
Optimized MaxEnt Modeling of Catalpa bungei Habitat for Sustainable Management Under Climate Change in China

Abstract
1. Introduction
2. Materials and Methods
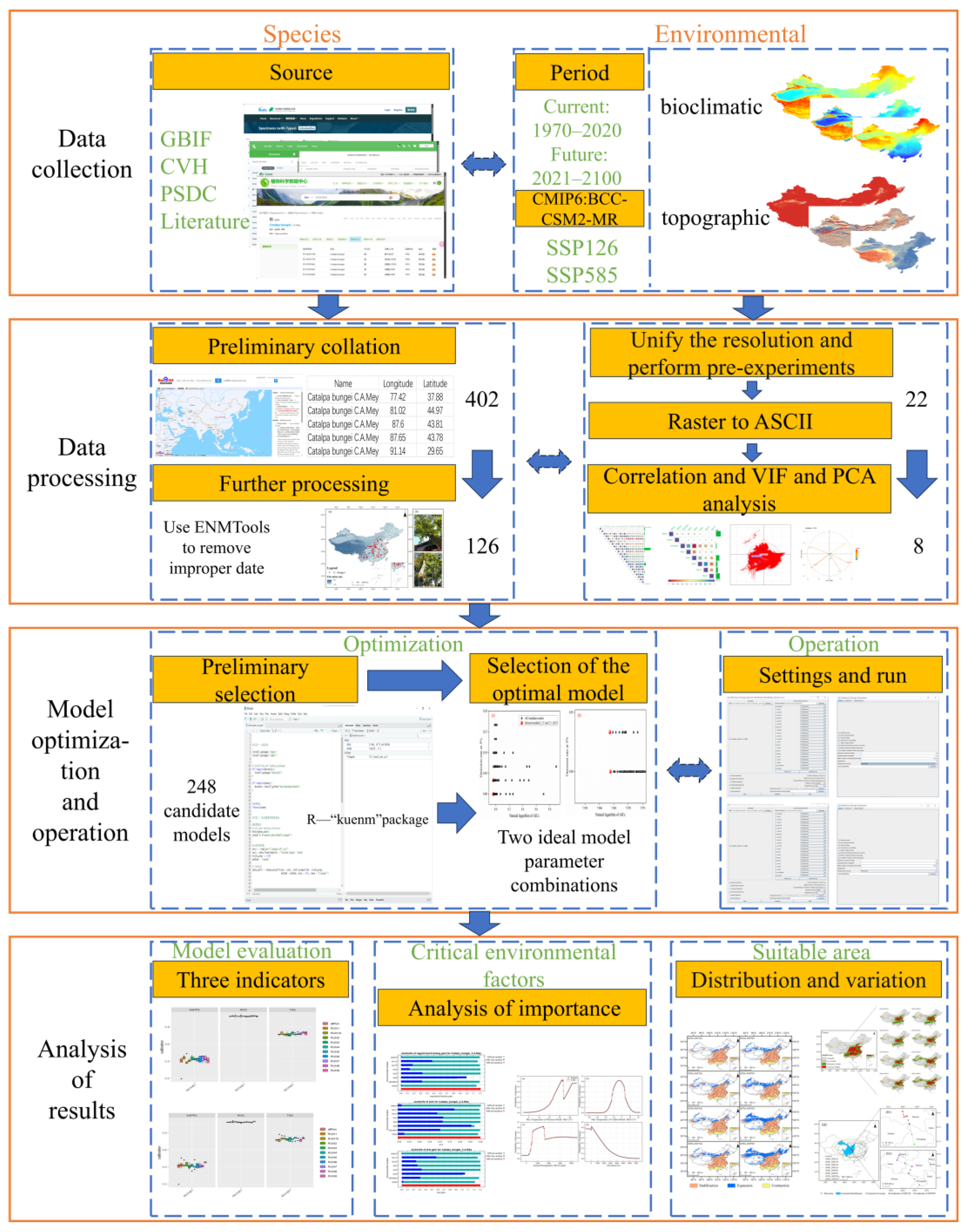
