
Tree Species Site Suitability as a Combination of Occurrence Probability and Growth and Derivation of Priority Regions for Climate Change Adaptation

Abstract
:1. Introduction
2. Materials and Methods
2.1. National Forest Inventory Data
- : Basal area of a single tree at time t
- : Diameter at breast height at time t
- : Annual relative basal area increment
- : Basal area of a single tree at time t
- : Measuring time (here: NFI 1, NFI 2)
- : Exact time interval in days between the measurement dates divided by 365.25
- c: Slope correction factor
- : Slope
- : Stand basal area
- N: Number of trees of an inventory plot
- k: Basal area factor
- c: Slope correction factor
2.2. Environmental Data
2.3. Species Distribution Models
- p: Occurrence of a tree species (presence, absence; binary)
- T: Mean annual temperature or other temperature-related BioClim variable
- P: Mean annual precipitation sum or other precipitation-related BioClim variable
- S: Soil type (factor with 12 levels)
- i: Index of summation
- : Parameters
- : Thin plate splines with basis dimension k = 3
- : Thin plate spline for interaction with basis dimension k = 5
- : Error (binomial distribution)
2.4. Growth Models
- : Annual relative basal area increment
- T: Mean annual temperature or other temperature-related BioClim variable
- P: Mean annual precipitation sum or other precipitation-related BioClim variable
- S: Soil type (factor with 12 levels)
- i: Index of summation
- : Diameter at breast height
- : Stand basal area (or stand basal area of larger trees)
- : Parameters
- : Thin plate splines with basis dimension k = 3
- : Thin plate spline for interaction with basis dimension k = 5
- : Error (binomial distribution)
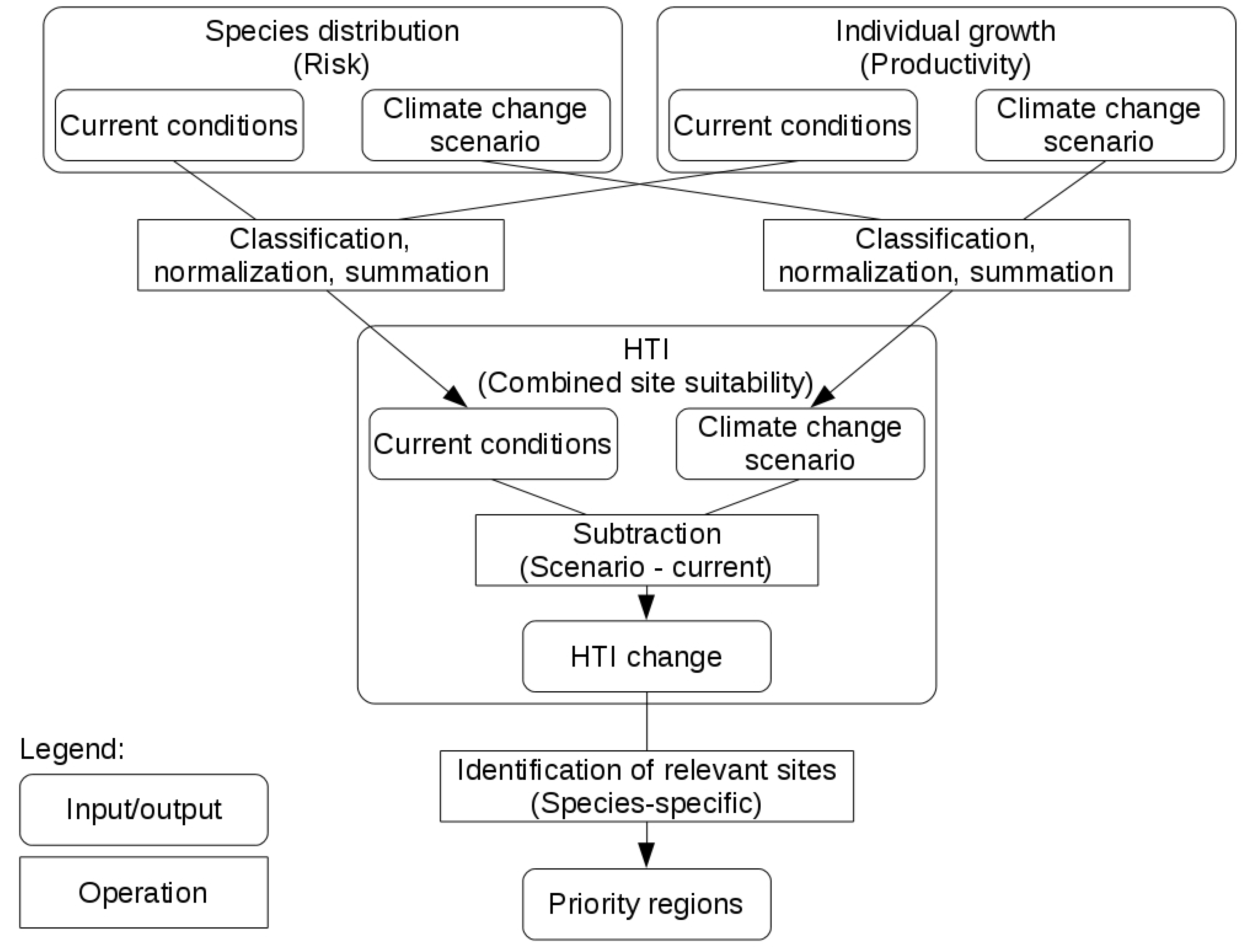
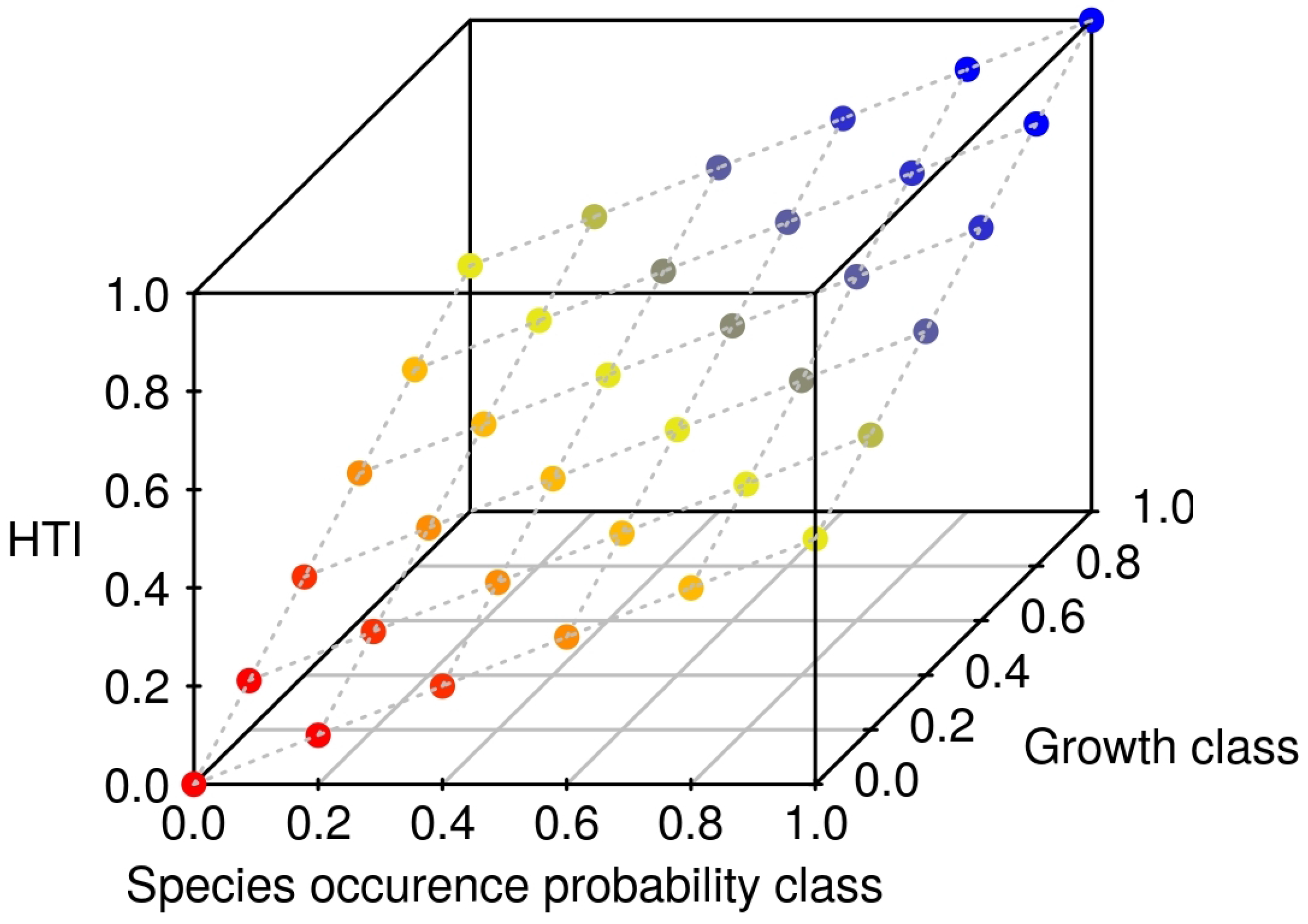
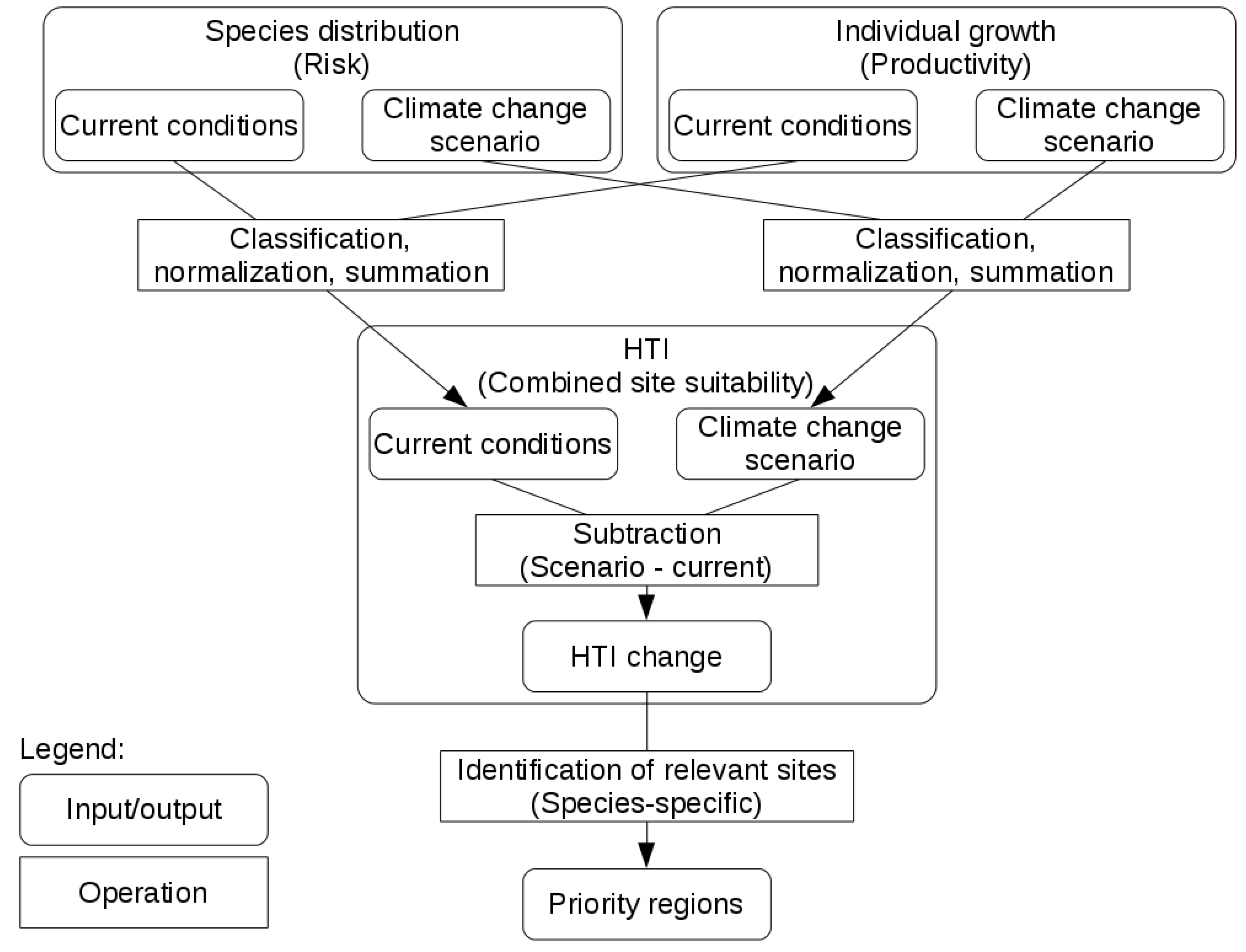
2.5. Combining Distribution and Growth
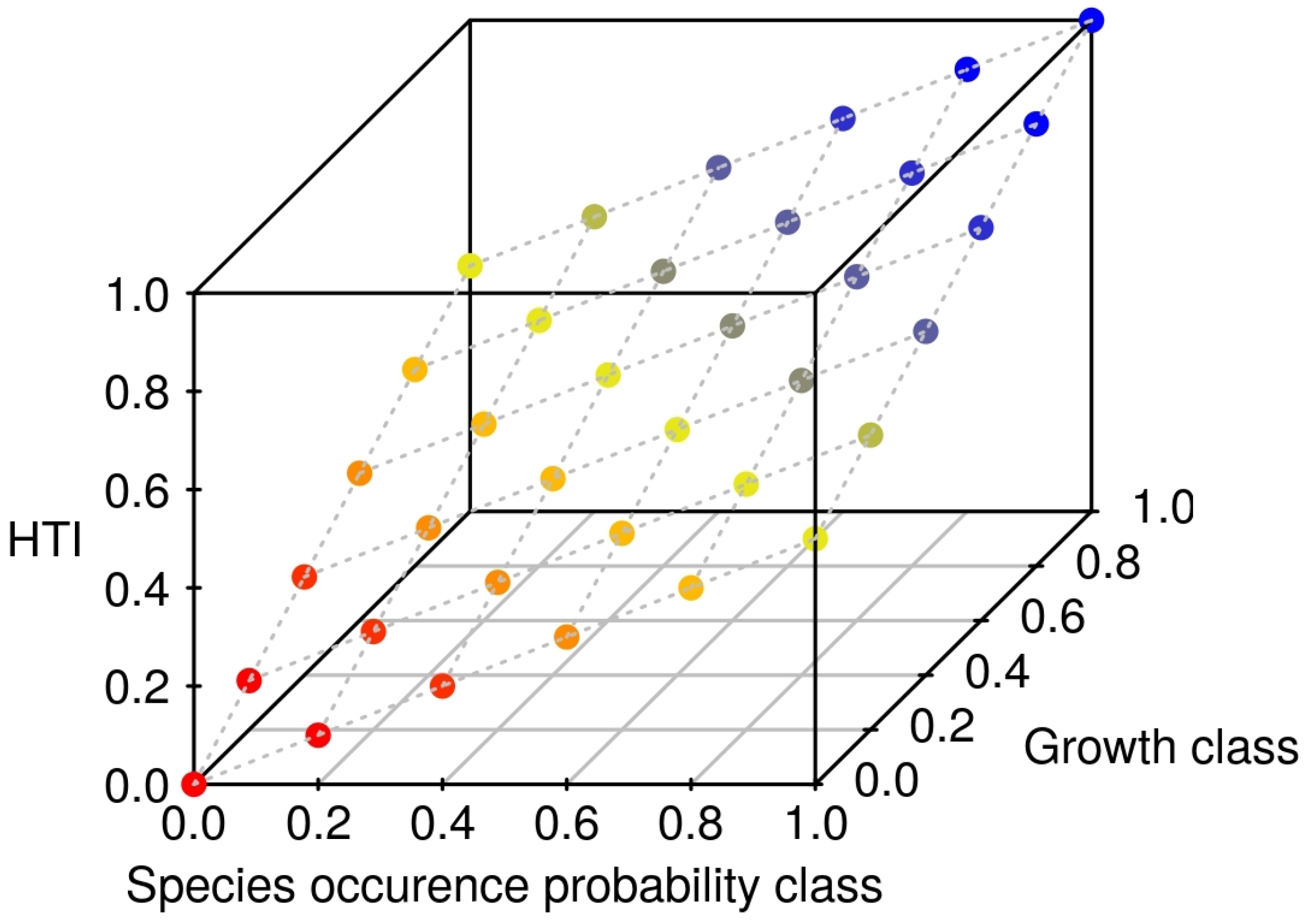
2.6. HTI Changes
2.7. Priority Regions
3. Results
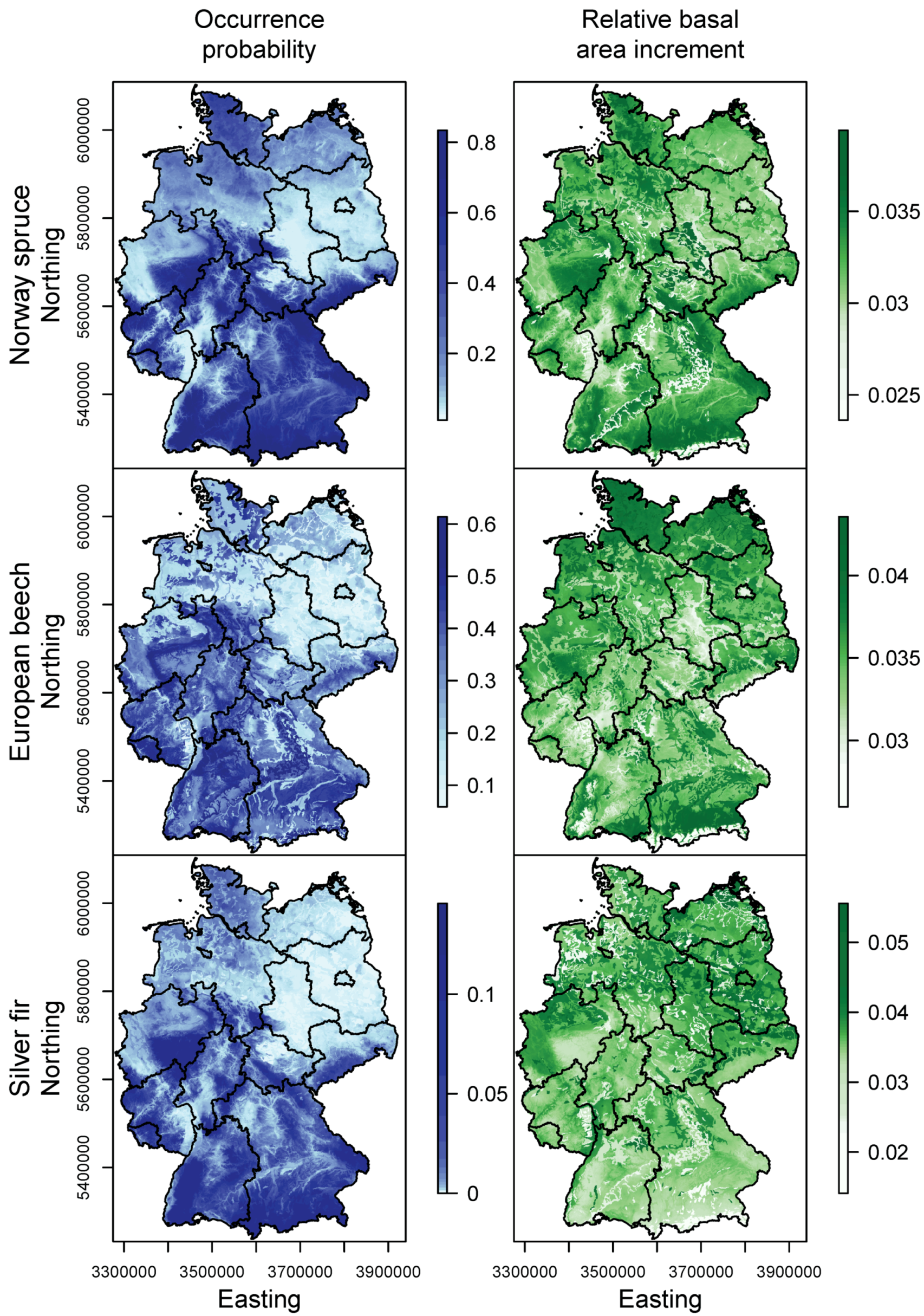
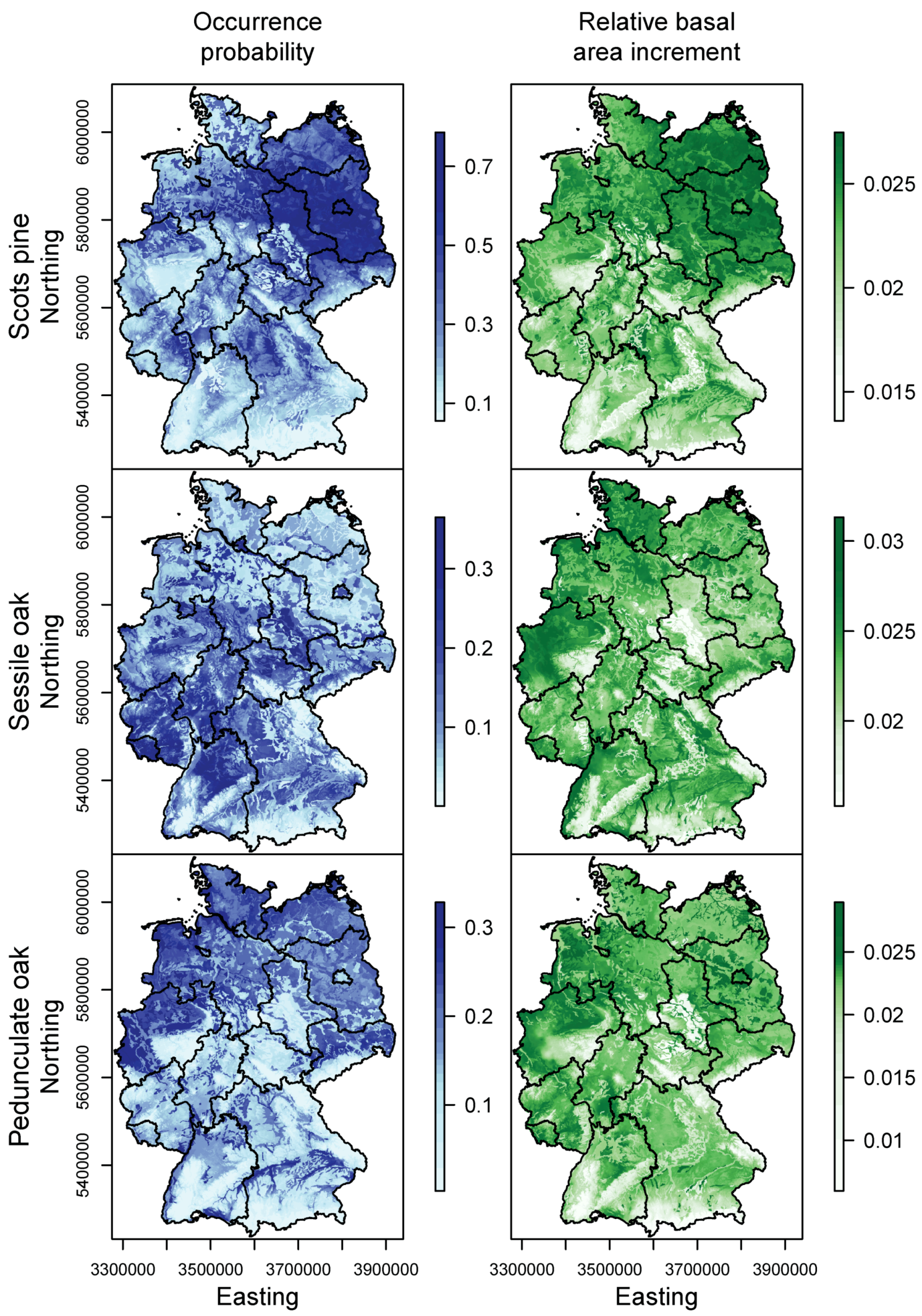
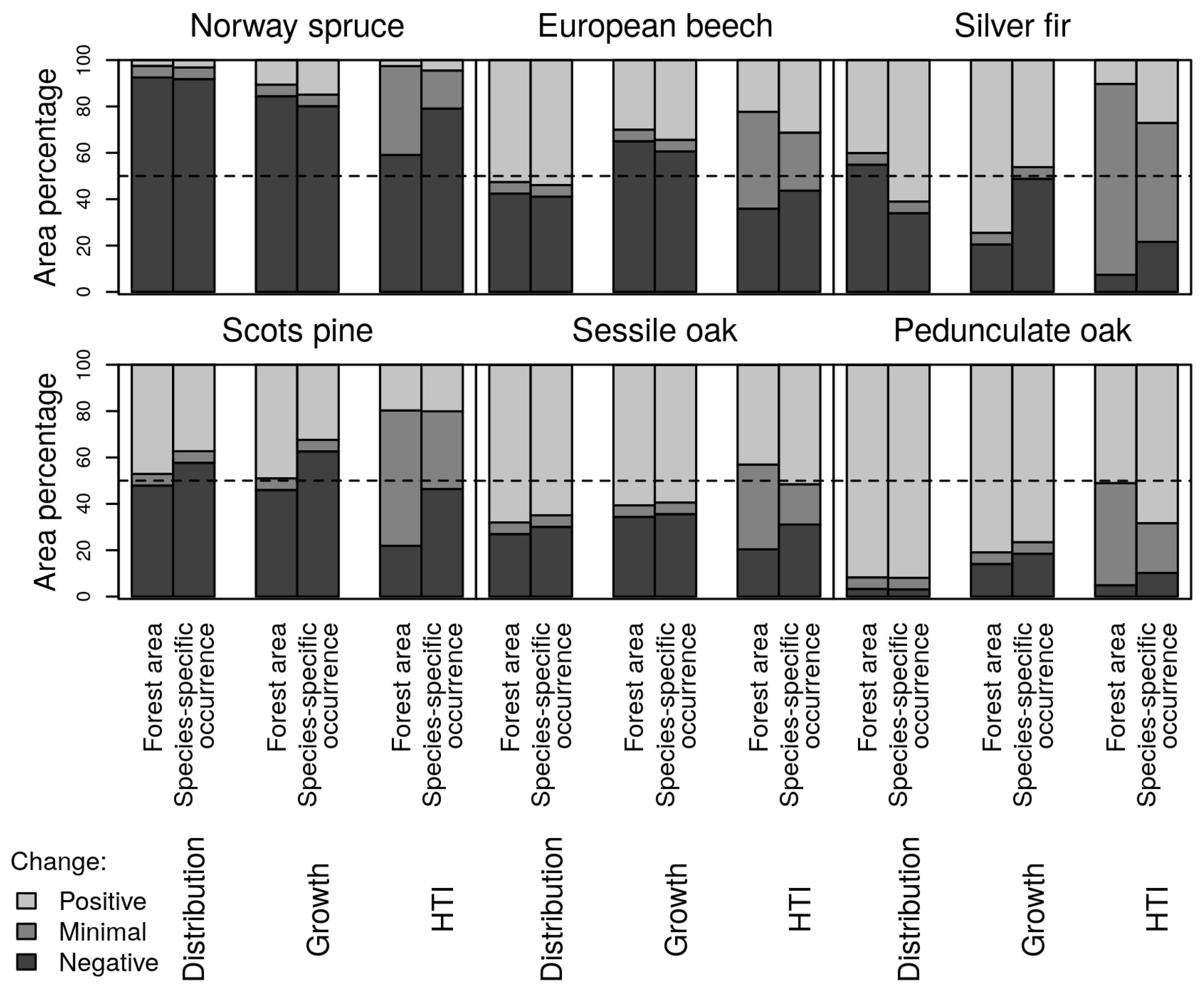
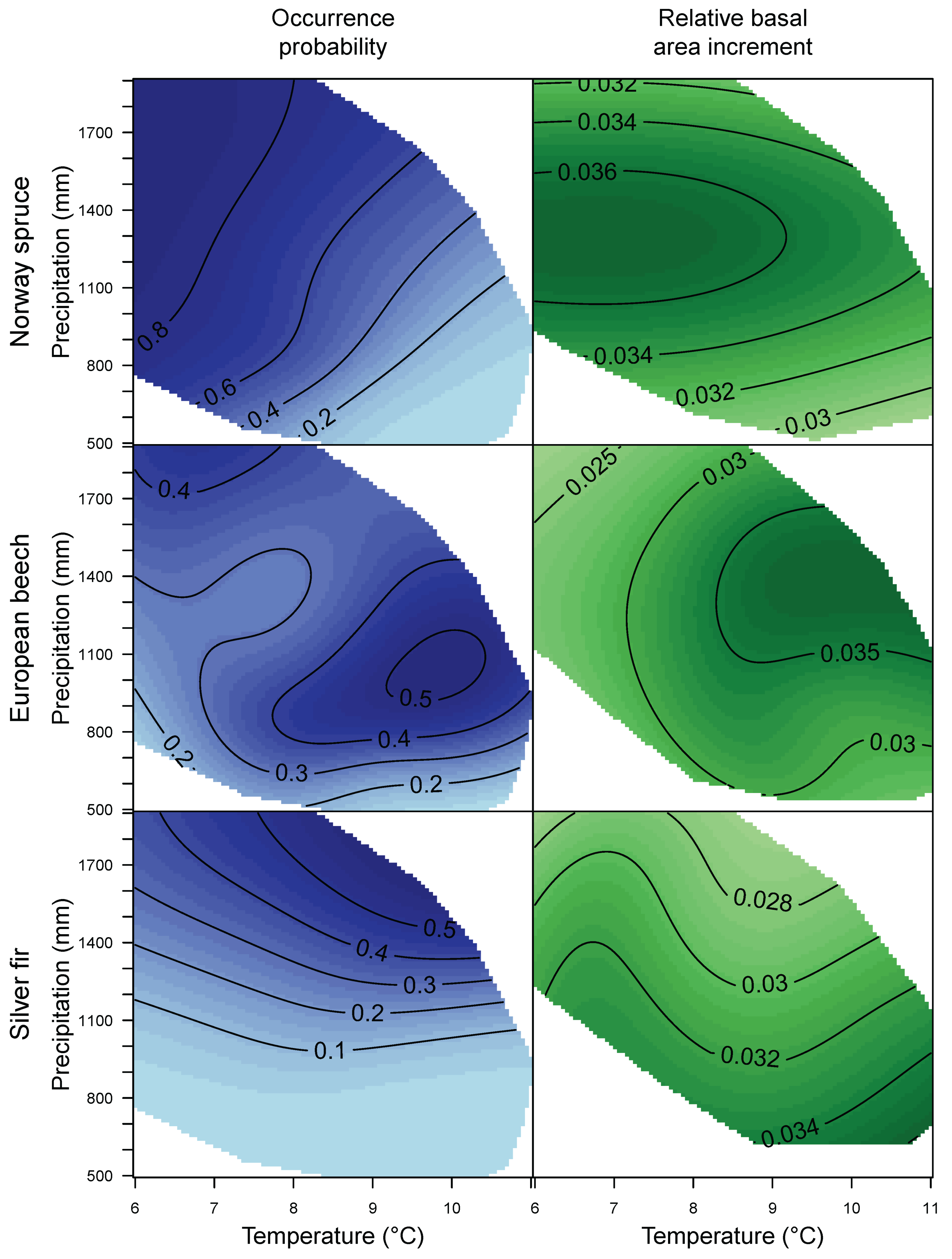
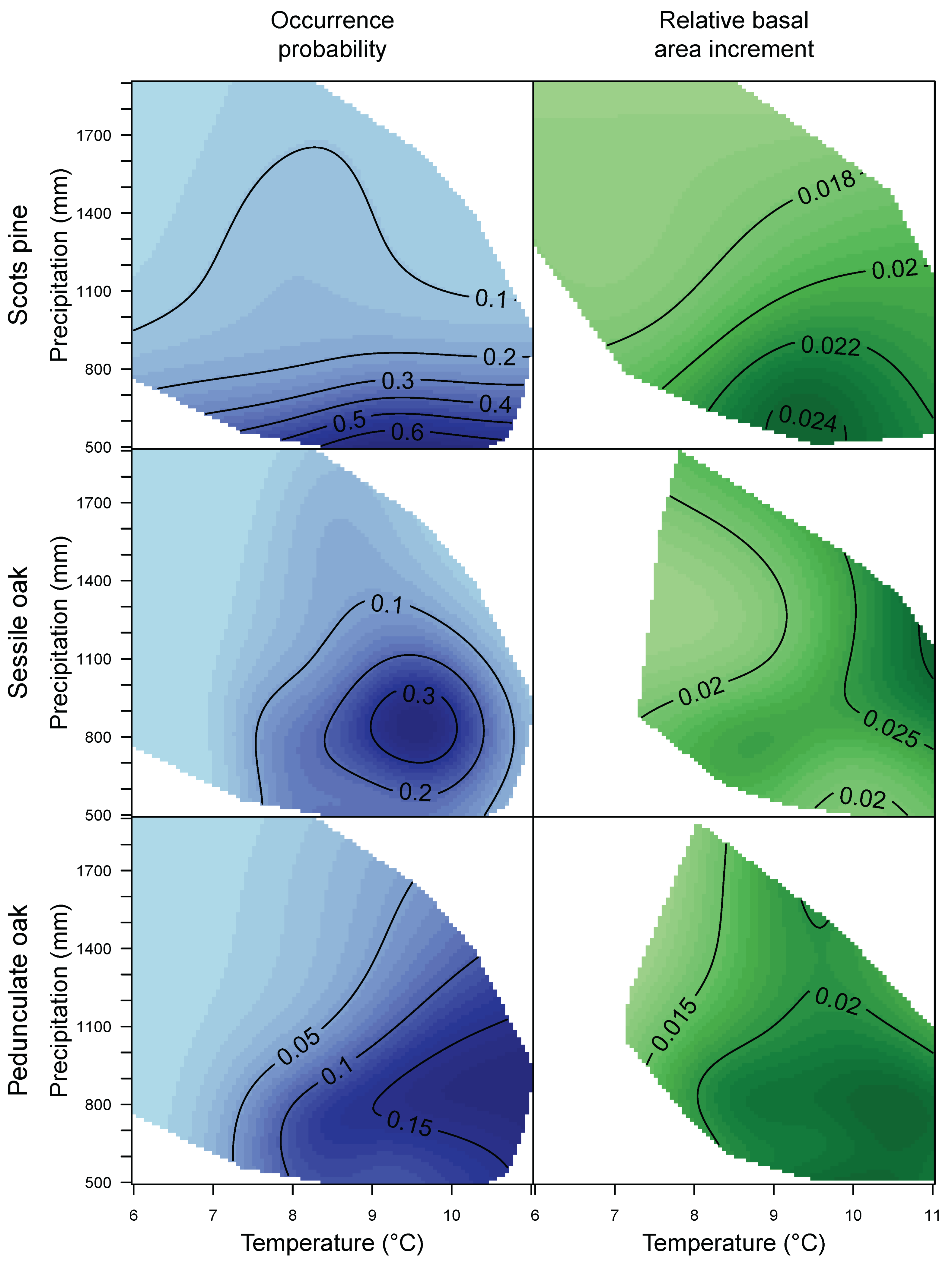
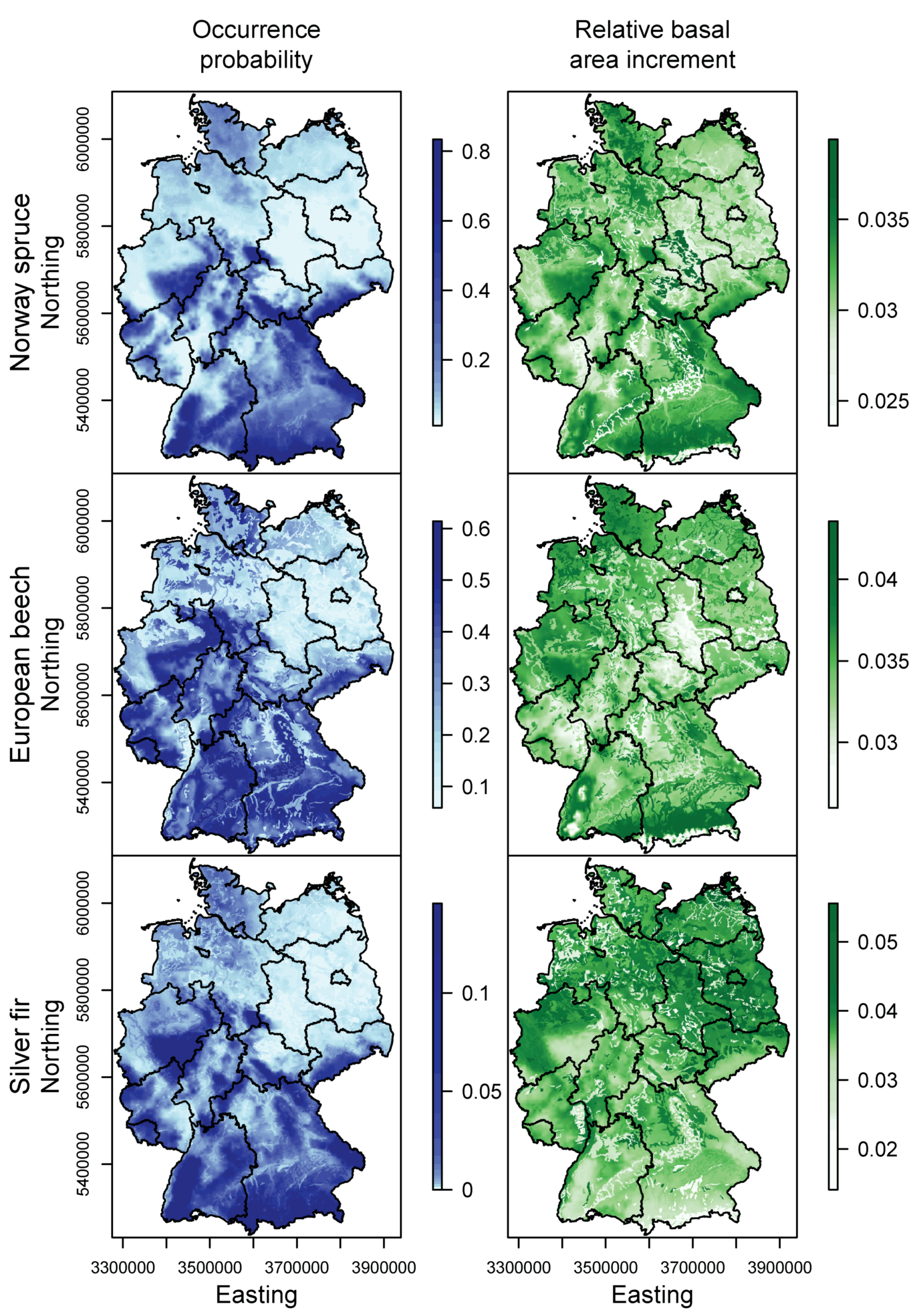
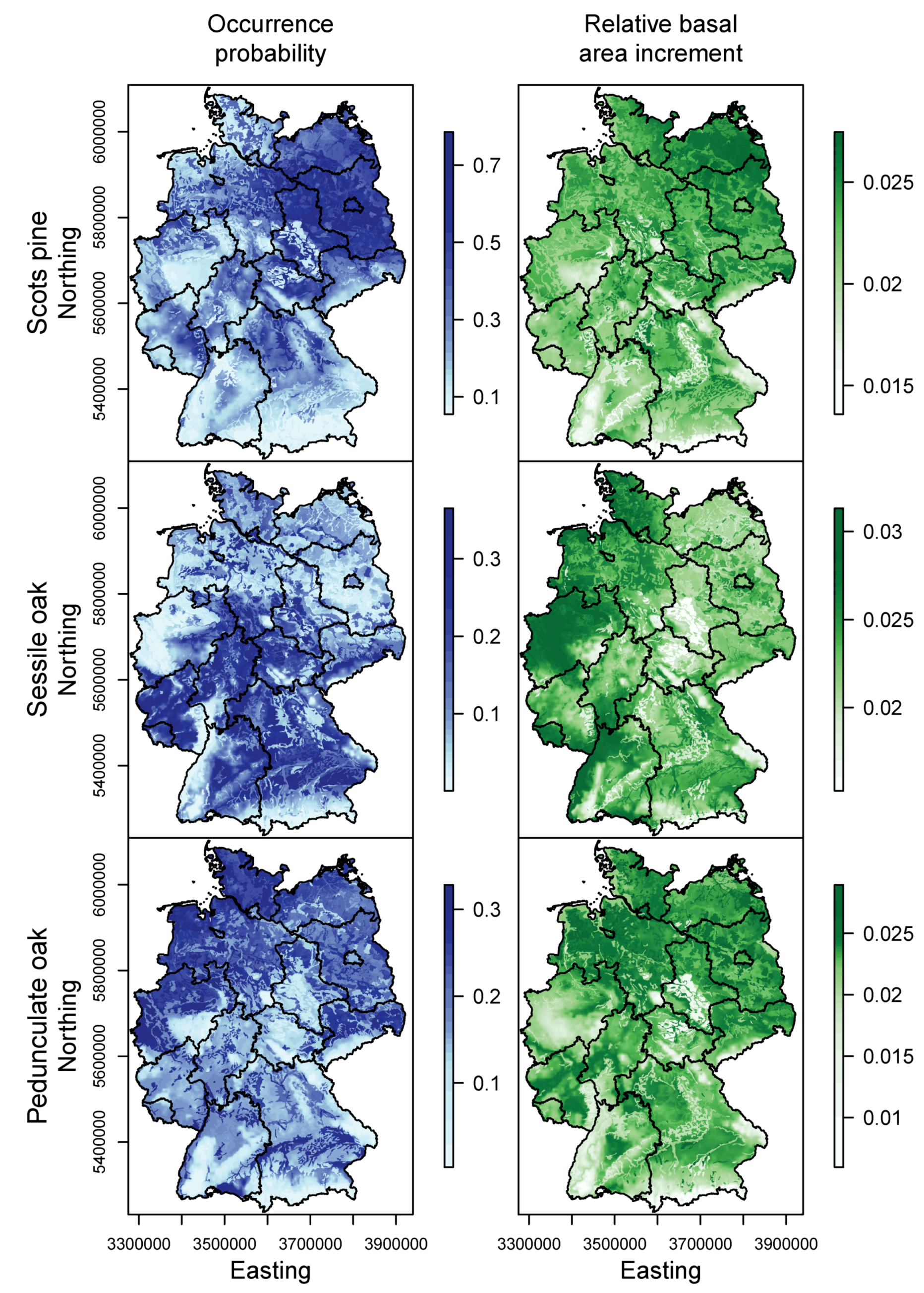
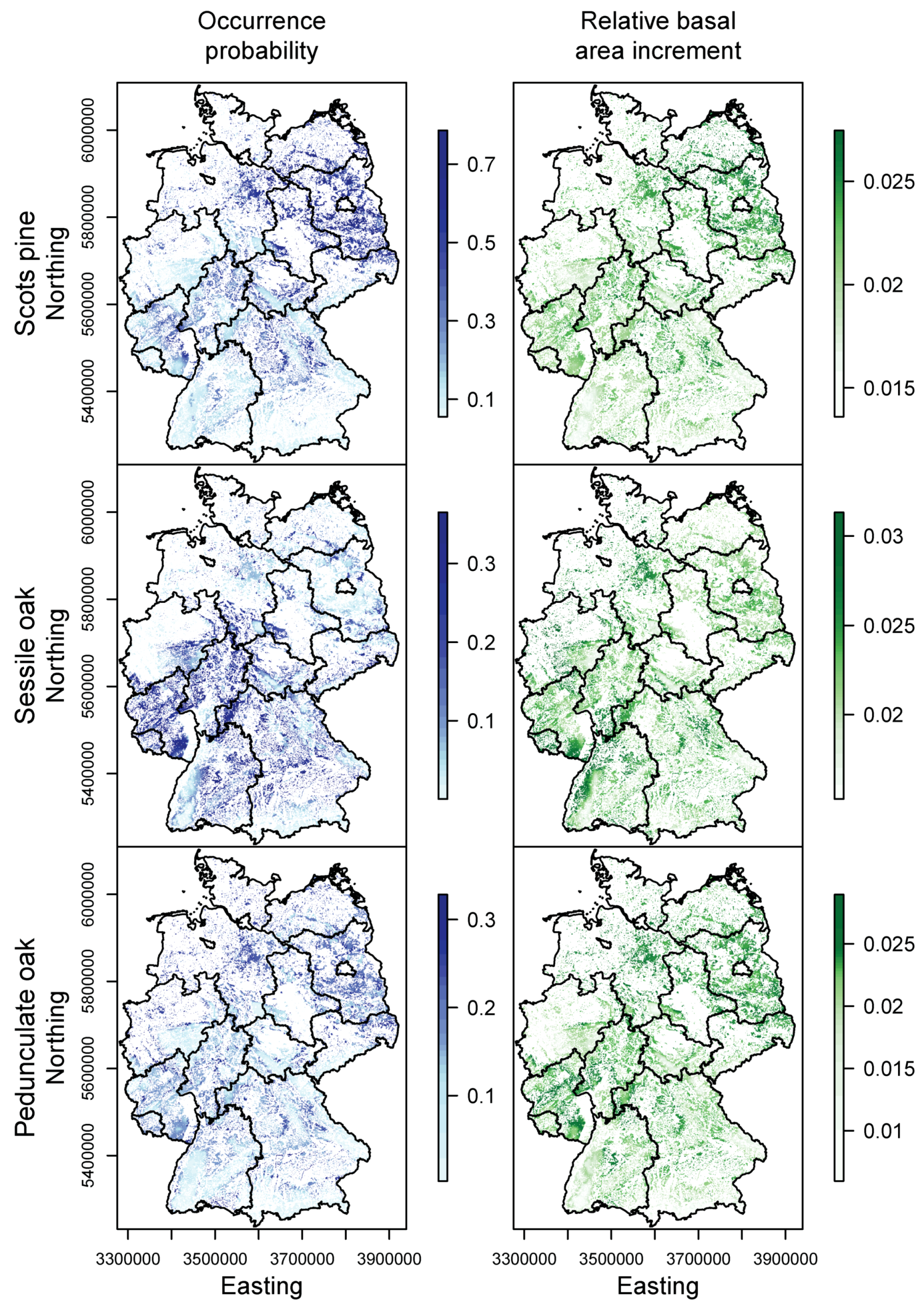
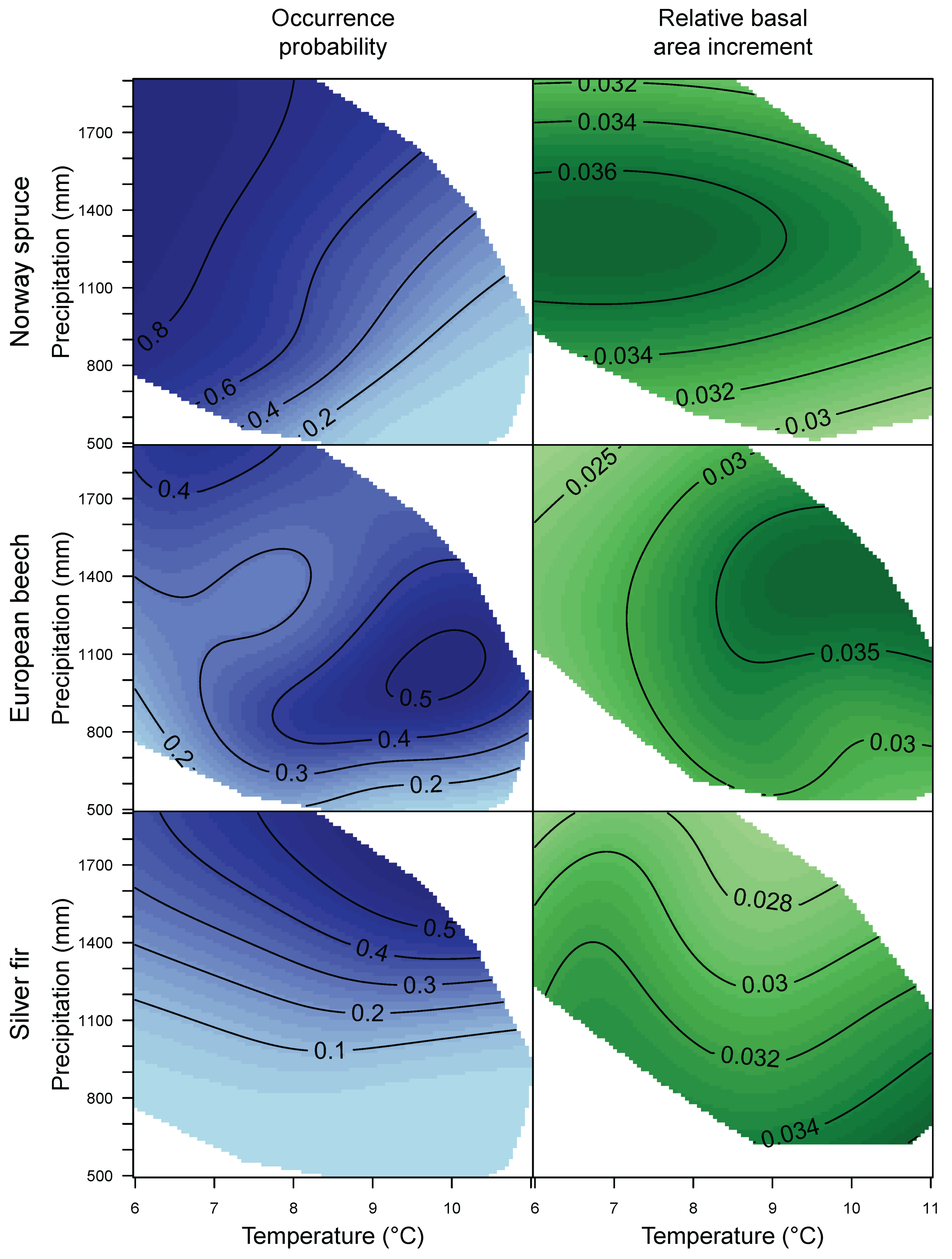
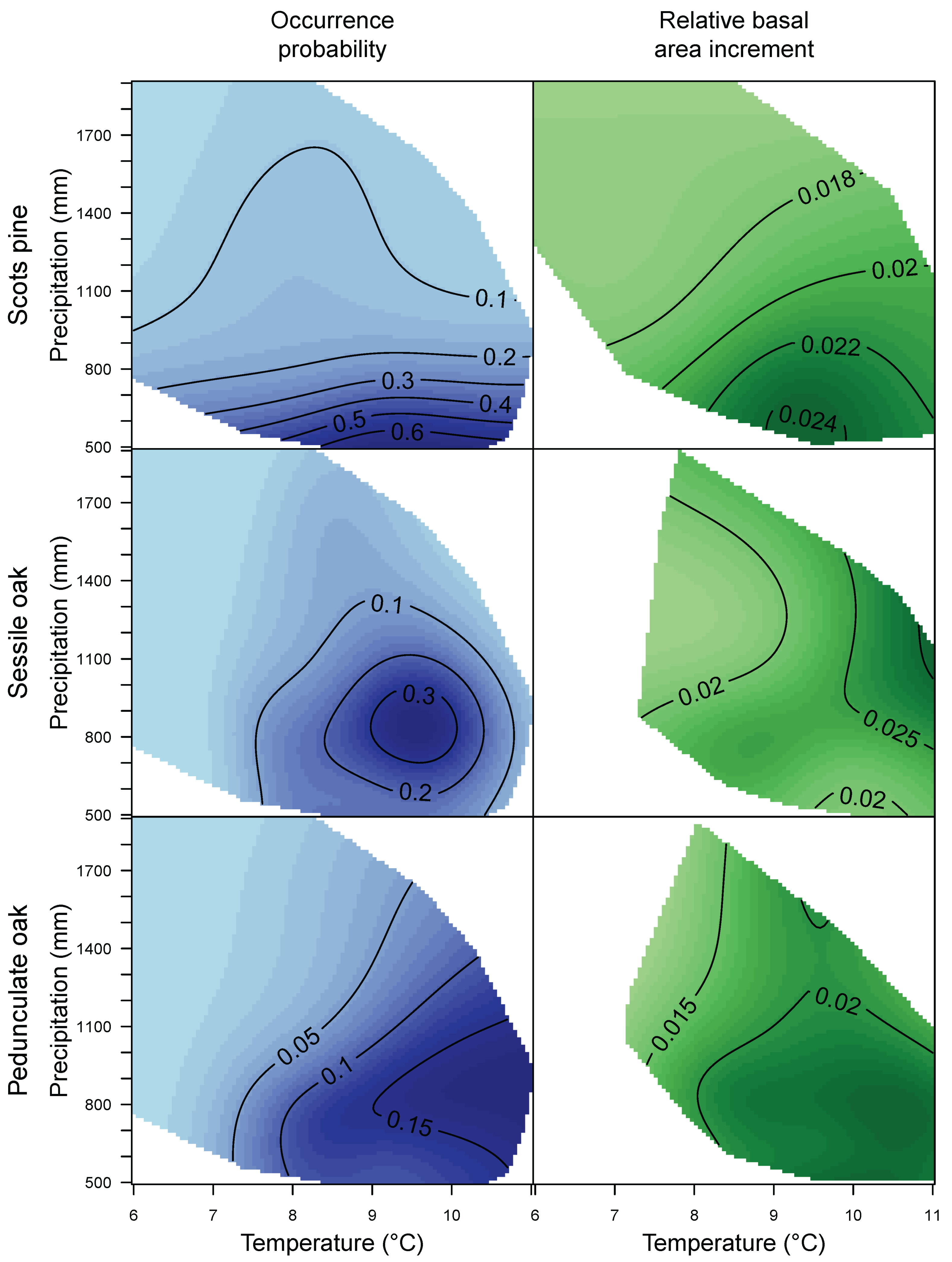
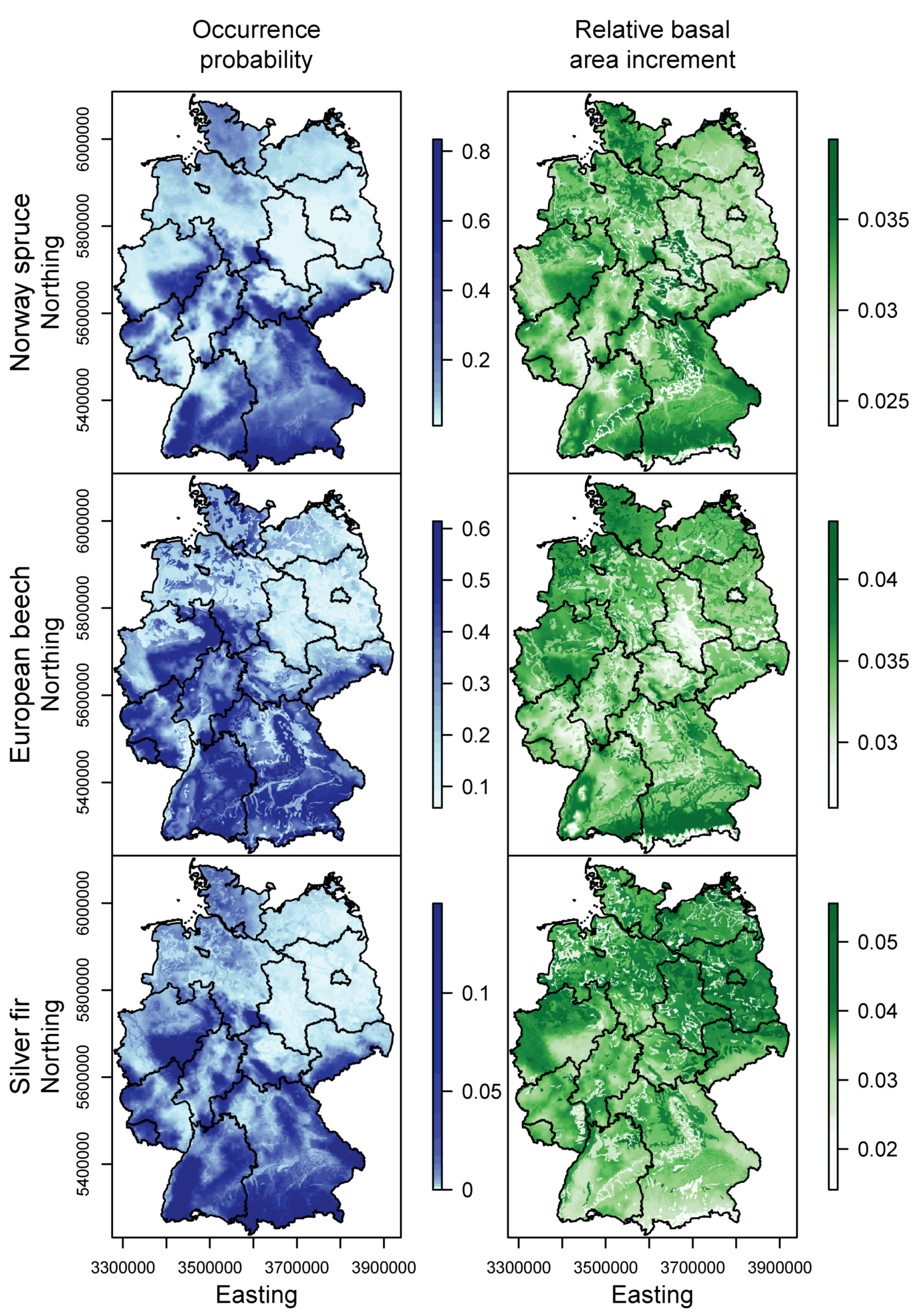
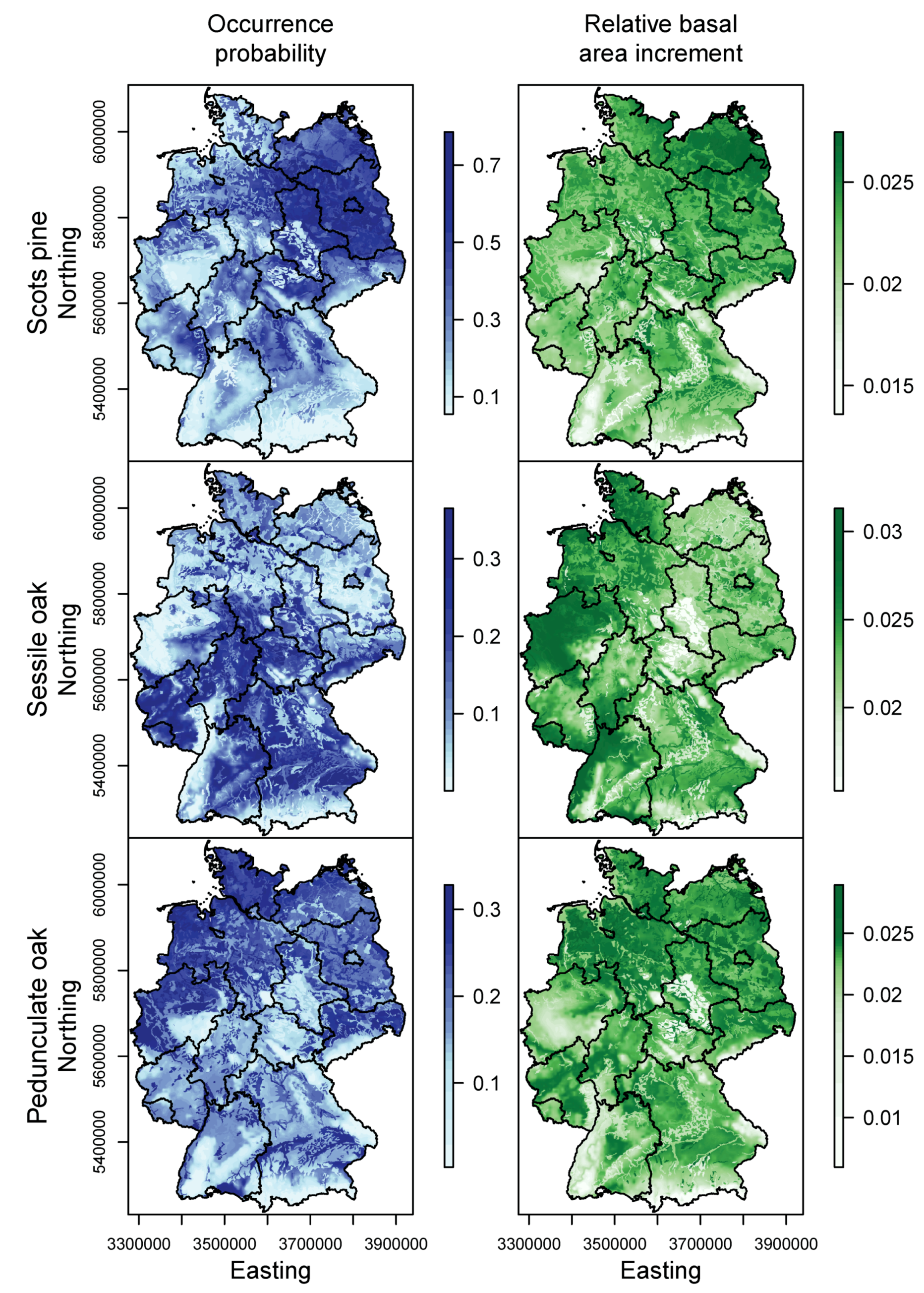
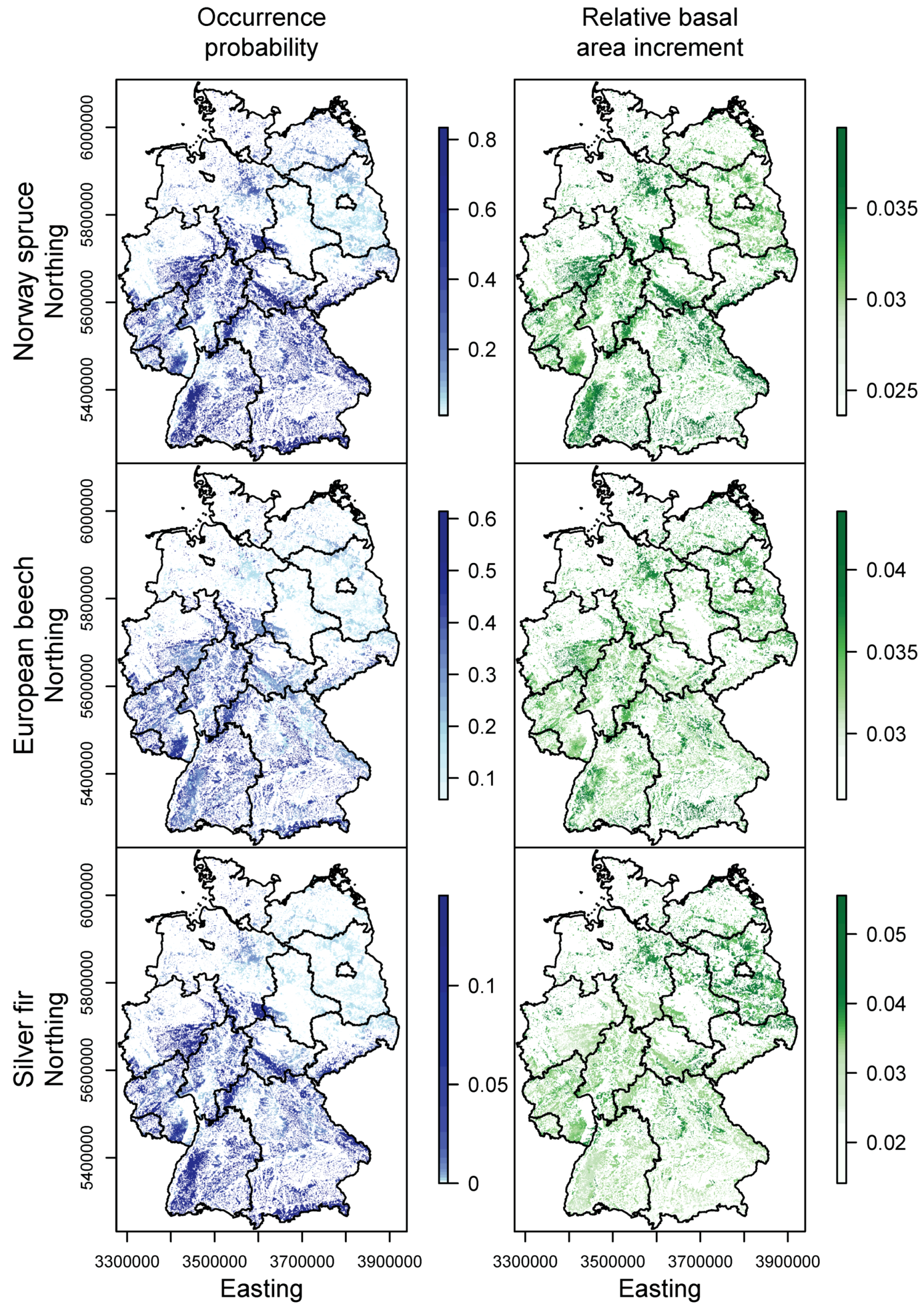
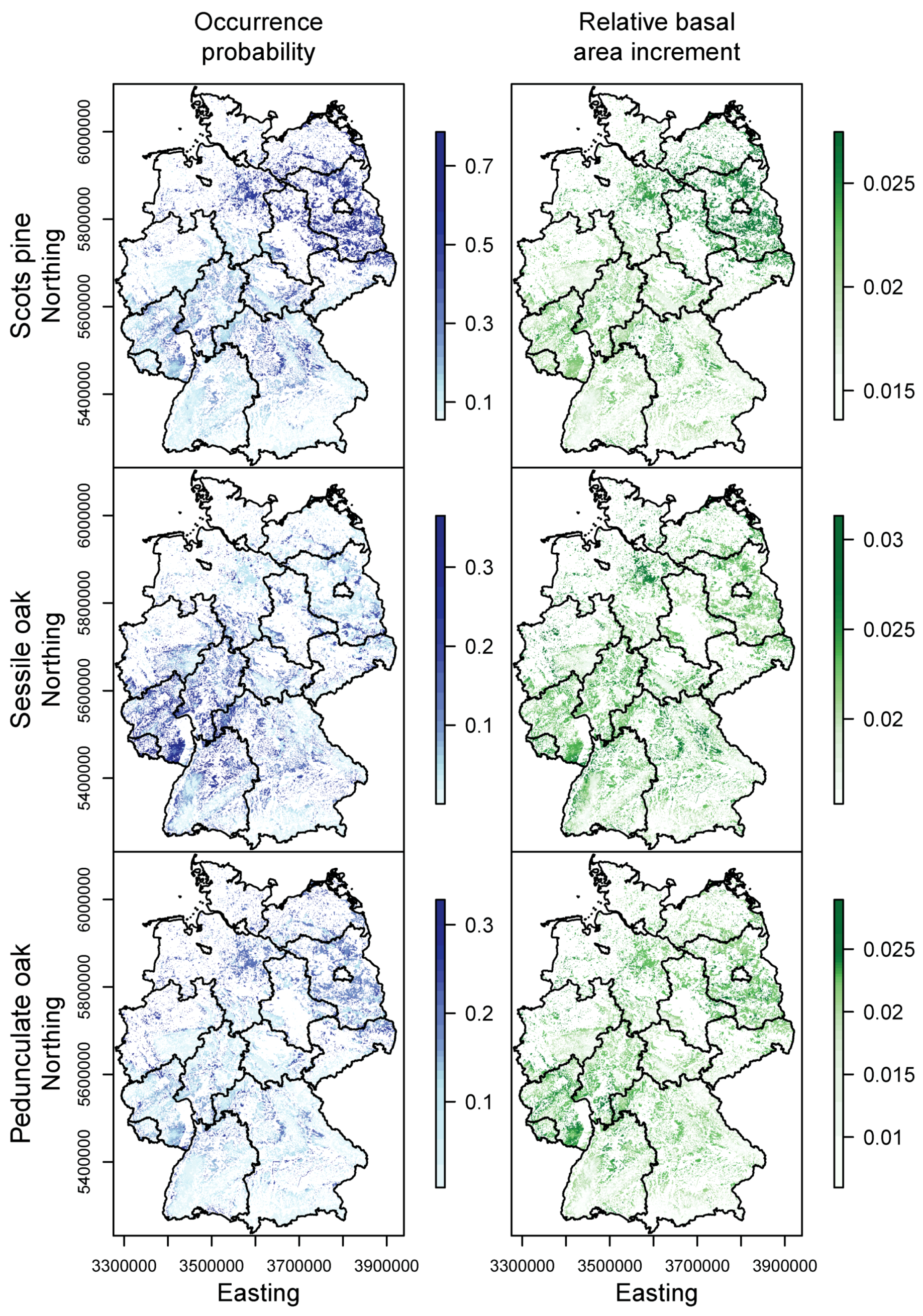
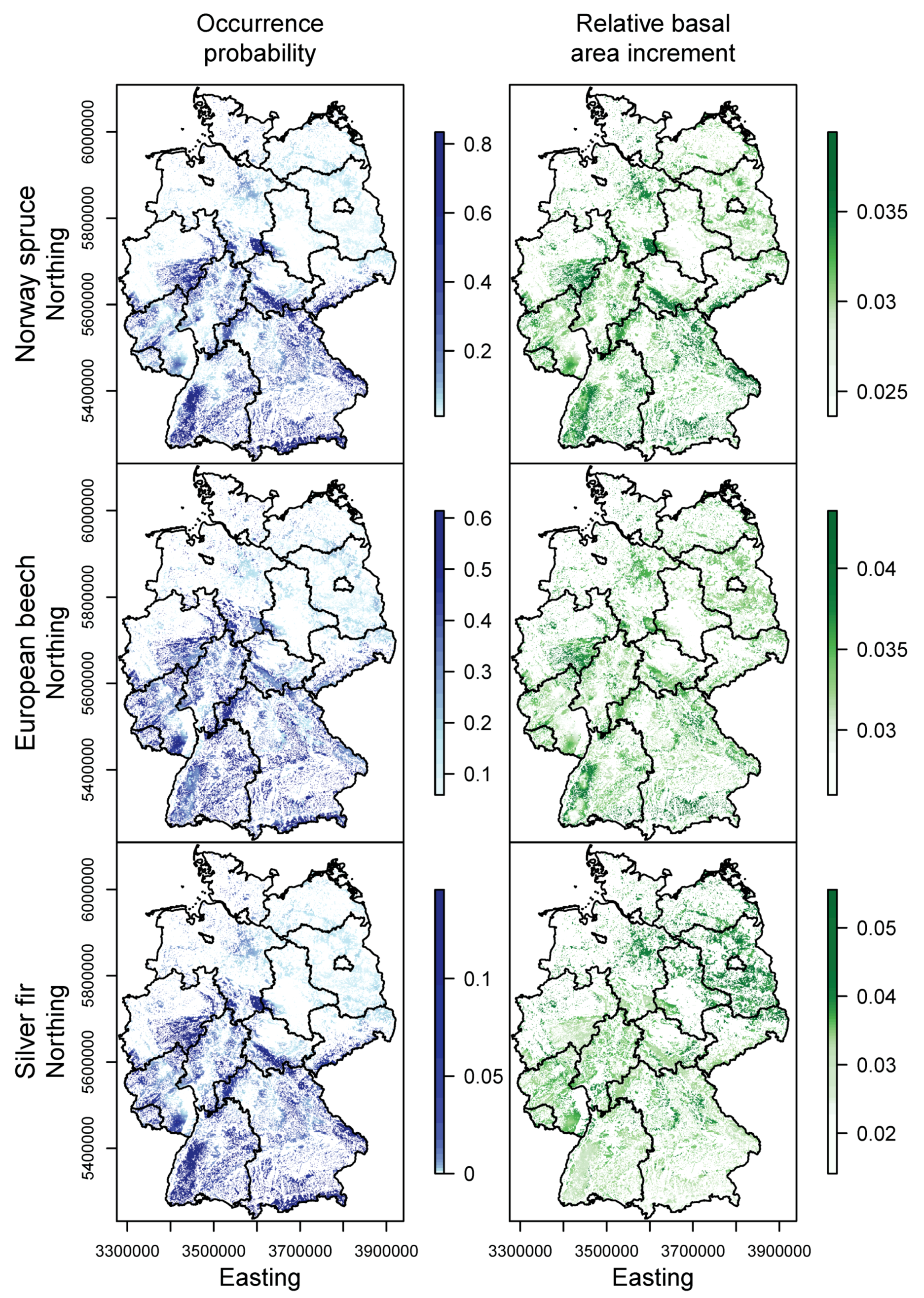
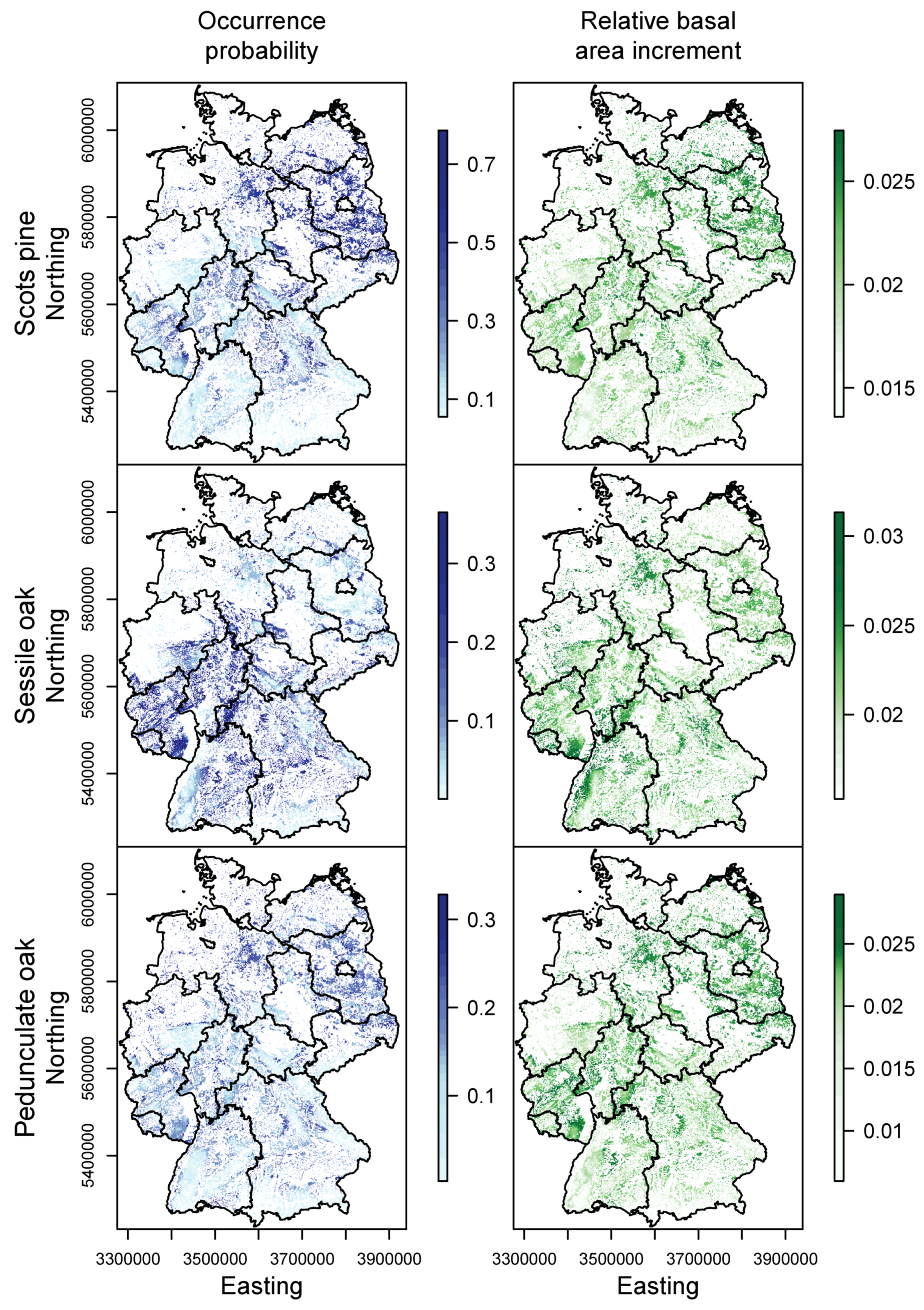
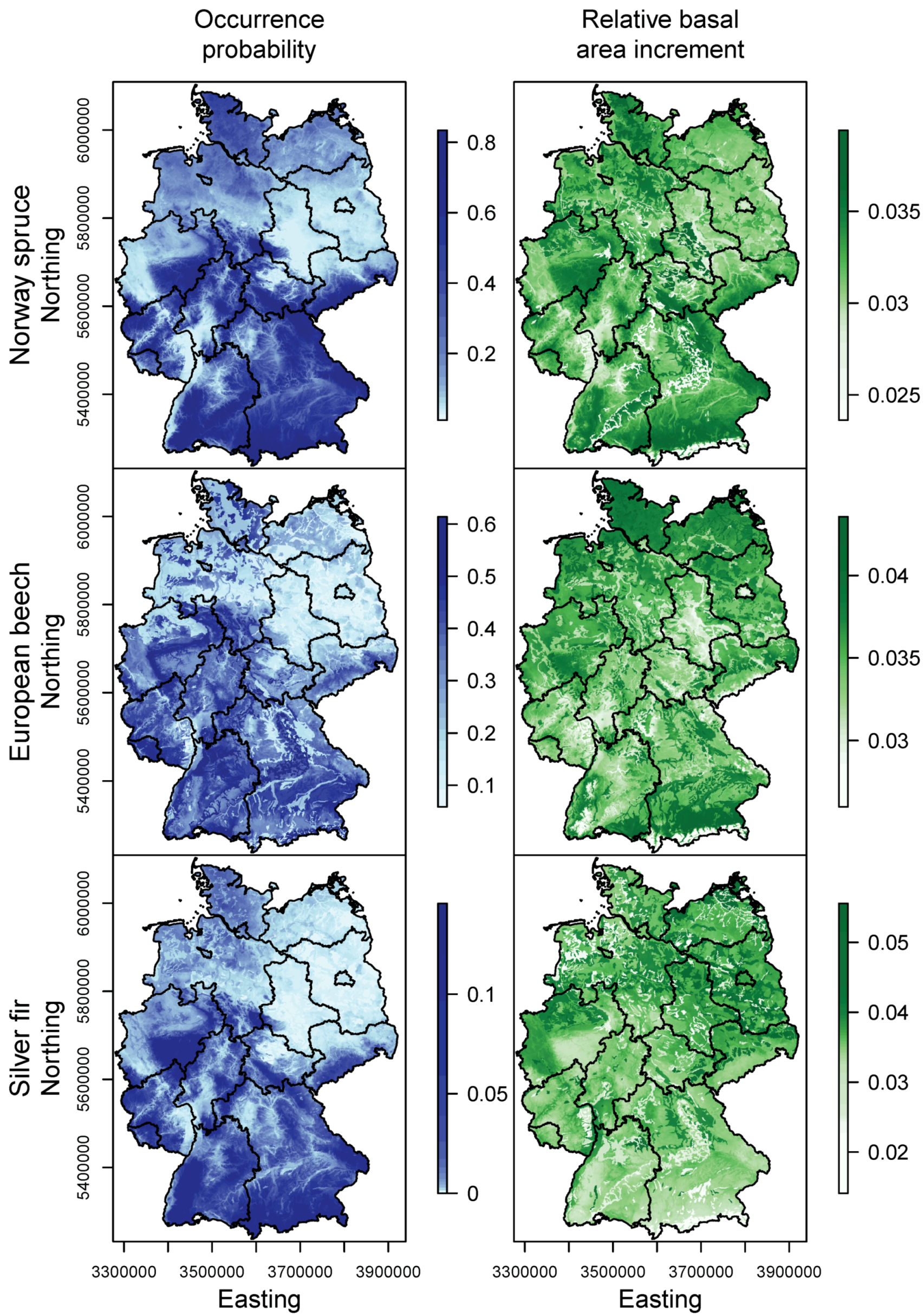
3.1. Species Distribution and Tree Growth
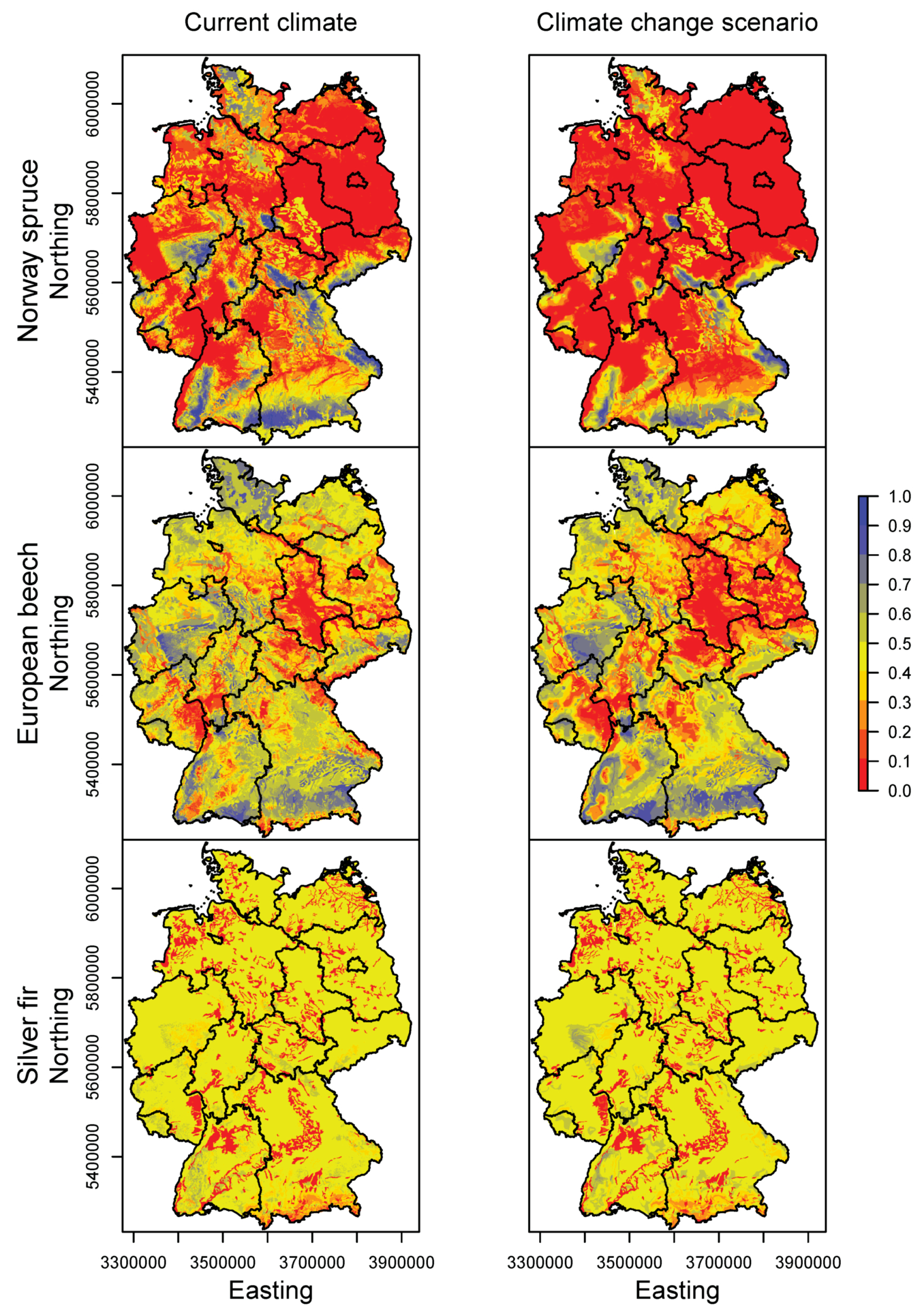
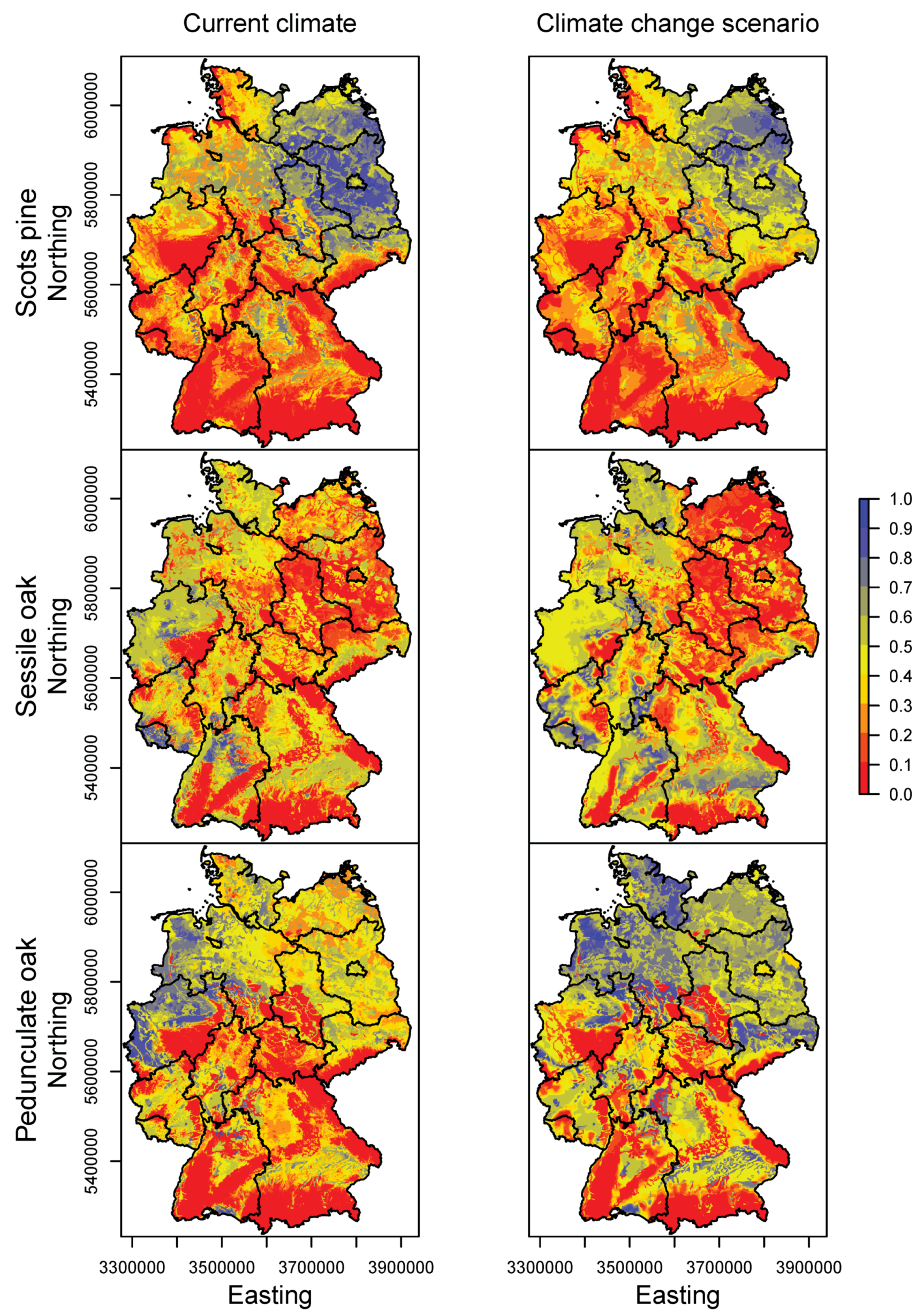
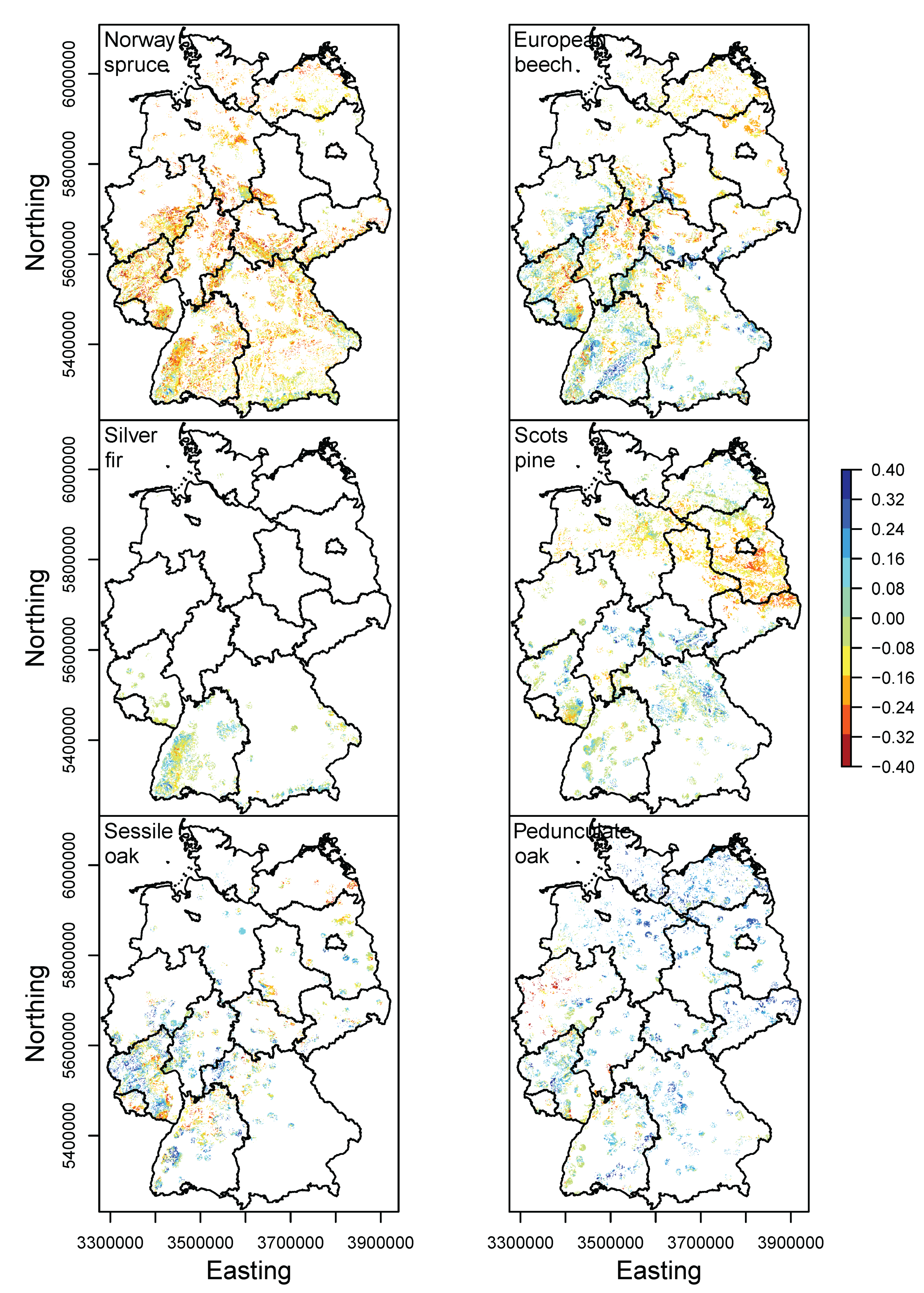
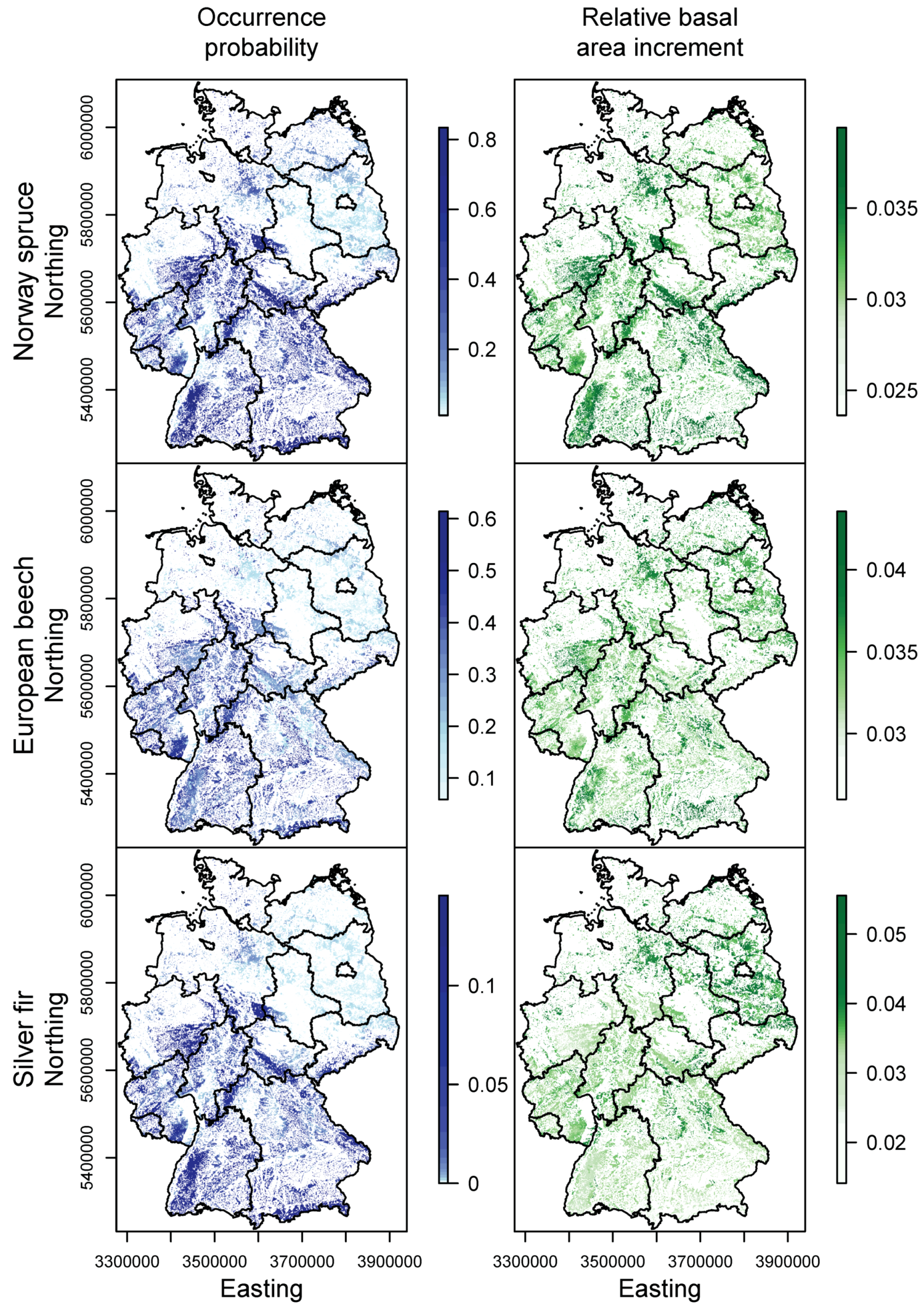
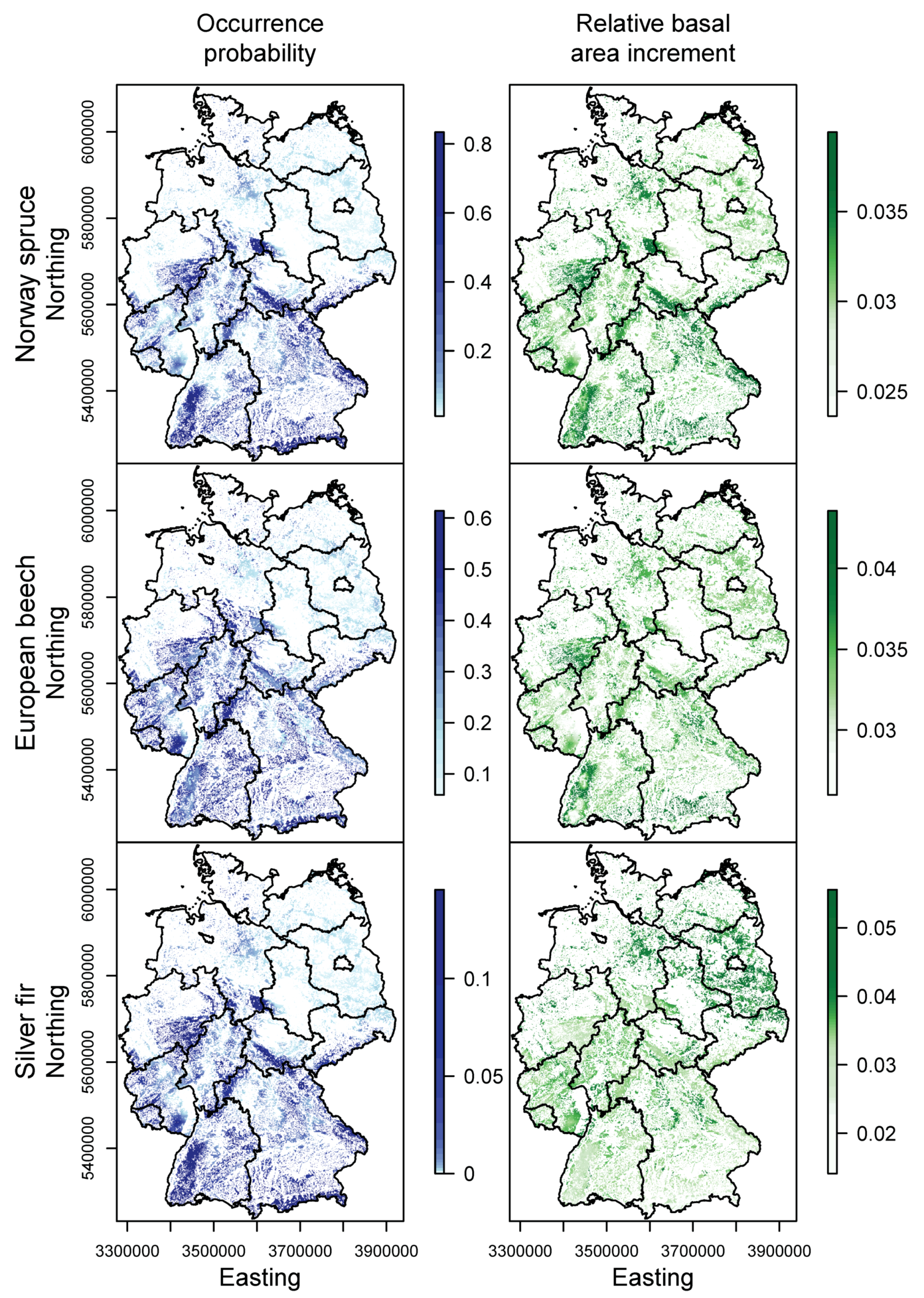
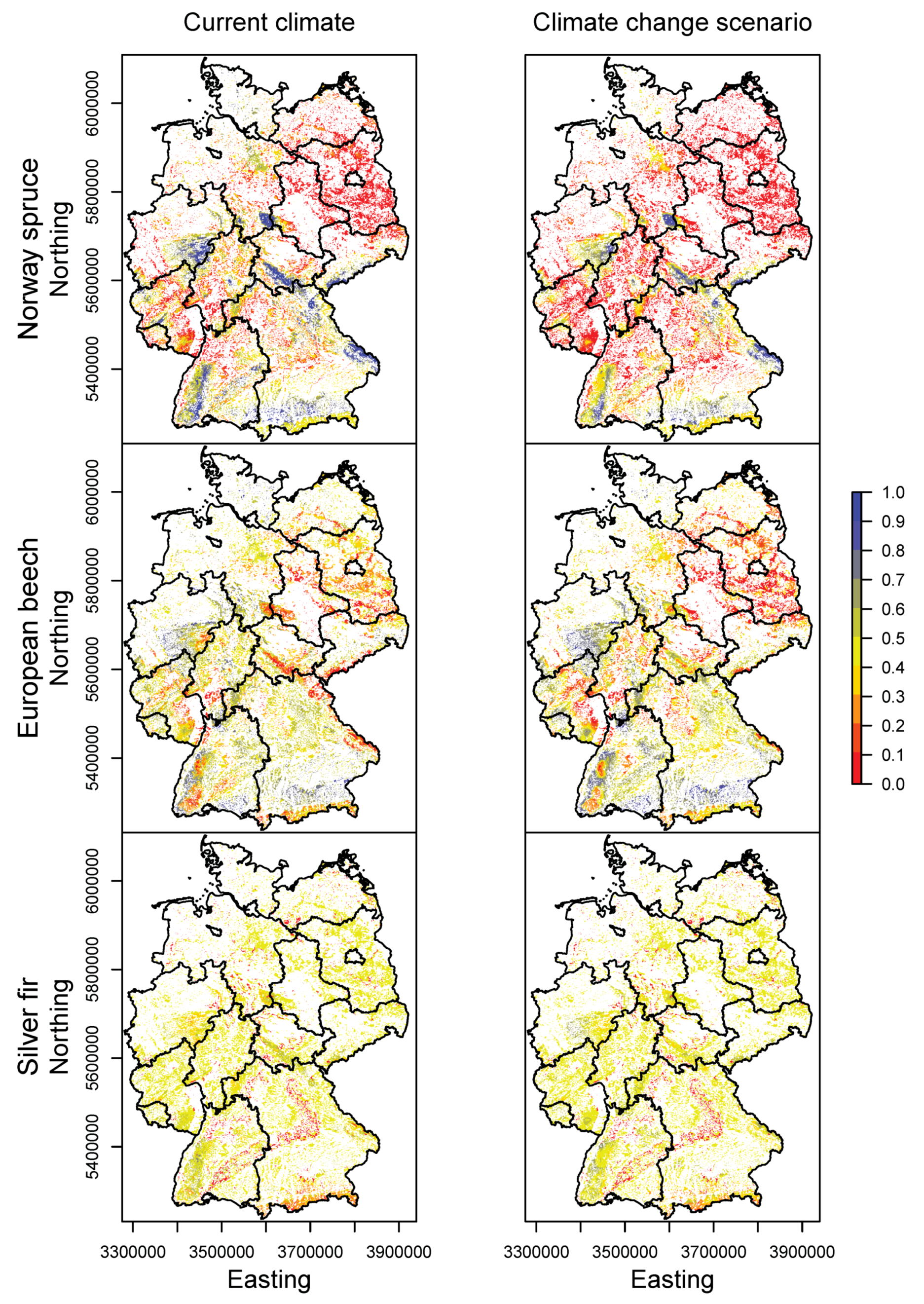
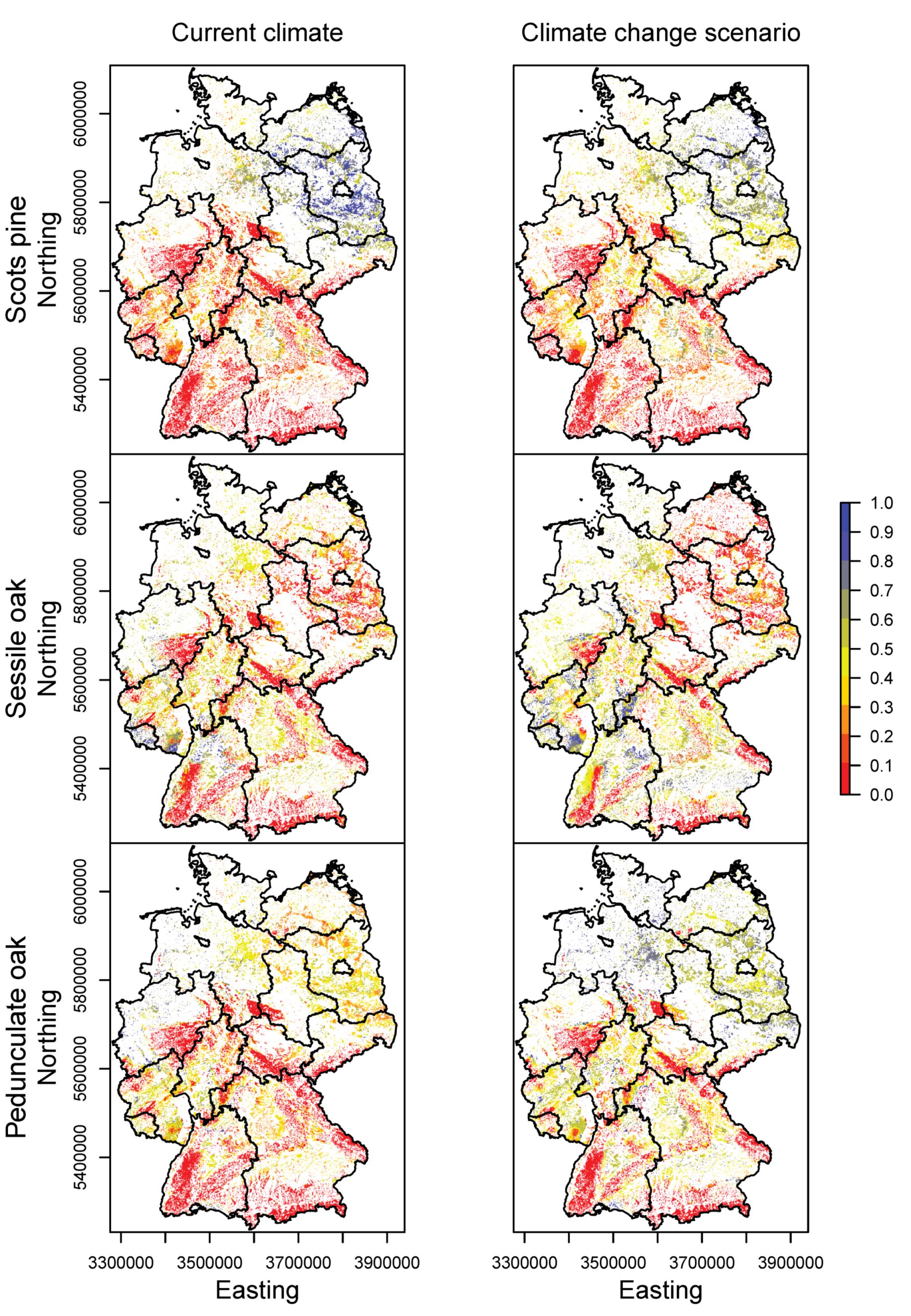
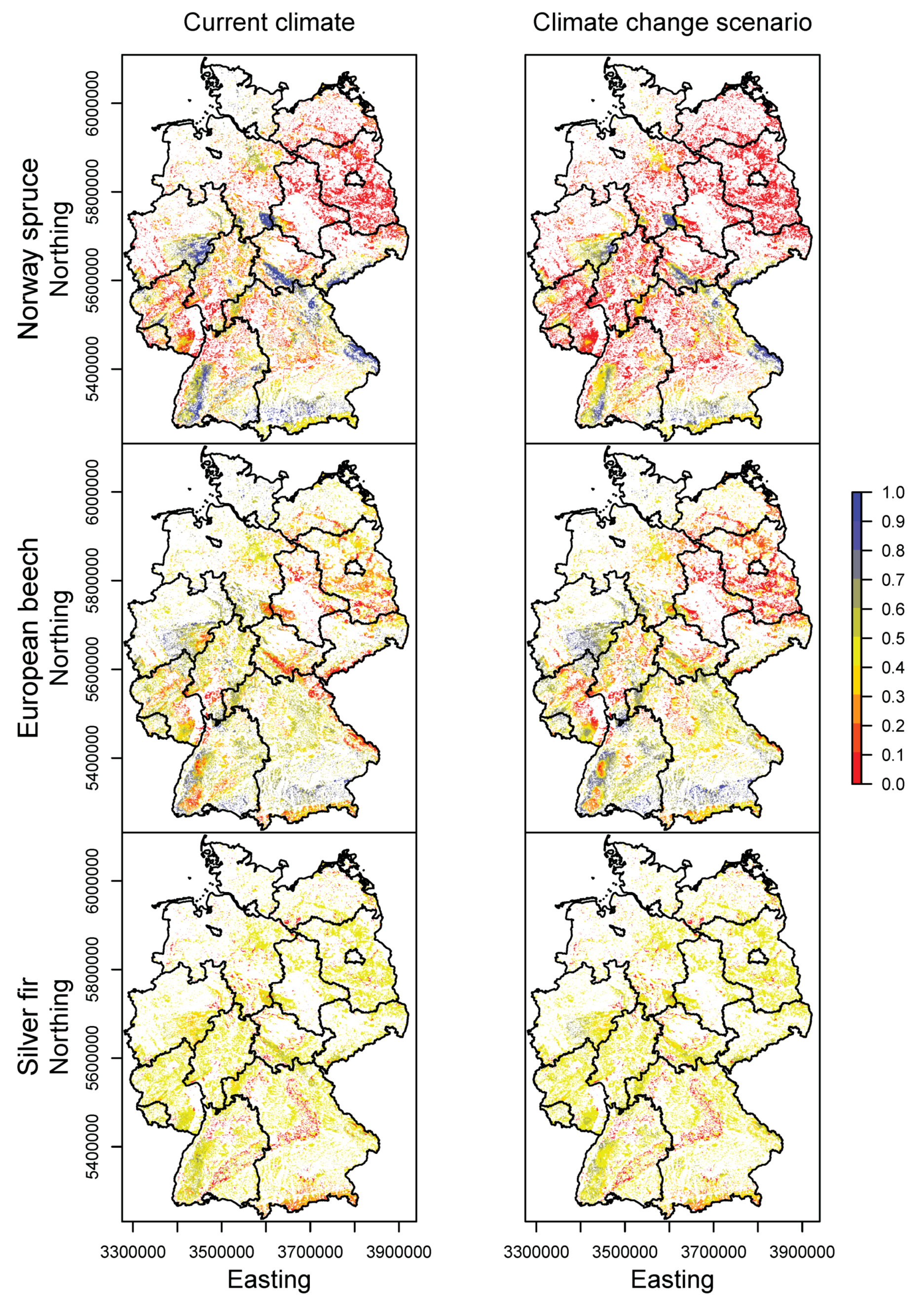
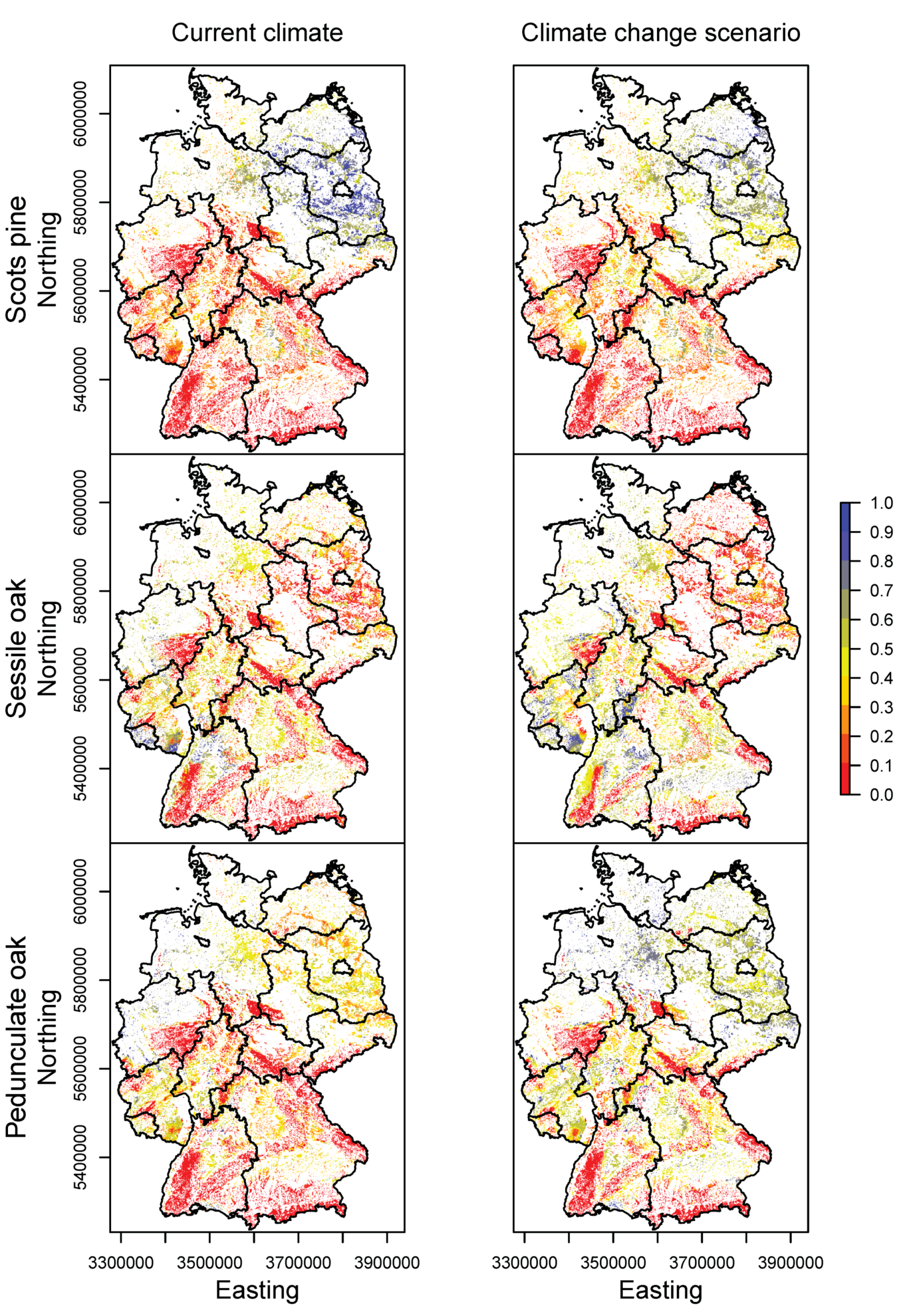
3.2. Happy Tree Index and Climate Change Projections
