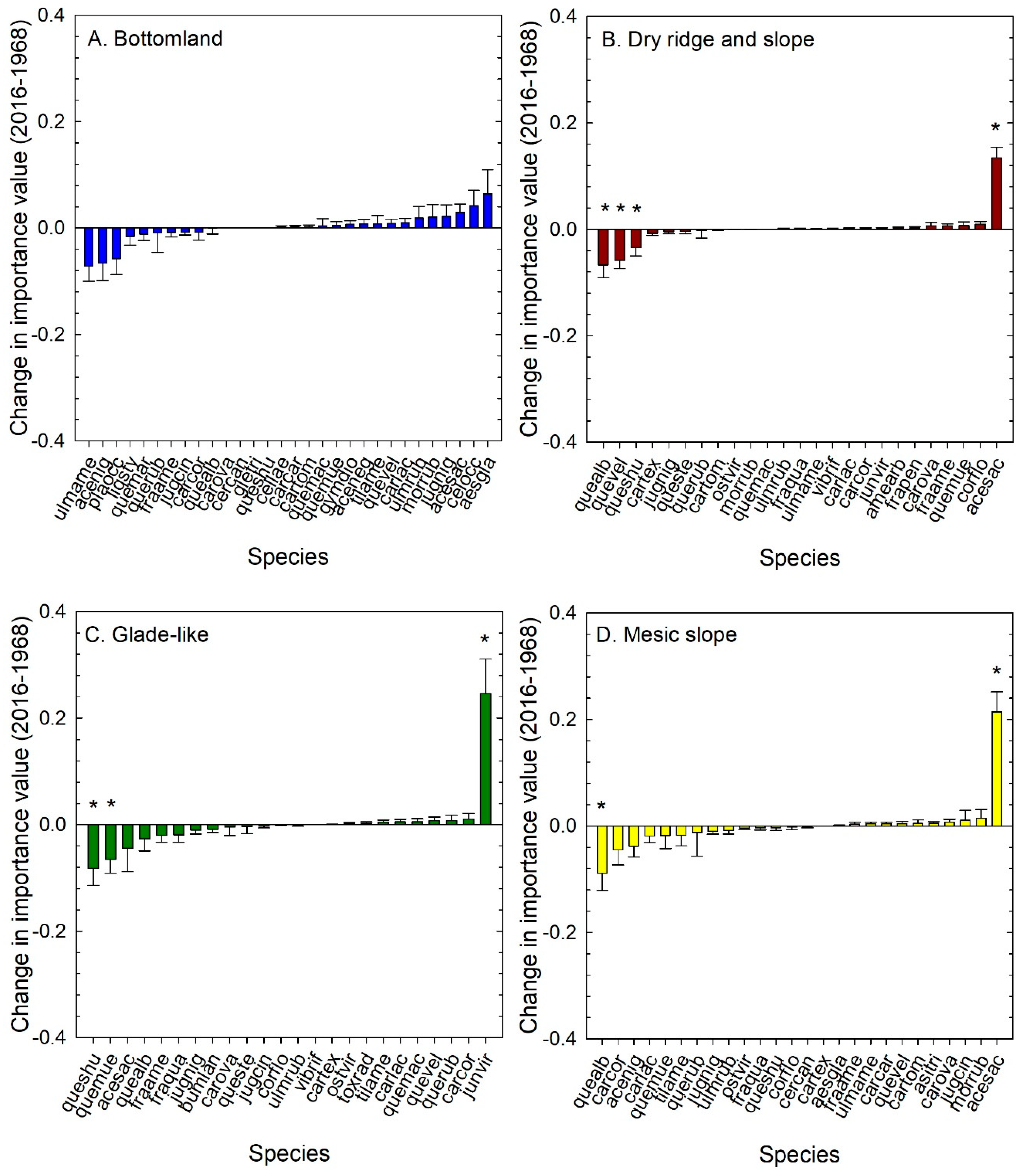
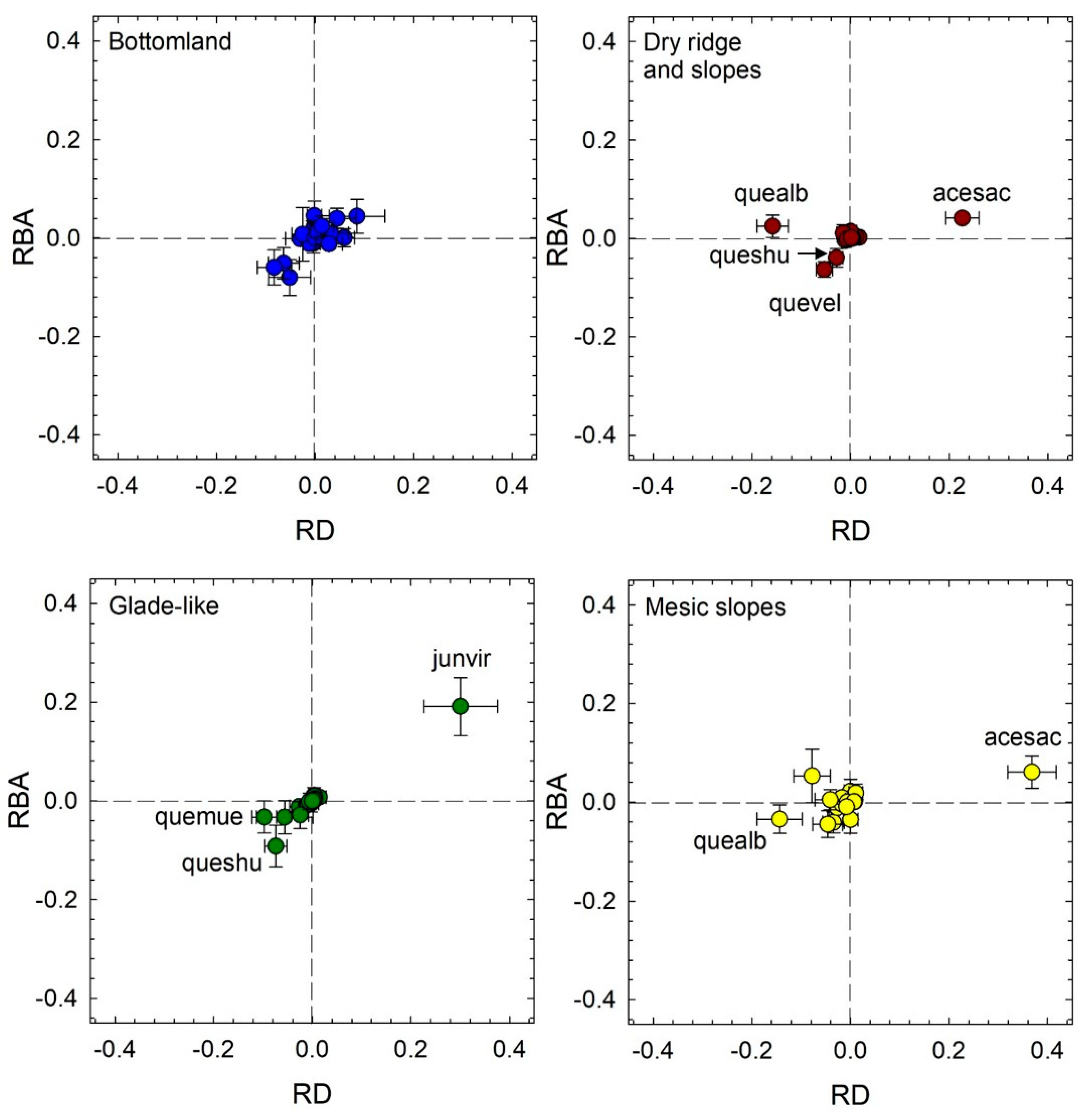
3.3. Tree Species Change through Time
4. Discussion
4.1. Patterns of Forest Change though Time
4.2. Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Finegan, B. Forest succession. Nature 1984, 312, 109. [Google Scholar] [CrossRef]
- Attiwill, P.M. The disturbance of forest ecosystems: The ecological basis for conservative management. For. Ecol. Manag. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Foster, D.; Swanson, F.; Aber, J.; Burke, I.; Brokaw, N.; Tilman, D.; Knapp, A. The importance of land-use legacies to ecology and conservation. BioScience 2003, 53, 77–88. [Google Scholar] [CrossRef]
- Dupouey, J.L.; Dambrine, E.; Laffite, J.D.; Moares, C. Irreversible impact of past land use on forest soils and biodiversity. Ecology 2002, 83, 2978–2984. [Google Scholar] [CrossRef]
- Christensen, N.L. An historical perspective on forest succession and its relevance to ecosystem restoration and conservation practice in North America. For. Ecol. Manag. 2014, 330, 312–322. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Braun, E.L. Deciduous Forests of Eastern North America; Hafner Publishing Company: New York, NY, USA, 1950; p. 596. [Google Scholar]
- Fralish, J.S. The Central Hardwood Forest: Its boundaries and physiographic provinces. In Proceedings of the 13th Central Hardwood Forest Conference, Urbana-Champaign, IL, USA, 1–3 April 2002; Van Sambeek, J.W., Dawson, J.O., Ponder, F., Jr., Loewenstein, E.F., Fralish, J.S., Eds.; General Technical Report NC-234; U.S. Department of Agriculture, Forest Service, North Central Research Station: St. Paul, MN, USA, 2003; pp. 1–20. [Google Scholar]
- Johnson, P.S.; Shifley, S.R.; Rogers, R. The Ecology and Silviculture of Oaks, 2nd ed.; CAB International: New York, NY, USA, 2009; p. 580. [Google Scholar]
- Hanberry, B.B.; Nowacki, G.J. Oaks were the historical foundation genus of the east-central United States. Quat. Sci. Rev. 2016, 145, 94–103. [Google Scholar] [CrossRef]
- Abrams, M.D. Fire and the development of oak forests. BioScience 1992, 42, 346–353. [Google Scholar] [CrossRef]
- McEwan, R.W.; Dyer, J.M.; Pederson, N. Multiple interacting ecosystem drivers: Toward an encompassing hypothesis of oak forest dynamics across eastern North America. Ecography 2011, 34, 244–256. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. The demise of fire and “mesophication” of forests in the eastern United States. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Fei, S.; Kong, N.; Steiner, K.C.; Moser, W.K.; Steiner, E.B. Change in oak abundance in the eastern United States from 1980 to 2008. For. Ecol. Manag. 2011, 262, 1370–1377. [Google Scholar] [CrossRef]