2.1. R-Based Integrated MaxEnt Modeling Framework
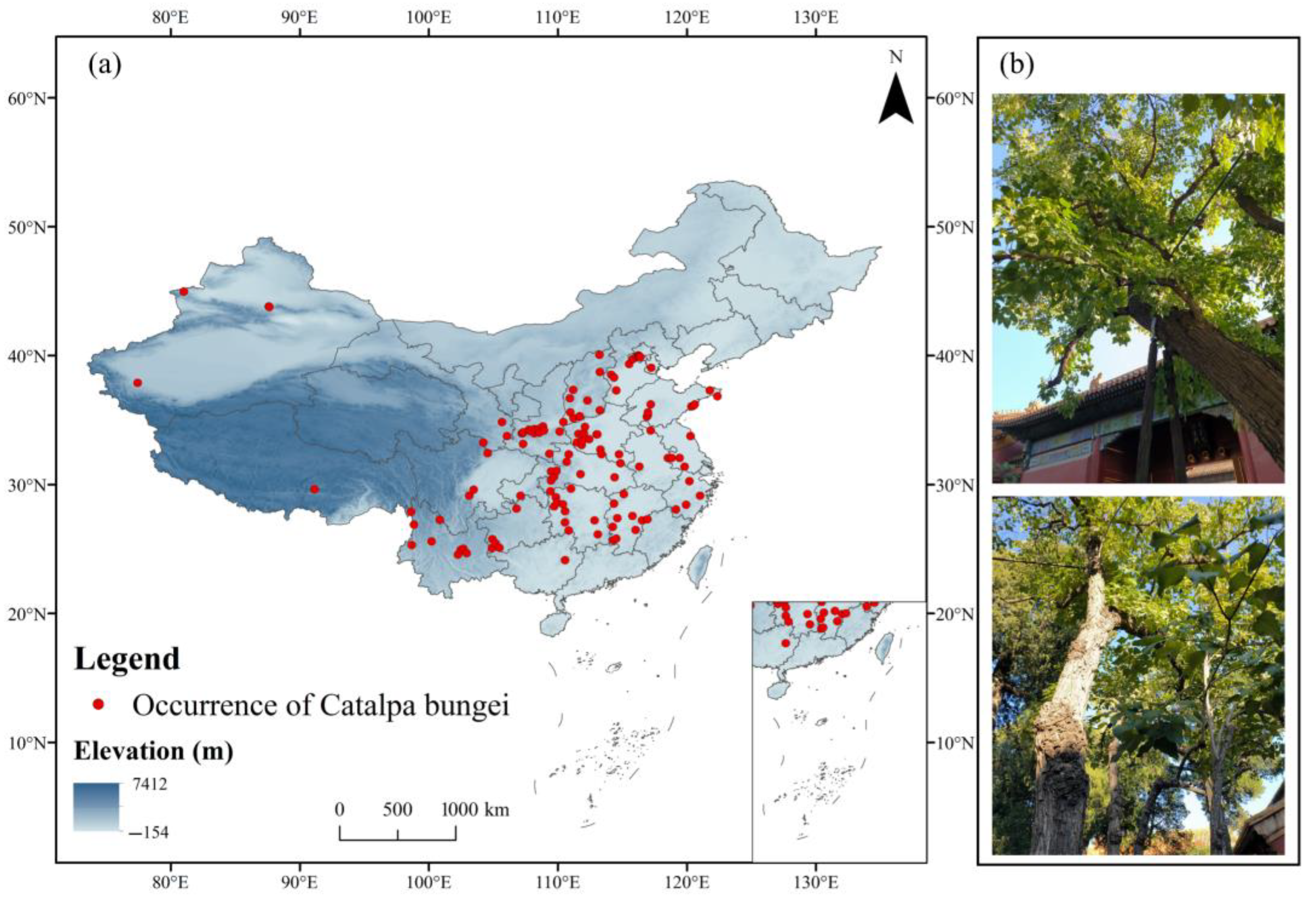
2.2. Compilation and Preprocessing of Species Occurrence Data
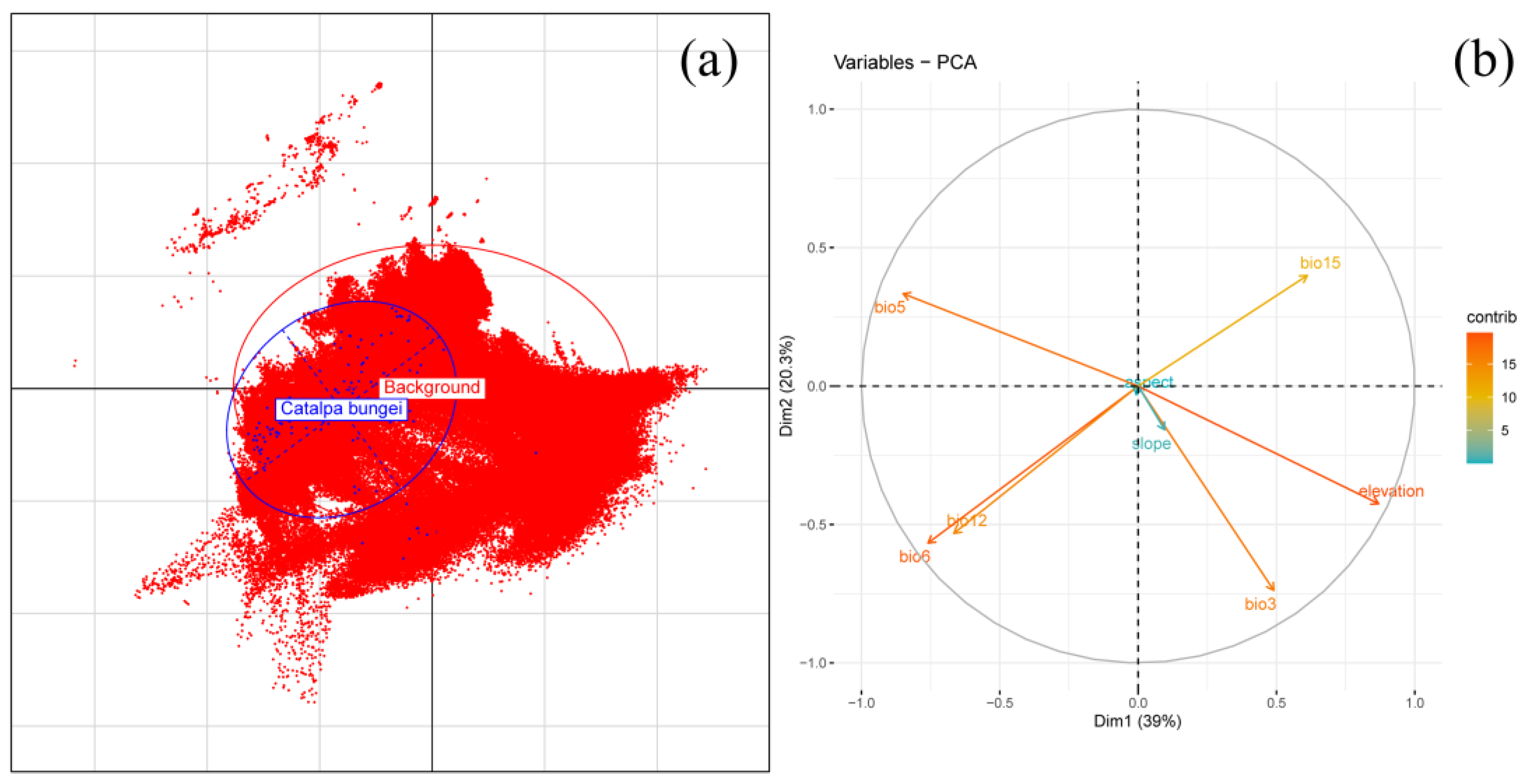