3.3. Current Stands under Climate Change
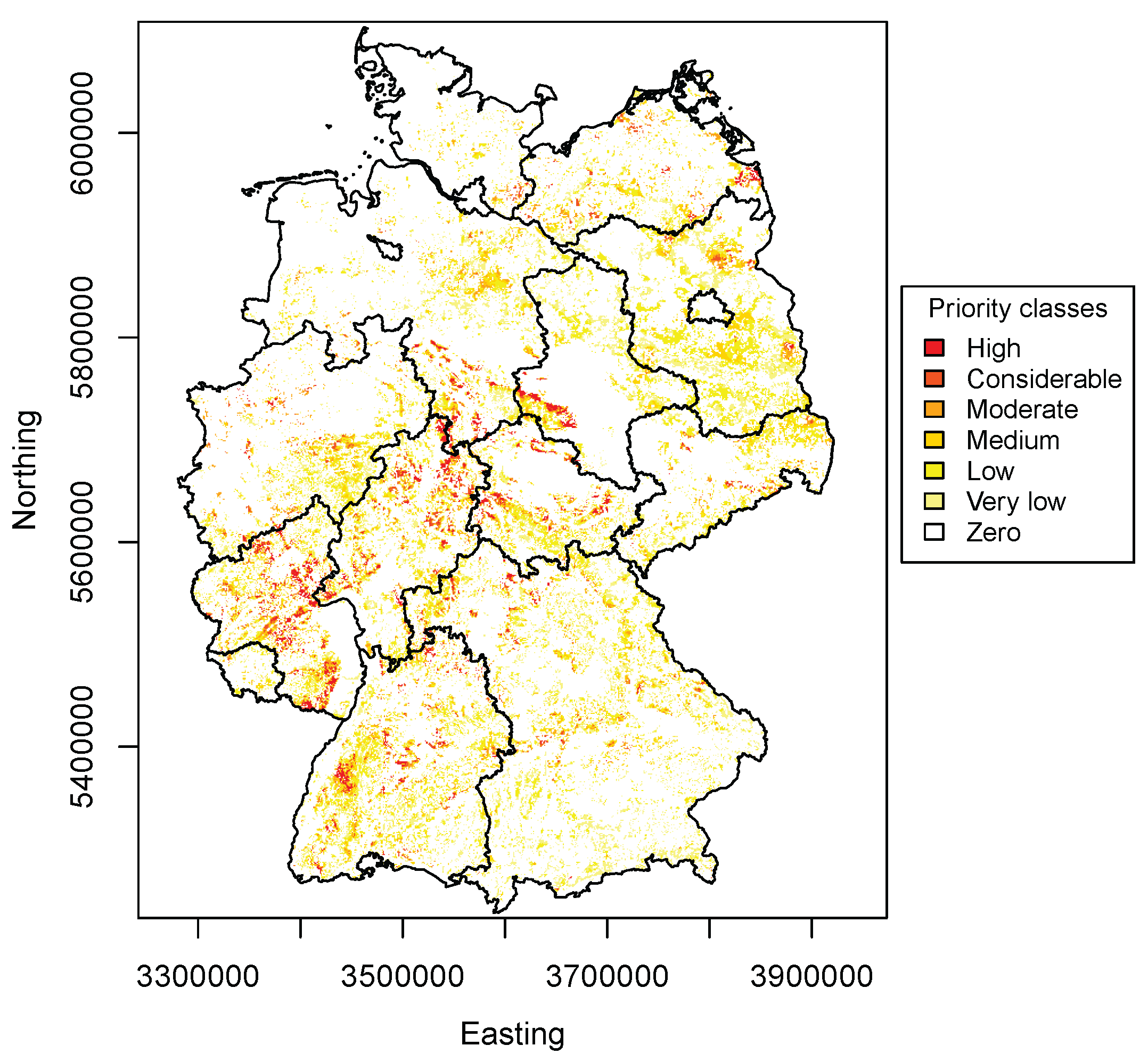
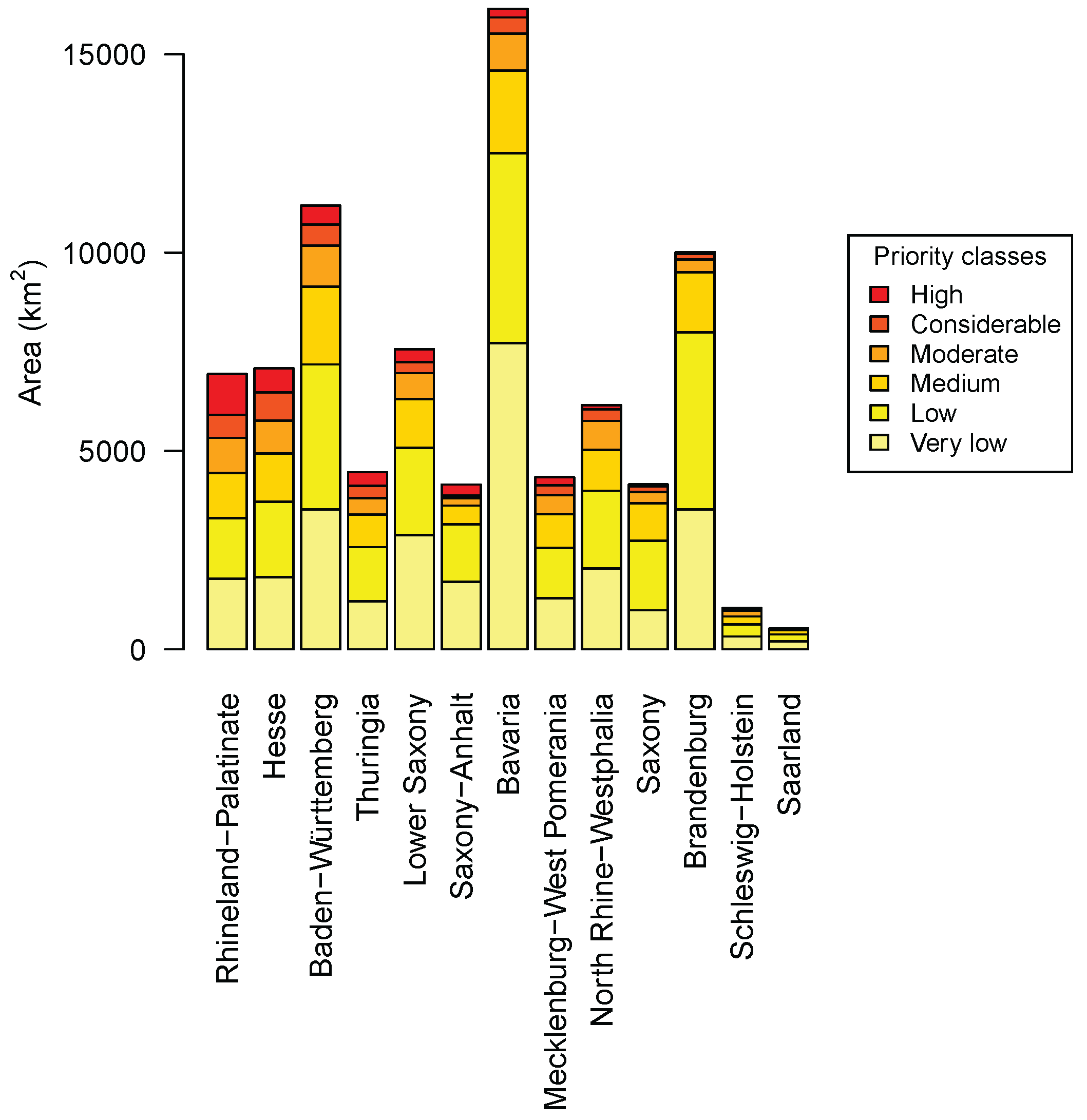
3.4. Priority Regions
4. Discussion
4.1. Current Site Suitability
4.2. Winners and Losers under Climate Change
4.3. Priority Regions
4.4. Validity of HTI and Priority Regions
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Distribution Model | |||||||
|---|---|---|---|---|---|---|---|
| Species | Data points Presence/Absence (Prevalence) | Temperature (C) Mean (min–max) | Precipitation (mm) Mean (min–max) | Soil Type (Most Frequent) | |||
| Norway spruce (Picea abies) | 19450/29327 (0.40) | 7.9 (2.6–10.8) | 1008 (499–2696) | Cambisol | |||
| European beech (Fagus sylvatica) | 16940/31837 (0.35) | 8.4 (2.6–11.0) | 908 (454–2503) | Cambisol | |||
| Scots pine (Pinus sylvestris) | 14220/34557 (0.29) | 8.8 (3.5–10.9) | 733 (471–2300) | Cambisol | |||
| Sessile oak (Quercus petraea) | 5716/43061 (0.12) | 8.8 (6.8–10.9) | 819 (458–1811) | Cambisol | |||
| Pedunculate oak (Quercus robur) | 5291/43486 (0.11) | 8.9 (6.1–11.0) | 755 (454–1902) | Cambisol | |||
| Silver fir (Abies alba) | 3161/45616 (0.06) | 7.8 (2.6–10.8) | 1285 (590–2575) | Cambisol | |||
| Growth model | |||||||
| Species | Data Points | Temperature (C) Mean (min–max) | Precipitation (mm) Mean (min–max) | Soil Type (Most Frequent) | Relative Basal Area Increment Mean (min–max) | Diameter at Breast Height (mm) Mean (min–max) | Stand Basal Area (m2/ha) Mean (min–max) |
| Norway spruce | 134151 | 8.2 (2.9–11.5) | 1106 (509–2745) | Cambisol | 0.037 (0–0.976) | 320 (70–1231) | 43.8 (4.0–125.0) |
| European beech | 70235 | 8.9 (2.9–11.5) | 955 (535–2590) | Cambisol | 0.03 (0–0.707) | 366 (70–1587) | 35.4 (4.0–122.8) |
| Scots pine | 55622 | 9.3 (3.1–11.7) | 767 (513–2391) | Cambisol | 0.027 (0–1.037) | 304 (76–899) | 37 (4.0–108.3) |
| Sessile oak | 13947 | 9.5 (7.3–11.5) | 823 (456–1905) | Cambisol | 0.023 (0–0.483) | 362 (70–1100) | 34.4 (4.0–97.3) |
| Pedunculate oak | 10532 | 9.5 (7.1–11.6) | 777 (509–1890) | Cambisol | 0.026 (0–0.828) | 439 (70–1432) | 32.9 (4.0–110) |
| Silver fir | 14719 | 8.3 (2.9–11.3) | 1319 (618–2635) | Cambisol | 0.033 (0–0.665) | 420 (70–1277) | 43.9 (4.0–107.3) |
| Threshold | Description | Norway Spruce | European Beech | Silver Fir | Scots Pine | Sessile Oak | Pedunculate Oak |
|---|---|---|---|---|---|---|---|
| Species distribution model | |||||||
| tv1 | Value for which false positive rate <1% | 0.80 | 0.58 | 0.43 | 0.70 | 0.37 | 0.30 |
| tv11 | Value for which false positive rate <10% | 0.64 | 0.48 | 0.23 | 0.48 | 0.29 | 0.26 |
| tv2 | Value for which Kappa is maximum | 0.42 | 0.35 | 0.19 | 0.37 | 0.21 | 0.22 |
| tv31 | Value for which false negative rate <10% | 0.24 | 0.25 | 0.36 | 0.17 | 0.17 | 0.17 |
| tv3 | Value for which false negative rate <1% | 0.10 | 0.12 | 0.05 | 0.09 | 0.05 | 0.08 |
| Growth model | |||||||
| q1 | 1/6 quantile | 0.0319 | 0.0316 | 0.0298 | 0.0211 | 0.0210 | 0.0213 |
| q2 | 2/6 quantile | 0.0332 | 0.0328 | 0.0310 | 0.0230 | 0.0224 | 0.0222 |
| q3 | 3/6 quantile | 0.0342 | 0.0340 | 0.0323 | 0.0241 | 0.0233 | 0.0226 |