- Shifley, S.R.; Moser, W.K.; Nowak, D.J.; Miles, P.D.; Butler, B.J.; Aguilar, F.X.; DeSantis, R.D.; Greenfield, E.J. Five anthropogenic factors that will radically alter forest conditions and management needs in the northern United States. For. Sci. 2014, 60, 914–925. [Google Scholar] [CrossRef]
- Kabrick, J.M.; Zenner, E.K.; Dey, D.C.; Gwaze, D.; Jensen, R.G. Using ecological land types to examine landscape-scale oak regeneration dynamics. For. Ecol. Manag. 2008, 255, 3051–3062. [Google Scholar] [CrossRef]
- Kabrick, J.M.; Villwock, J.L.; Dey, D.C.; Keyser, T.L.; Larsen, D.R. Modeling and mapping oak advance reproduction density using soil and site variables. For. Sci. 2014, 60, 1107–1117. [Google Scholar] [CrossRef]
- Olson, M.G.; Knapp, B.O.; Kabrick, J.M. Dynamics of a temperate deciduous forest under landscape-scale management: Implications for adaptability to climate change. For. Ecol. Manag. 2017, 387, 73–85. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Dey, D.C.; He, H.S. Regime shifts and weakened environmental gradients in open oak and pine ecosystems. PLoS ONE 2012, 7, e41337. [Google Scholar] [CrossRef] [PubMed]
- Hanberry, B.B.; Kabrick, J.M.; He, H.S. Densification and state transition across the Missouri Ozarks landscape. Ecosystems 2014, 17, 66–81. [Google Scholar] [CrossRef]
- Guyette, R.P.; Muzika, R.M.; Dey, D.C. Dynamics of an anthropogenic fire regime. Ecosystems 2002, 5, 472–486. [Google Scholar]
- Batek, M.J.; Rebertus, A.J.; Schroeder, W.A.; Haithcoat, T.L.; Compas, E.; Guyette, R.P. Reconstruction of early nineteenth-century vegetation and fire regimes in the Missouri Ozarks. J. Biogeogr. 1999, 26, 397–412. [Google Scholar] [CrossRef]
- Nelson, P.W. Fire-adapted communities natural communities of the ozark highlands at the time of european settlement and now. In Proceedings of the 4th Fire in Eastern Oak Forests Conference, Springfield, MO, USA, 17–19 May 2011; Dey, D.C., Stambaugh, M.C., Clark, S.L., Schweitzer, C.J., Eds.; General Technical Report NRS-P-102; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012; pp. 92–102. [Google Scholar]
- Ware, S. Rock outcrop plant communities (glades) in the Ozarks: A synthesis. Southwest. Nat. 2002, 47, 585–597. [Google Scholar] [CrossRef]
- Rochow, J.J. A vegetational description of a mid-Missouri forest using gradient analysis techniques. Am. Midl. Nat. 1972, 87, 377–396. [Google Scholar] [CrossRef]