2.3. Environmental Variable Screening and Modeling Data Processing
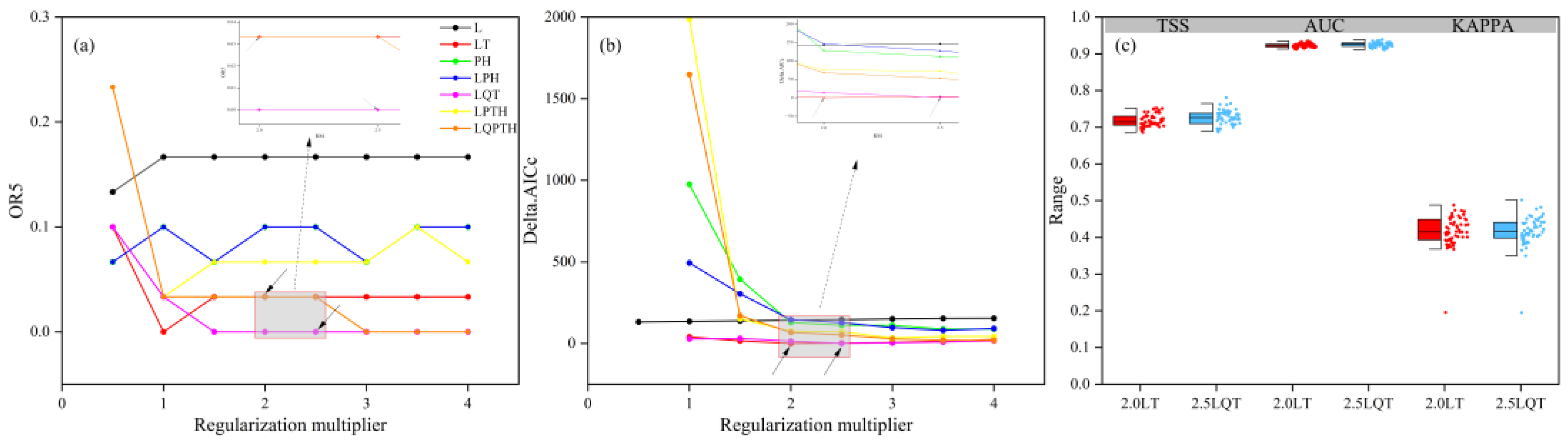
2.4. Parameter Optimization of MaxEnt Model Using the Kuenm Package
2.5. Model Operation and Evaluation
2.6. Classification of Suitable Areas