| q4 | 4/6 quantile | 0.0350 | 0.0354 | 0.0332 | 0.0253 | 0.0238 | 0.0231 |
| q5 | 5/6 quantile | 0.0360 | 0.0376 | 0.0341 | 0.0260 | 0.0245 | 0.0239 |
References
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M. Predicting Abundance of 80 Tree Species Following Climate Change in the Eastern United States. Ecol. Monogr. 1998, 68, 465–485. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Svenning, J.C.; Skov, F. Limited filling of the potential range in European tree species. Ecol. Lett. 2004, 7, 565–573. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Morin, X.; Thuiller, W. Comparing niche- and process-based models to reduce prediction uncertainty in species range shifts under climate change. Ecology 2009, 90, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Purves, D.W. The demography of range boundaries versus range cores in eastern US tree species. Proc. R. Soc. B Biol. Sci. 2009, 276, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Benito-Garzon, M.; Ruiz-Benito, P.; Zavala, M.A. Interspecific differences in tree growth and mortality responses to environmental drivers determine potential species distributional limits in Iberian forests. Glob. Ecol. Biogeogr. 2013, 22, 1141–1151. [Google Scholar] [CrossRef]
- Peng, C. From static biogeographical model to dynamic global vegetation model: A global perspective on modelling vegetation dynamics. Ecol. Model. 2000, 135, 33–54. [Google Scholar] [CrossRef]
- Albert, M.; Schmidt, M. Climate-sensitive modelling of site-productivity relationships for Norway spruce (Picea abies (L.) Karst.) and common beech (Fagus sylvatica L.). For. Ecol. Manag. 2010, 259, 739–749. [Google Scholar] [CrossRef]
- Charru, M.; Seynave, I.; Morneau, F.; Bontemps, J.D. Recent changes in forest productivity: An analysis of national forest inventory data for common beech (Fagus sylvatica L.) in north-eastern France. For. Ecol. Manag. 2010, 260, 864–874. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; García-Valdés, R.; Ruíz-Benito, P.; Zavala, M.A. Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: Implications for forest management under global change. Glob. Chang. Biol. 2011, 17, 2400–2414. [Google Scholar] [CrossRef]
- Nothdurft, A.; Wolf, T.; Ringeler, A.; Böhner, J.; Saborowski, J. Spatio-temporal prediction of site index based on forest inventories and climate change scenarios. For. Ecol. Manag. 2012, 279, 97–111. [Google Scholar] [CrossRef]
- Brandl, S.; Falk, W.; Klemmt, H.J.; Stricker, G.; Bender, A.; Rötzer, T.; Pretzsch, H. Possibilities and Limitations of Spatially Explicit Site Index Modelling for Spruce Based on National Forest Inventory Data and Digital Maps of Soil and Climate in Bavaria (SE Germany). Forests 2014, 5, 2626–2646. [Google Scholar] [CrossRef]
- Kölling, C. Was wächst künftig wo? Klima-Risikokarten als Planungsunterlagen für den Waldumbau. Bayer. Landwirtsch. Wochenbl. 2009, 48, 58–60. [Google Scholar]
- Falk, W.; Mellert, K.H. Species distribution models as a tool for forest management planning under climate change: risk evaluation of Abies alba in Bavaria. J. Veg. Sci. 2011, 22, 621–634. [Google Scholar] [CrossRef]