- Rochow, J.J. Mineral nutrient pool and cycling in a Missouri forest. J. Ecol. 1975, 63, 985–994. [Google Scholar] [CrossRef]
- Rochow, J.J. Litter fall relations in a Missouri forest. Oikos 1974, 25, 80–85. [Google Scholar] [CrossRef]
- Belden, A.C.; Pallardy, S.G. Successional trends and apparent Acer saccharum regeneration failure in an oak-hickory forest in central Missouri, USA. Plant Ecol. 2009, 204, 305–322. [Google Scholar] [CrossRef]
- Pallardy, S.G.; Nigh, T.A.; Garrett, H.E. Changes in forest composition in central Missouri: 1968–1982. Am. Midl. Nat. 1988, 120, 380–390. [Google Scholar] [CrossRef]
- Nigh, T.A.; Schroeder, W.A. Atlas of Missouri Ecoregions; Missouri Department of Conservation: Jefferson City, MO, USA, 2002; p. 212. [Google Scholar]
- Critchfield, H.J. General Climatology; Prentice-Hall: Englewood Cliffs, NJ, USA, 1966; p. 446. [Google Scholar]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Peet, R.K.; Christensen, N.L. Competition and tree death. BioScience 1987, 37, 586–595. [Google Scholar] [CrossRef]
- Oliver, C.D. Forest development in North America following major disturbances. For. Ecol. Manag. 1980, 3, 153–168. [Google Scholar] [CrossRef]
- Kabrick, J.M.; Dey, D.C.; Jensen, R.G.; Wallendorf, M. The role of environmental factors in oak decline and mortality in the Ozark Highlands. For. Ecol. Manag. 2008, 255, 1409–1417. [Google Scholar] [CrossRef]
- Shifley, S.R.; Fan, Z.; Kabrick, J.M.; Jensen, R.G. Oak mortality risk factors and mortality estimation. For. Ecol. Manag. 2006, 229, 16–26. [Google Scholar] [CrossRef]
- Fan, Z.; Fan, X.; Crosby, M.K.; Moser, W.K.; He, H.; Spetich, M.A.; Shifley, S.R. Spatio-temporal trends of oak decline and mortality under periodic regional drought in the Ozark Highlands of Arkansas and Missouri. Forests 2012, 3, 614–631. [Google Scholar] [CrossRef]
- Voelker, S.L.; Muzika, R.-M.; Guyette, R.P. Individual tree and stand level influences on the growth, vigor, and decline of red oaks in the Ozarks. For. Sci. 2008, 54, 8–20. [Google Scholar]
- Jenkins, M.A.; Pallardy, S.G. The influence of drought on red oak group species growth and mortality in the Missouri Ozarks. Can. J. For. Res. 1995, 25, 1119–1127. [Google Scholar] [CrossRef]
- Wood, J.D.; Knapp, B.O.; Muzika, R.M.; Stambaugh, M.C.; Gu, L. The importance of drought–pathogen interactions in driving oak mortality events in the Ozark Border region. Environ. Res. Lett. 2018, 13, 015004. [Google Scholar] [CrossRef]
- Nigh, T.A.; Pallardy, S.G.; Garrett, H.E. Sugar maple-environment relationships in the River Hills and central Ozark Mountains of Missouri. Am. Midl. Nat. 1985, 114, 235–251. [Google Scholar] [CrossRef]
- Stevens, J.H. Site Quality Factors Affecting Acer saccharum, Quercus rubra, and Quercus alba Abundance and Height Growth Rates in Young Even-Aged Stands Located in Central Missouri. Master’s Thesis, University of Missouri, Columbia, MO, USA, 2013. [Google Scholar]
- Howell, D.L.; Kucera, C.L. Composition of pre-settlement forests in three counties of Missouri. Bull. Torrey Bot. Club 1956, 83, 207–217. [Google Scholar] [CrossRef]
- Wenk, E.S.; Callaham, M.A.; O’Brien, J.J.; Hanson, P.J. Soil macroinvertebrate communities across a productivity gradient in deciduous forests of eastern North America. Northeast. Nat. 2016, 23, 25–44. [Google Scholar] [CrossRef]
- McFarlane, K.J.; Torn, M.S.; Hanson, P.J.; Porras, R.C.; Swanston, C.W.; Callaham, M.A.; Guilderson, T.P. Comparison of soil organic matter dynamics at five temperate deciduous forests with physical fractionation and radiocarbon measurements. Biogeochemistry 2013, 112, 457–476. [Google Scholar] [CrossRef]
- Groffman, P.M.; Bohlen, P.J.; Fisk, M.C.; Fahey, T.J. Exotic earthworm invasion and microbial biomass in temperate forest soils. Ecosystems 2004, 7, 45–54. [Google Scholar] [CrossRef]
- Burtelow, A.E.; Bohlen, P.J.; Groffman, P.M. Influence of exotic earthworm invasion on soil organic matter, microbial biomass and denitrification potential in forest soils of the northeastern United States. Appl. Soil Ecol. 1998, 9, 197–202. [Google Scholar] [CrossRef]
- Suárez, E.R.; Fahey, T.J.; Yavitt, J.B.; Groffman, P.M.; Bohlen, P.J. Patterns of litter disappearance in a northern hardwood forest invaded by exotic earthworms. Ecol. Appl. 2006, 16, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Hale, C.M.; Frelich, L.E.; Reich, P.B. Changes in hardwood forest understory plant communities in response to European earthworm invasions. Ecology 2006, 87, 1637–1649. [Google Scholar] [CrossRef]
- Bohlen, P.J.; Scheu, S.; Hale, C.M.; McLean, M.A.; Migge, S.; Groffman, P.M.; Parkinson, D. Non-native invasive earthworms as agents of change in northern temperate forests. Front. Ecol. Environ. 2004, 2, 427–435. [Google Scholar] [CrossRef]
- Corio, K.; Wolf, A.; Draney, M.; Fewless, G. Exotic earthworms of great lakes forests: A search for indicator plant species in maple forests. For. Ecol. Manag. 2009, 258, 1059–1066. [Google Scholar] [CrossRef]