- Falk, W.; Mellert, K.H.; Bachmann-Gigl, U.; Kölling, C. Bäume für die Zukunft: Baumartenwahl auf wissenschaftlicher Grundlage. Anbaurisikokarten jetzt um Boden- und Reliefparameter ergänzt. LWF Aktuell 2013, 94, 8–11. [Google Scholar]
- Hanewinkel, M.; Cullmann, D.A.; Michiels, H.G.; Kändler, G. Converting probabilistic tree species range shift projections into meaningful classes for management. J. Environ. Manag. 2014, 134, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Polley, H.; Schmitz, F.; Hennig, P.; Kroiher, F. Germany. In National Forest Inventories—Pathways for Common Reporting; Tomppo, E., Gschwantner, T., Lawrence, M., McRoberts, R.E., Eds.; Springer: New York, NY, USA, 2010. [Google Scholar]
- Albrecht, A.T.; Fortin, M.; Kohnle, U.; Ningre, F. Coupling a tree growth model with storm damage modeling—Conceptual approach and results of scenario simulations. Environ. Model. Softw. 2015, 69, 63–76. [Google Scholar] [CrossRef]
- Thuiller, W.; Münkemüller, T.; Schiffers, K.H.; Georges, D.; Dullinger, S.; Eckhart, V.M.; Edwards, T.C.; Gravel, D.; Kunstler, G.; Merow, C.; et al. Does probability of occurrence relate to population dynamics? Ecography 2014, 37, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Dolos, K.; Bauer, A.; Albrecht, S. Site suitability for tree species: Is there a positive relation between a tree species’ occurrence and its growth? Eur. J. For. Res. 2015, 134, 609–621. [Google Scholar] [CrossRef]
- Tinner, W.; Lotter, A.F. Holocene expansions of Fagus silvatica and Abies alba in Central Europe: Where are we after eight decades of debate? Quat. Sci. Rev. 2006, 25, 526–549. [Google Scholar] [CrossRef]
- Mauri, A.; de Rigo, D.; Caudullo, G. Abies alba in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Hansen, J.K.; Larsen, J.B. European silver fir (Abies alba Mill.) provenances from Calabria, southern Italy: 15-year results from Danish provenance field trials. Eur. J. For. Res. 2004, 123, 127–138. [Google Scholar] [CrossRef]
- Wykoff, W. A Basal Area Increment Model for Individual Conifers in the Northern Rocky-Mountains. For. Sci. 1990, 36, 1077–1104. [Google Scholar]
- Hijmans, R.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling; R Package; 2016. [Google Scholar]
- Babst, F.; Carrer, M.; Poulter, B.; Urbinati, C.; Neuwirth, B.; Frank, D. 500 years of regional forest growth variability and links to climatic extreme events in Europe. Environ. Res. Lett. 2012, 7, 045705. [Google Scholar] [CrossRef]
- Moss, R.; Babiker, W.; Brinkman, S.; Calvo, E.; Carter, T.; Edmonds, J.; Elgizouli, I.; Emori, S.; Erda, L.; Hibbard, K.; et al. Towards New Scenarios for the Analysis of Emissions: Climate Change, Impacts and Response Strategies; Intergovernmental Panel on Climate Change Secretariat (IPCC): Geneva, Switzerland, 2008. [Google Scholar]
- Sedlmeier, K.; Schädler, G. Ensembles Hoch Aufgelöster Regionaler Klimasimulationen zur Analyse Regionaler Klimaänderungen in Baden-Württemberg und Ihre Auswirkungen; LUBW Landesanstalt für Umwelt, Messungen und Naturschutz Baden-Württemberg: Karlsruhe, Germany, 2015. [Google Scholar]