- Holdsworth, A.R.; Frelich, L.E.; Reich, P.B. Effects of earthworm invasion on plant species richness in northern hardwood forests. Conserv. Biol. 2007, 21, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.R.; Dickinson, Y.L.; Burton, J.I.; Frelich, L.E.; Jenkins, M.A.; Kern, C.C.; Raymond, P.; Saunders, M.R.; Walters, M.B.; Willis, J.L. Promoting and maintaining diversity in contemporary hardwood forests: Confronting contemporary drivers of change and the loss of ecological memory. For. Ecol. Manag. 2018, 421, 98–108. [Google Scholar] [CrossRef]
- Nelson, P.W. The Terrestrial Natural Communities of Missouri; Missouri Natural Areas Committee: Jefferson City, MO, USA, 2005; p. 550. [Google Scholar]
- Kucera, C.L.; Martin, S.C. Vegetation and soil relationships in the glade region of the southwestern Missouri Ozarks. Ecology 1957, 38, 285–291. [Google Scholar] [CrossRef]
- Van Zandt, P.A.; Collins, E.; Losos, J.B.; Chase, J.M. Implications of food web interactions for restoration of Missouri Ozark glade habitats. Restor. Ecol. 2005, 13, 312–317. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Vegetation of limestone and dolomite glades in the Ozarks and Midwest regions of the United States. Ann. Mo. Bot. Gard. 2000, 87, 286–294. [Google Scholar] [CrossRef]
- Kolb, T.E.; McCormick, L.H. Etiology of sugar maple decline in four Pennsylvania stands. Can. J. For. Res. 1993, 23, 2395–2402. [Google Scholar] [CrossRef]
- Horsley, S.B.; Long, R.P.; Bailey, S.W.; Hallett, R.A.; Wargo, P.M. Health of eastern North American sugar maple forests and factors affecting decline. North. J. Appl. For. 2002, 19, 34–44. [Google Scholar]
- Putnam, R.C.; Reich, P.B. Climate and competition affect growth and survival of transplanted sugar maple seedlings along a 1700-km gradient. Ecol. Monogr. 2017, 87, 130–157. [Google Scholar] [CrossRef]
- Gu, L.; Pallardy, S.G.; Hosman, K.P.; Sun, Y. Drought-influenced mortality of tree species with different predawn leaf water dynamics in a decade-long study of a central US forest. Biogeosciences 2015, 12, 2831–2845. [Google Scholar] [CrossRef]
- Larsen, D.R.; Johnson, P.S. Linking the ecology of natural oak regeneration to silviculture. For. Ecol. Manag. 1998, 106, 1–7. [Google Scholar] [CrossRef]
- Morrissey, R.C.; Jacobs, D.F.; Seifert, J.R.; Fischer, B.C.; Kershaw, J.A. Competitive success of natural oak regeneration in clearcuts during the stem exclusion stage. Can. J. For. Res. 2008, 38, 1419–1430. [Google Scholar] [CrossRef]
- Dey, D.C.; Johnson, P.S.; Garrett, H.E. Modeling the regeneration of oak stands in the Missouri Ozark Highlands. Can. J. For. Res. 1996, 26, 573–583. [Google Scholar] [CrossRef]
- Sander, I.L.; Johnson, P.S.; Rogers, R. Evaluating Oak Advance Reproduction in the Missouri Ozarks; General Technical Report NC-251; U.S. Department of Agriculture, Forest Service, North Central Experiment Station: St. Paul, MN, USA, 1984; p. 16.
- Larsen, D.R.; Metzger, M.A.; Johnson, P.S. Oak regeneration and overstory density in the Missouri Ozarks. Can. J. For. Res. 1997, 27, 869–875. [Google Scholar] [CrossRef]
- Loftis, D.L. A shelterwood method for regenerating red oak in the Southern Appalachians. For. Sci. 1990, 36, 917–929. [Google Scholar]
- Iverson, L.R.; Peters, M.P.; Bartig, J.L.; Rebbeck, J.; Hutchinson, T.F.; Matthews, S.N.; Stout, S. Spatial modeling and inventories for prioritizing investment into oak-hickory restoration. For. Ecol. Manag. 2018, 424, 355–366. [Google Scholar] [CrossRef]
- Fei, S.; Steiner, K.C. Relationships between advance oak regeneration and biotic and abiotic factors. Tree Physiol. 2008, 28, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Loewenstein, E.F.; Johnson, P.S.; Garrett, H.E. Age and diameter structure of a managed uneven-aged oak forest. Can. J. For. Res. 2000, 30, 1060–1070. [Google Scholar] [CrossRef]
- Brose, P.H.; Van Lear, D.H.; Keyser, P.D. A shelterwood—Burn technique for regenerating productive upland oak sites in the Piedmont region. South. J. Appl. For. 1999, 23, 158–163. [Google Scholar]
- Arthur, M.A.; Alexander, H.D.; Dey, D.C.; Schweitzer, C.J.; Loftis, D.L. Refining the oak-fire hypothesis for management of oak-dominated forests of the eastern United States. J. For. 2012, 110, 257–266. [Google Scholar] [CrossRef]
- Brose, P.H.; Dey, D.C.; Phillips, R.J.; Waldrop, T.A. A meta-analysis of the fire-oak hypothesis: Does prescribed burning promote oak reproduction in eastern North America? For. Sci. 2013, 59, 322–334. [Google Scholar] [CrossRef]
- Brose, P.H.; Gottschalk, K.W.; Horsley, S.B.; Knopp, P.D.; Kochenderfer, J.N.; McGuinness, B.J.; Miller, G.W.; Ristau, T.E.; Stoleson, S.H.; Stout, S.L. Prescribing Regeneration Treatments for Mixed-Oak Forests in the Mid-Atlantic Region; General Technical Report NRS-33; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2008; p. 100.
- Kochenderfer, J.D.; Kochenderfer, J.N.; Miller, G.W. Manual Herbicide Applicaiton Methods for Managing Vegetation in Appalachian Hardwood Forests; General Technical Report NRS-96; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012; p. 59.