- Sedlmeier, K.; Mieruch, S.; Schädler, G.; Kottmeier, C. Compound extremes in a changing climate—A Markov Chain approach. Nonlin. Processes Geophys. 2016, 23, 375–390. [Google Scholar] [CrossRef]
- Panagos, P.; Van Liedekerke, M.; Jones, A.; Montanarella, L. European Soil Data Centre: Response to European policy support and public data requirements. Land Use Policy 2012, 29, 329–338. [Google Scholar] [CrossRef]
- Wood, S.N. Thin plate regression splines. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2003, 65, 95–114. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 6, 716–723. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2016. [Google Scholar]
- Wood, S.N. Stable and efficient multiple smoothing parameter estimation for generalized additive models. J. Am. Stat. Assoc. 2004, 99, 673–686. [Google Scholar] [CrossRef]
- Hijmans, R.J.; van Etten, J. Raster: Geographic Analysis and Modeling With Raster Data; R Package Version 2.0-12; 2012. [Google Scholar]
- Heagerty, P.J.; Zeger, S.L. Marginalized multilevel models and likelihood inference. Stat. Sci. 2000, 15, 1–19. [Google Scholar]
- European Environment Agency. Corine Land Cover 2012; European Environment Agency: Copenhagen, Denmark, 2012. [Google Scholar]
- Polley, H.; Hennig, P.; Kroiher, F.; Marks, A.; Riedel, T.; Schmidt, U.; Schwitzgebel, F.; Stauber, T. Der Wald in Deutschland - Ausgewählte Ergebnisse der Dritten Bundeswaldinventur; Bundesministerium für Ernährung und Landwirtschaft (BMEL): Berlin, Germany, 2016. [Google Scholar]
- Kohnle, U.; Hein, S.; Michiels, H.G. Waldbauliche Handlungsmöglichkeiten angesichts des Klimawandels. FVA-Einblick+ 2008, 1, 53–55. [Google Scholar]
- Gärtner, S.; Reif, A.; Xystrakis, F.; Sayer, U.; Bendagha, N.; Matzarakis, A. The drought tolerance limit of Fagus sylvatica forest on limestone in southwestern Germany. J. Veg. Sci. 2008, 19, 757–768. [Google Scholar] [CrossRef]
- Destatis. Land-und Forstwirtschaft, Fischerei-Forstwirtschaftliche Bodennutzung-Holzeinschlagsstatistik-Fachserie 3 Reihe 3.3.1; Destatis: Wiesbaden, Germany, 2016. [Google Scholar]
- Cheddadi, R.; Birks, H.J.B.; Tarroso, P.; Liepelt, S.; Gömöry, D.; Dullinger, S.; Meier, E.S.; Hülber, K.; Maiorano, L.; Laborde, H. Revisiting tree-migration rates: Abies alba (Mill.), a case study. Veg. Hist. Archaeobot. 2014, 23, 113–122. [Google Scholar] [CrossRef]
- Carcaillet, C.; Muller, S. Holocene tree-limit and distribution of Abies alba in the inner French Alps: Anthropogenic or climatic changes? Boreas 2005, 34, 468–476. [Google Scholar] [CrossRef]
- Muck, P.; Borchert, H.; Elling, W.; Hahn, J.; Immler, T.; Konnert, M.; Walentowski, H.; Walter, A. Die Weißtanne—Ein Baum mit Zukunft—Die Weißtanne ist ein Hoffnungsträger für den Waldbau im Klimawandel. LWF Aktuell 2008, 67, 56–58. [Google Scholar]
- Walentowski, H.; Kölling, C. Die Waldkiefer – bereit für den Klimawandel? LWF Wissen 2007, 57, 37–46. [Google Scholar]
- Gärtner, S.; Nill, M.; Prinz, J.; Essmann, H.; Reif, A. Transparenz in der Landschaftsplanung—Partizipation und Verwendung eines Entscheidungsunterstützungssystems am Beispiel xerothermer Lebensräume der “Trockenaue” am Südlichen Oberrhein. Nat. Landsch. 2008, 48, 229–238. [Google Scholar]