- Lhotka, J.M.; Loewenstein, E.F. Effect of midstory removal on understory light availability and the 2-year response of underplanted cherrybark oak seedlings. South. J. Appl. For. 2009, 33, 171–177. [Google Scholar]
- Brisson, J.A.; Strasburg, J.L.; Templeton, A.R. Impact of fire management on the ecology of collared lizard (Crotaphytus collaris) populations living on the Ozark Plateau. Anim. Conserv. 2003, 6, 247–254. [Google Scholar] [CrossRef]
- Young, C.C.; Morrison, L.W.; Haack, J.L. Habitat relationships and management implications for Lesquerella filiformis Rollins (missouri bladderpod) on a xeric limestone prairie. J. Torrey Bot. Soc. 2009, 136, 233–241. [Google Scholar] [CrossRef]
- Templeton, A.R.; Neuwald, J.L.; Brazeal, H.; Robertson, R.J. Invited Minireview: Restoring demographic processes in translocated populations: The case of collared lizards in the Missouri Ozarks using prescribed forest fires. Isr. J. Ecol. Evol. 2007, 53, 179–196. [Google Scholar] [CrossRef]
- Barger, N.N.; Archer, S.R.; Campbell, J.L.; Huang, C.Y.; Morton, J.A.; Knapp, A.K. Woody plant proliferation in North American drylands: A synthesis of impacts on ecosystem carbon balance. J. Geophys. Res. Biogeosci. 2011, 116. [Google Scholar] [CrossRef]
- Comer, C.E.; Bell, A.L.; Oswald, B.P.; Conway, W.C.; Burt, D.B. Vegetation and avian response to prescribed fire on glade habitats in the Missouri Ozarks. Am. Midl. Nat. 2010, 165, 91–104. [Google Scholar] [CrossRef]
- Miller, J.E.D.; Damschen, E.I.; Ratajczak, Z.; Özdoğan, M. Holding the line: Three decades of prescribed fires halt but do not reverse woody encroachment in grasslands. Landsc. Ecol. 2017, 32, 2297–2310. [Google Scholar] [CrossRef]
- McCarty, K. Landscape-scale restoration in Missouri savannas and woodlands. Restor. Manag. Notes 1998, 16, 22–32. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basal Area (m2/ha) | Density (Trees/ha) | Quadratic Mean Diameter (cm) | ||||
|---|---|---|---|---|---|---|
| Forest Type | Mean | SE | Mean | SE | Mean | SE |
| BL | 27.7 A | 0.96 | 477.9 B | 20.22 | 27.9 B | 0.80 |
| DRS | 23.0 B | 0.93 | 434.6 B | 25.96 | 26.7 B | 1.15 |
| GL | 19.1 C | 1.54 | 569.6 A | 23.11 | 20.7 C | 0.48 |
| MS | 27.3 A | 0.81 | 341.4 C | 8.85 | 32.1 A | 0.58 |
| p-value | <0.001 | <0.001 | <0.001 | |||
| Year | ||||||
| 1968 | 22.0 C | 2.28 | 487.0 | 38.90 | 24.7 C | 2.24 |
| 1982 | 22.2 C | 2.39 | 431.1 | 41.07 | 26.4 BC | 2.52 |
| 1992 | 24.3 BC | 2.28 | 472.4 | 56.14 | 26.4 BC | 2.37 |
| 2004 | 25.5 AB | 1.64 | 444.2 | 57.29 | 27.8 AB | 2.38 |
| 2016 | 27.4 A | 1.61 | 444.8 | 61.11 | 29.1 A | 2.40 |
| p-value | <0.001 | 0.246 | <0.001 | |||
| Axis 1 | Axis 2 | Axis 3 | ||||
|---|---|---|---|---|---|---|
| Rank | Species | r | Species | r | Species | r |
| Negative | ||||||
| 1 | Acer saccharum | −0.721 | Carya ovata | −0.623 | Juniperus virginiana | −0.775 |
| 2 | Ostrya virginiana | −0.659 | Carya texana | −0.620 | Fraxinus quadrangulata | −0.620 |
| 3 | Carpinus caroliniana | −0.566 | Quercus stellata | −0.574 | Sideroxylon lanuginosum | −0.609 |
| 4 | Cornus florida | −0.511 | Quercus shumardii | −0.509 | Quercus muehlenbergii | −0.561 |