- Reif, A.; Brucker, U.; Kratzer, R.; Schmiedinger, A.; Bauhus, J. Waldbau und Baumartenwahl in Zeiten des Klimawandels aus Sicht des Naturschutzes—Abschlussbericht eines F+E-Vorhabens im Auftrag des Bundesamtes für Naturschutz, BfN-Skripten 272; FKZ 3508 84 0200; Bundesamt für Naturschutz: Bonn, Germany, 2010. [Google Scholar]
- Reif, A.; Gärtner, S. Die natürliche Verjüngung der laubabwerfenden Eichenarten Stieleiche (Quercus robur L.) und Traubeneiche (Quercus petraea Liebl.) – eine Literaturstudie mit besonderer Berücksichtigung der Waldweide. Waldökol. Online 2007, 5, 79–116. [Google Scholar]
- Schütz, J.P. Die Prinzipien des naturnahen Waldbaus sind auch bei Klimawandel gültig (Essay)|The principles of ecoforestry are also valid in a changing climate (essay). Schweiz. Z. Forstwes. 2009, 160, 68–73. [Google Scholar] [CrossRef]
- Thomas, F.M.; Blank, R.; Hartmann, G. Abiotic and biotic factors and their interactions as causes of oak decline in Central Europe. For. Pathol. 2002, 32, 277–307. [Google Scholar] [CrossRef]
- Möges, M. Klima-Konzept für den Staatswald. LWF Aktuell 2007, 60, 42–44. [Google Scholar]
- Landesanstalt für Umwelt, Messungen und Naturschutz Baden-Württemberg (LUBW). Anpassungsstrategie an den Klimawandel – Fachgutachten für das Handlungsfeld Wald und Forstwirtschaft (U13-W04-N12); LUBW Landesanstalt für Umwelt, Messungen und Naturschutz Baden-Württemberg: Karlsruhe, Germany, 2013. [Google Scholar]
- Vasconcelos, A.; Matthes, U.; Konold, W. Auswirkungen des Klimawandels auf den Wald in Rheinland-Pfalz. In Schlussberichte des Landesprojekts Klima- und Landschaftswandel in Rheinland-Pfalz (KlimLandRP), Teil 4, Modul Wald: 333 S; Rheinland-Pfalz Kompetenzzentrum für Klimawandelfolgen: Mainz, Germany, 2013. [Google Scholar]
- Bayerisches Staatsministerium für Umwelt und Verbraucherschutz (StMUV). Klima-Report Bayern 2015 Klimawandel, Auswirkungen, Anpassungs- und Forschungsaktivitäten; StMUV: Munich, Germany, 2015. [Google Scholar]
- Hessisches Landesamt für Umwelt und Geologie. Klimawandel und seine Folgen in Hessen; Hessisches Landesamt für Umwelt und Geologie: Wiesbaden, Germany, 2007. [Google Scholar]
- Thüringer Ministerium für Landwirtschaft, Forsten, Umwelt und Naturschutz. IMPAKT—Integriertes Maßnahmenkonzeptnahmenprogramm zur Anpassung an die Folgen des Klimawandels im Freistaat Thüringen; Thüringer Ministerium für Landwirtschaft, Forsten, Umwelt und Naturschutz: Erfurt, Germany, 2013. [Google Scholar]
- Ministerium für Landwirtschaft, Umwelt und Verbraucherschutz Mecklenburg-Vorpommern (LU). Maßnahmenkonzept zur Anpassung der Wälder Mecklenburg-Vorpommerns an den Klimawandel; Ministerium für Landwirtschaft, Umwelt und Verbraucherschutz Mecklenburg-Vorpommern (LU): Schwerin, Germany, 2010. [Google Scholar]
- Ministerium für Infrastruktur und Landwirtschaft (MIL) des Landes Brandenburg. Wald im Klimawandel—Risiken und Anpassungsstrategien; Ministerium für Infrastruktur und Landwirtschaft (MIL) des Landes Brandenburg: Potsdam, Germany, 2009. [Google Scholar]
- Ministerium für Klimaschutz, Umwelt, Landwirtschaft, Natur- und Verbraucherschutz des Landes Nordrhein-Westfalen. Wald und Waldmanagement im Klimawandel Anpassungsstrategie für Nordrhein-Westfalen; Ministerium für Klimaschutz, Umwelt, Landwirtschaft, Natur- und Verbraucherschutz des Landes Nordrhein-Westfalen: Düsseldorf, Germany, 2015. [Google Scholar]
- Beese, F.; Aspelmeier, S. Klimafolgenforschung in Niedersachsen (KLIFF)—Abschlussbericht 2009 bis 2013; KLIFF: Göttingen, Germany, 2014. [Google Scholar]
- Ministerium für Landwirtschaft, Umwelt und ländliche Räume. Klimaschutzbericht 2009 Schleswig-Holstein; Ministerium für Landwirtschaft, Umwelt und ländliche Räume: Kiel, Germany, 2009. [Google Scholar]
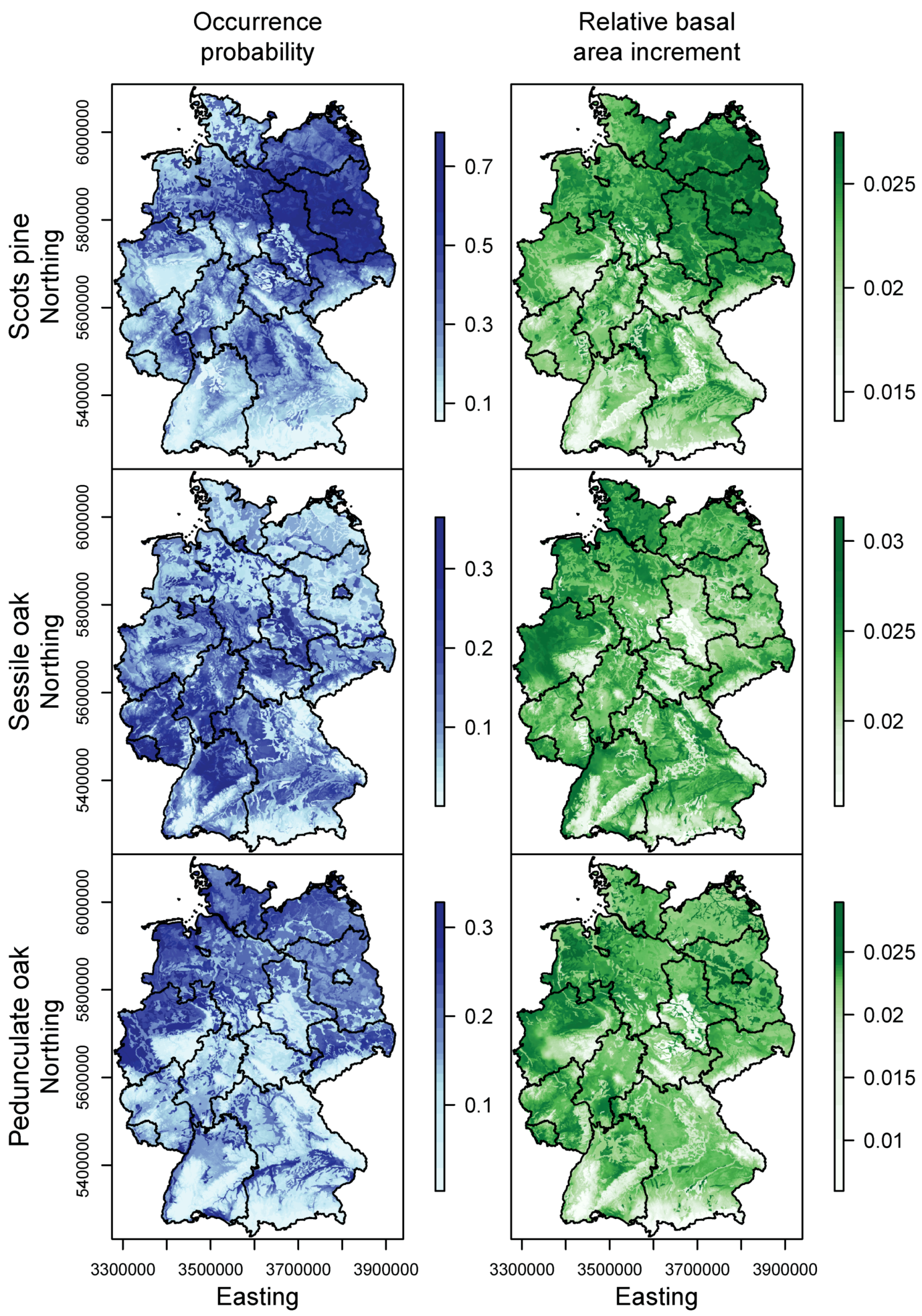
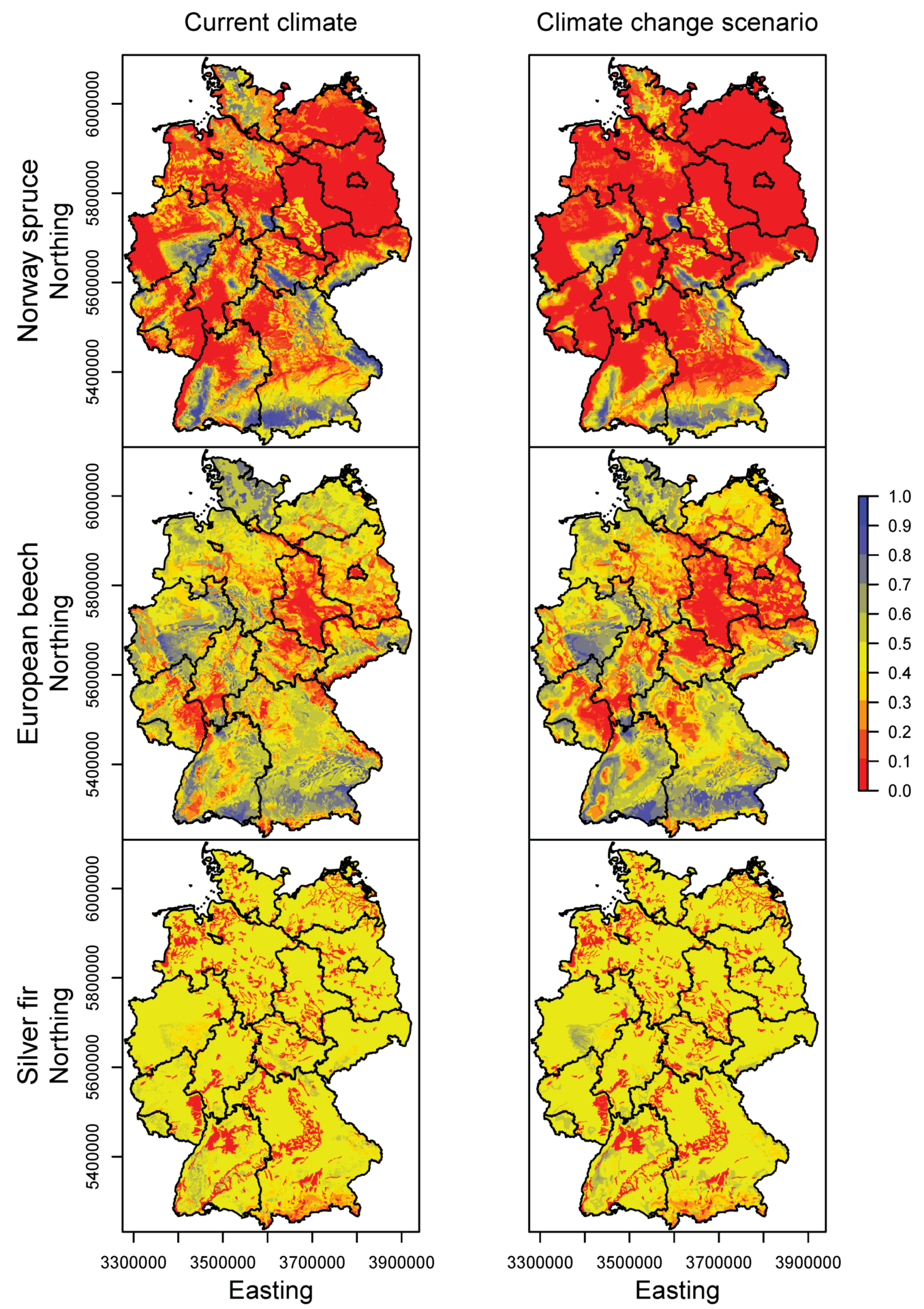
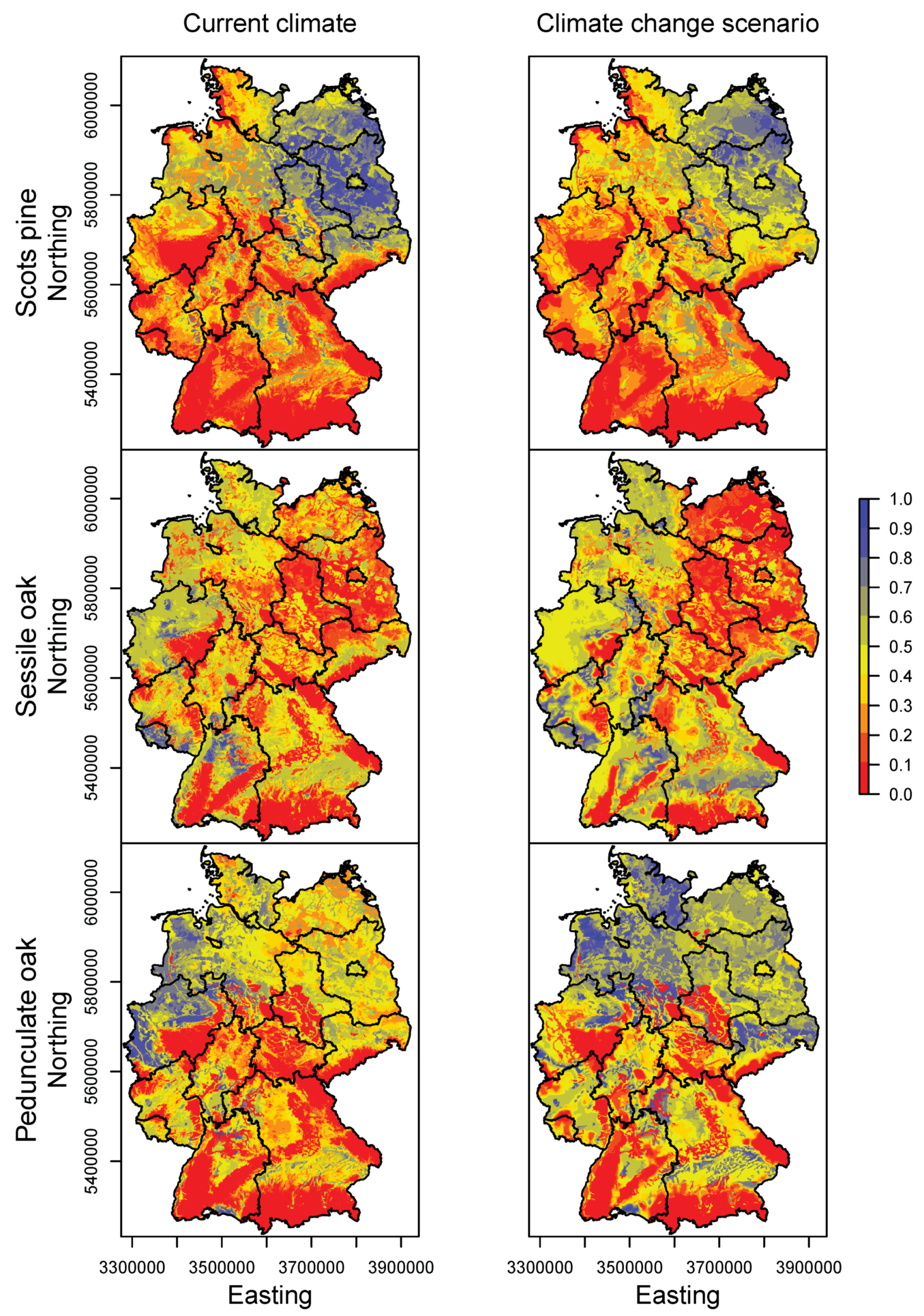
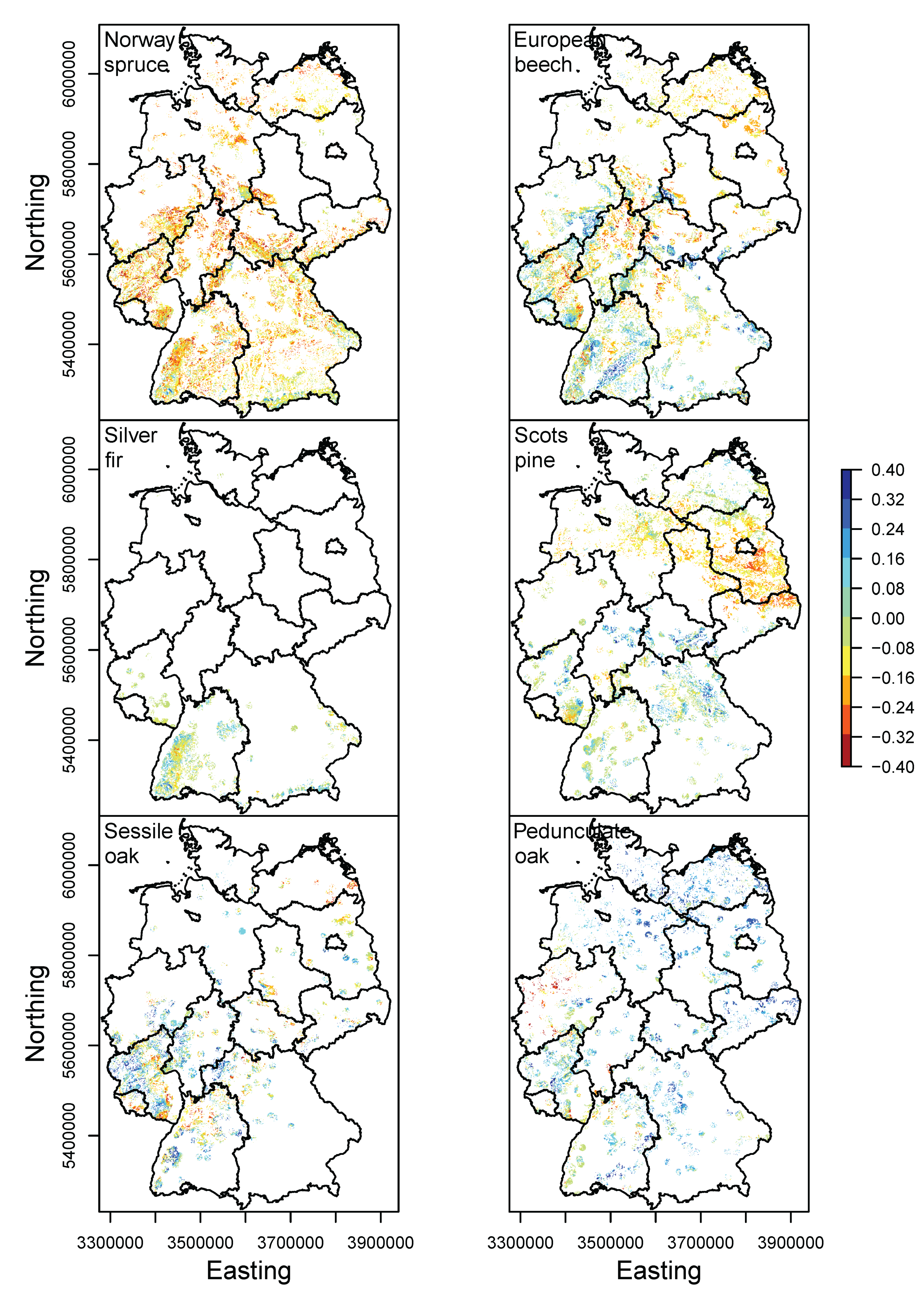
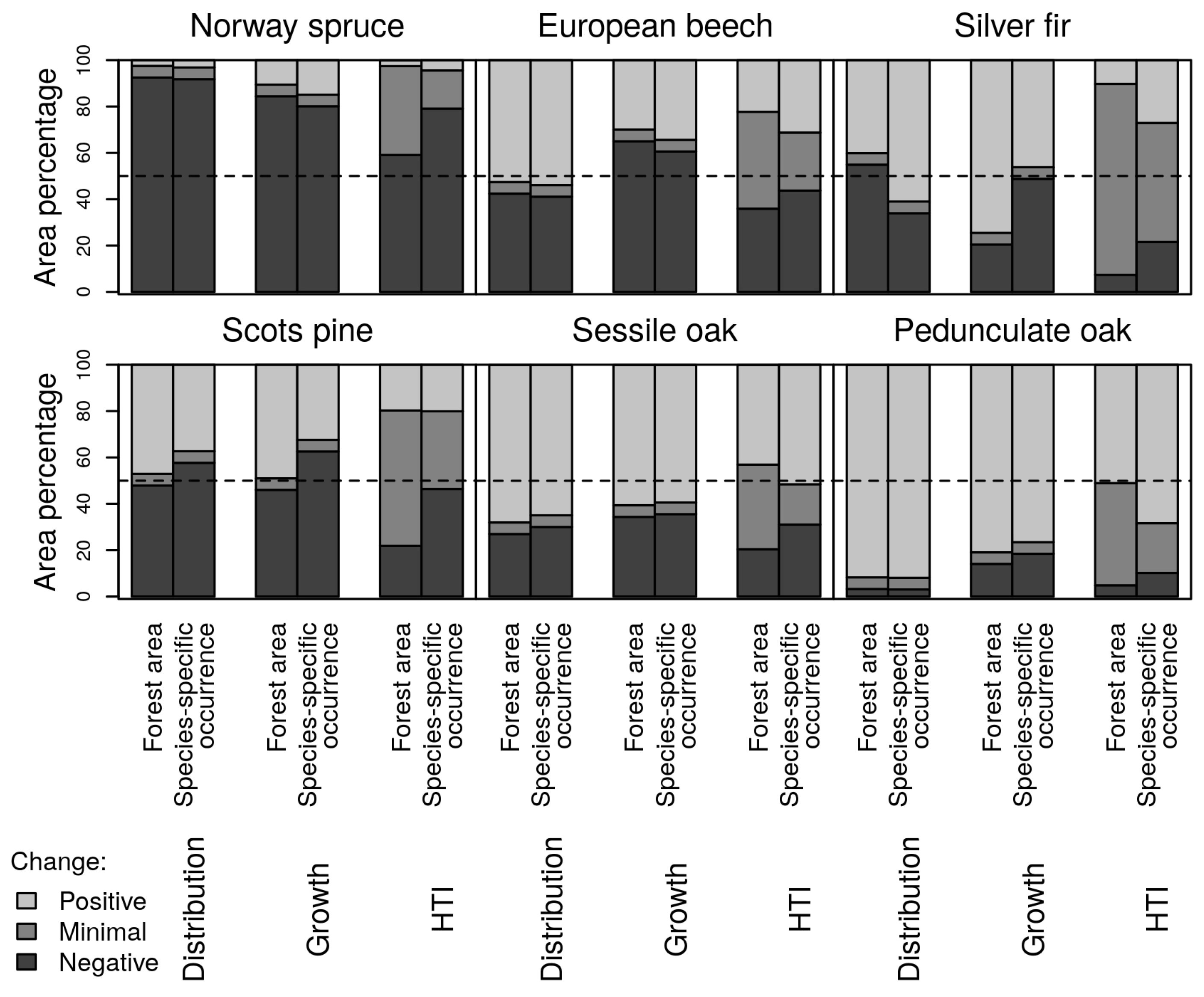

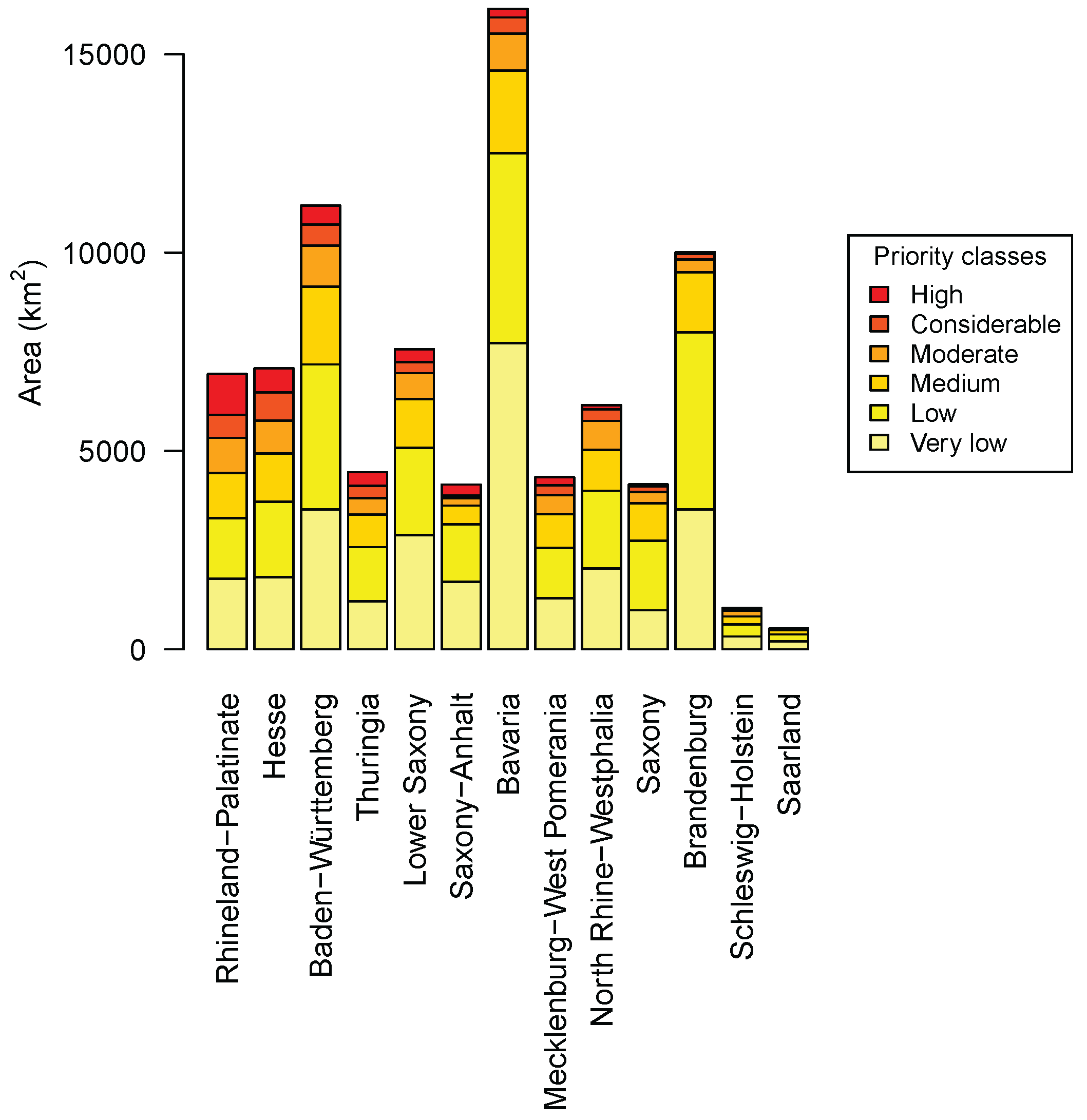
| Species | Response | Model | Formula | Explained Deviance | AIC |
|---|---|---|---|---|---|
| Norway spruce | Species distribution | Best model | Mean temperature of coldest quarter × Annual precipitation + Soil | 23.23% | 3724.66 |
| Standard model | Annual mean temperature × Annual precipitation + Soil | 23.17% | 3747.68 | ||
| Growth | Best model | Standard model | - | - | |
| Standard model | Annual mean temperature × Annual precipitation + Soil + DBH + SBA | 54.58% | −5282.82 | ||
| European beech | Species distribution | Best model | Standard model | - | - |
| Standard model | Annual mean temperature × Annual precipitation + Soil | 8.25% | 4378.61 | ||
| Growth | Best model | Standard model | - | - | |
| Standard model | Annual mean temperature × Annual precipitation + Soil + DBH + SBA | 46.90% | −5403.11 | ||
| Scots pine | Species distribution | Best model | Standard model | - | - |
| Standard model | Annual mean temperature × Annual precipitation + Soil | 19.70% | 3717.81 | ||
| Growth | Best model | Standard model | - | - | |
| Standard model | Annual mean temperature × Annual precipitation + Soil + DBH + SBA | 54.46% | −5063.36 | ||
| Sessile oak | Species distribution | Best model | Annual mean temperature + Precipitation of warmest quarter + Soil | 18.58% | 2370.84 |
| Standard model | Annual mean temperature × Annual precipitation + Soil | 17.59% | 2386.42 | ||
| Growth | Best model | Standard model | - | - | |
| Standard model | Annual mean temperature × Annual precipitation + Soil + DBH + SBA | 47.50% | −2089.45 | ||
| Pedunculate oak | Species distribution | Best model | Mean temperature of coldest quarter + Precipitation of driest month + Soil | 9.97% | 4214.26 |
| Standard model | Annual mean temperature × Annual precipitation + Soil | 9.59% | 4220.1 | ||
| Growth | Best model | Standard model | - | - | |
| Standard model | Annual mean temperature × Annual precipitation + Soil + DBH + SBA | 50.48% | −2339.1 | ||
| Silver fir | Species distribution | Best model | Standard model | - | - |
| Standard model | Annual mean temperature × Annual precipitation + Soil | 31.17% | 929.97 | ||
| Growth | Best model | Standard model | - | - | |