| 5 | Cercis canadensis | −0.475 | Quercus muehlenbergii | −0.448 | Quercus shumardii | −0.388 |
| Positive | ||||||
| 1 | Quercus alba | 0.736 | Aesculus glabra | 0.692 | Acer saccharum | 0.410 |
| 2 | Quercus velutina | 0.546 | Acer nigrum | 0.610 | Asimina triloba | 0.396 |
| 3 | Juglans nigra | 0.542 | Acer negundo | 0.543 | Cornus florida | 0.384 |
| 4 | Quercus rubra | 0.542 | Ulmus americana | 0.512 | Quercus alba | 0.357 |
| 5 | Carya laciniosa | 0.527 | Celtis occidentalis | 0.504 | Carya tomentosa | 0.304 |
| Species | Size Class | 1968 | 1982 | 1992 | 2004 | 2016 |
|---|---|---|---|---|---|---|
| Dry Ridge and Slope | ||||||
| Acer saccharum | Class 1 | 505 | 501 | 344 | 54 | 6 |
| Class 2 | 484 | 605 | 799 | 425 | 217 | |
| Class 3 | 78 | 192 | 259 | 253 | 250 | |
| Class 4 | 35 | 58 | 84 | 133 | 116 | |
| Quercus alba | Class 1 | 50 | 20 | 22 | 0 | 0 |
| Class 2 | 104 | 8 | 15 | 0 | 0 | |
| Class 3 | 134 | 25 | 19 | 5 | 4 | |
| Class 4 | 83 | 39 | 20 | 6 | 0 | |
| Quercus shumardii | Class 1 | 17 | 3 | 1 | 0 | 0 |
| Class 2 | 11 | 1 | 2 | 0 | 0 | |
| Class 3 | 1 | 0 | 1 | 0 | 0 | |
| Class 4 | 0 | 0 | 0 | 0 | 0 | |
| Quercus velutina | Class 1 | 27 | 20 | 42 | 6 | 0 |
| Class 2 | 16 | 1 | 13 | 10 | 0 | |
| Class 3 | 6 | 2 | 3 | 0 | 0 | |
| Class 4 | 1 | 0 | 2 | 0 | 0 | |
| Glade-Like | ||||||
| Juniperus virginiana | Class 1 | 466 | 250 | 194 | 53 | 0 |
| Class 2 | 144 | 166 | 259 | 163 | 69 | |
| Class 3 | 41 | 106 | 166 | 106 | 79 | |
| Class 4 | 13 | 72 | 91 | 100 | 43 | |
| Quercus muehlenbergii | Class 1 | 66 | 41 | 38 | 0 | 0 |
| Class 2 | 103 | 34 | 28 | 13 | 0 | |
| Class 3 | 75 | 34 | 31 | 38 | 7 | |
| Class 4 | 53 | 34 | 47 | 28 | 7 | |
| Quercus shumardii | Class 1 | 25 | 25 | 13 | 0 | 0 |
| Class 2 | 13 | 9 | 9 | 0 | 0 | |
| Class 3 | 6 | 3 | 3 | 0 | 0 | |
| Class 4 | 13 | 6 | 0 | 0 | 0 | |
| Mesic Slope | ||||||
| Acer saccharum | Class 1 | 525 | 280 | 325 | 55 | 6 |
| Class 2 | 1050 | 533 | 700 | 278 | 115 | |
| Class 3 | 178 | 290 | 320 | 210 | 113 | |
| Class 4 | 23 | 115 | 136 | 125 | 115 | |
| Quercus alba | Class 1 | 3 | 0 | 5 | 0 | 0 |
| Class 2 | 3 | 0 | 0 | 0 | 0 | |
| Class 3 | 3 | 0 | 0 | 0 | 0 | |
| Class 4 | 3 | 0 | 0 | 0 | 0 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
O. Knapp, B.; G. Pallardy, S. Forty-Eight Years of Forest Succession: Tree Species Change across Four Forest Types in Mid-Missouri. Forests 2018, 9, 633. https://doi.org/10.3390/f9100633
O. Knapp B, G. Pallardy S. Forty-Eight Years of Forest Succession: Tree Species Change across Four Forest Types in Mid-Missouri. Forests. 2018; 9(10):633. https://doi.org/10.3390/f9100633
Chicago/Turabian StyleO. Knapp, Benjamin, and Stephen G. Pallardy. 2018. "Forty-Eight Years of Forest Succession: Tree Species Change across Four Forest Types in Mid-Missouri" Forests 9, no. 10: 633. https://doi.org/10.3390/f9100633
APA StyleO. Knapp, B., & G. Pallardy, S. (2018). Forty-Eight Years of Forest Succession: Tree Species Change across Four Forest Types in Mid-Missouri. Forests, 9(10), 633. https://doi.org/10.3390/f9100633