| Standard model | Annual mean temperature × Annual precipitation + Soil + DBH + SBA | 54.36% | −650.27 |
| Species | Species Distribution | Growth |
|---|---|---|
| Pearson’s r2 (CV) | ||
| Norway spruce | 0.25 (0.023) | 0.38 (0.005) |
| European beech | 0.07 (0.045) | 0.23 (0.004) |
| Scots pine | 0.21 (0.032) | 0.37 (0.007) |
| Sessile oak | 0.08 (0.044) | 0.25 (0.009) |
| Pedunculate oak | 0.05 (0.055) | 0.31 (0.013) |
| Silver fir | 0.18 (0.049) | 0.34 (0.011) |
| Federal State | Norway Spruce | European Beech | Silver Fir | Scots Pine | Sessile Oak | Pedunculate Oak |
|---|---|---|---|---|---|---|
| Germany | 0.79/0.16/0.04 | 0.44/0.25/0.31 | 0.22/0.51/0.27 | 0.46/0.34/0.2 | 0.31/0.17/0.52 | 0.1/0.22/0.68 |
| Baden-Württemberg | 0.79/0.14/0.07 | 0.26/0.3/0.44 | 0.23/0.47/0.3 | 0.12/0.61/0.27 | 0.36/0.17/0.47 | 0.14/0.34/0.52 |
| Bavaria | 0.72/0.22/0.06 | 0.35/0.29/0.37 | 0.21/0.47/0.32 | 0.12/0.46/0.42 | 0.25/0.15/0.6 | 0.03/0.2/0.77 |
| Brandenburg | 0.52/0.48/0 | 0.93/0.07/0 | - | 0.86/0.13/0.01 | 0.43/0.31/0.26 | 0.01/0.08/0.91 |
| Hesse | 0.87/0.1/0.03 | 0.56/0.22/0.22 | - | 0.28/0.27/0.45 | 0.2/0.16/0.64 | 0.17/0.16/0.67 |
| Lower Saxony | 0.86/0.12/0.03 | 0.54/0.29/0.18 | - | 0.68/0.28/0.04 | 0.11/0.14/0.76 | 0.03/0.16/0.81 |
| Mecklenburg-West Pomerania | 0.9/0.1/0 | 0.9/0.1/0 | - | 0.32/0.5/0.18 | 0.91/0.06/0.03 | 0/0.15/0.85 |
| North Rhine-Westphalia | 0.83/0.14/0.03 | 0.25/0.29/0.46 | - | 0.24/0.65/0.11 | 0.18/0.21/0.6 | 0.38/0.31/0.31 |
| Rhineland-Palatinate | 0.78/0.18/0.04 | 0.43/0.26/0.32 | 0.15/0.8/0.05 | 0.2/0.54/0.26 | 0.29/0.15/0.56 | 0.15/0.3/0.55 |
| Saarland | 0.81/0.13/0.06 | 0.28/0.22/0.5 | - | 0.92/0.07/0.01 | 0.49/0.1/0.41 | 0.38/0.05/0.57 |
| Saxony-Anhalt | 0.89/0.1/0.01 | 0.55/0.2/0.25 | - | 0.84/0.16/0 | 0.5/0.25/0.25 | 0.02/0.17/0.8 |
| Saxony | 0.75/0.23/0.02 | 0.4/0.21/0.4 | - | 0.75/0.12/0.13 | 0.29/0.21/0.5 | 0/0.18/0.82 |
| Schleswig-Holstein | 0.94/0.06/0 | 0.67/0.3/0.03 | 0/1/0 | 0.32/0.59/0.09 | 0.2/0.12/0.68 | 0/0.18/0.82 |
| Thuringia | 0.83/0.14/0.03 | 0.48/0.2/0.32 | - | 0.04/0.24/0.71 | 0.42/0.22/0.36 | 0.01/0.2/0.79 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Märkel, U.; Dolos, K. Tree Species Site Suitability as a Combination of Occurrence Probability and Growth and Derivation of Priority Regions for Climate Change Adaptation. Forests 2017, 8, 181. https://doi.org/10.3390/f8060181
Märkel U, Dolos K. Tree Species Site Suitability as a Combination of Occurrence Probability and Growth and Derivation of Priority Regions for Climate Change Adaptation. Forests. 2017; 8(6):181. https://doi.org/10.3390/f8060181
Chicago/Turabian StyleMärkel, Ulrike, and Klara Dolos. 2017. "Tree Species Site Suitability as a Combination of Occurrence Probability and Growth and Derivation of Priority Regions for Climate Change Adaptation" Forests 8, no. 6: 181. https://doi.org/10.3390/f8060181
APA StyleMärkel, U., & Dolos, K. (2017). Tree Species Site Suitability as a Combination of Occurrence Probability and Growth and Derivation of Priority Regions for Climate Change Adaptation. Forests, 8(6), 181. https://doi.org/10.3390/f8060181