3. Results
3.1. Evaluation of MaxEnt Model Optimization and Prediction Results
3.2. The Current Distribution of C. bungei
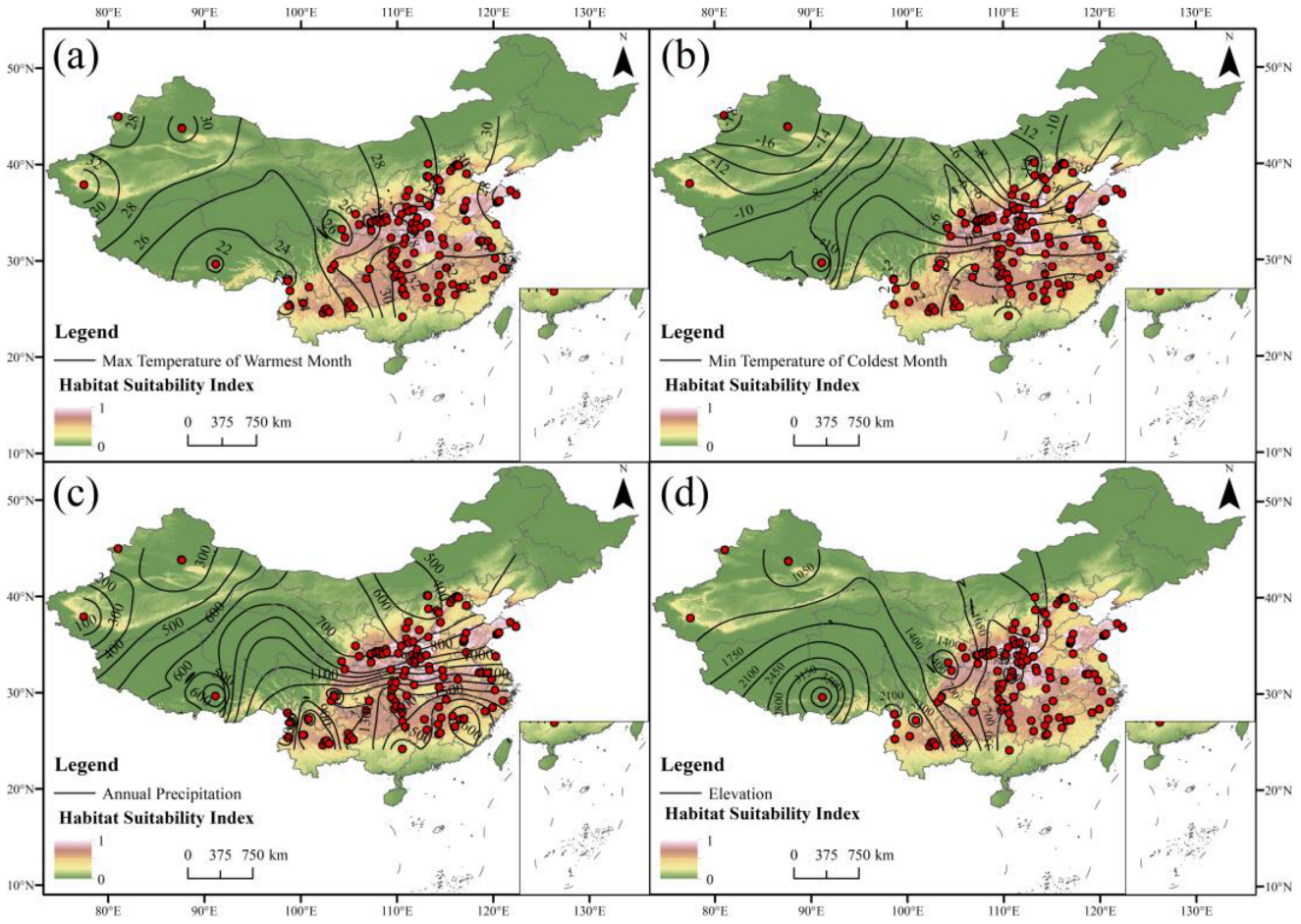
3.3. Critical Environmental Factors
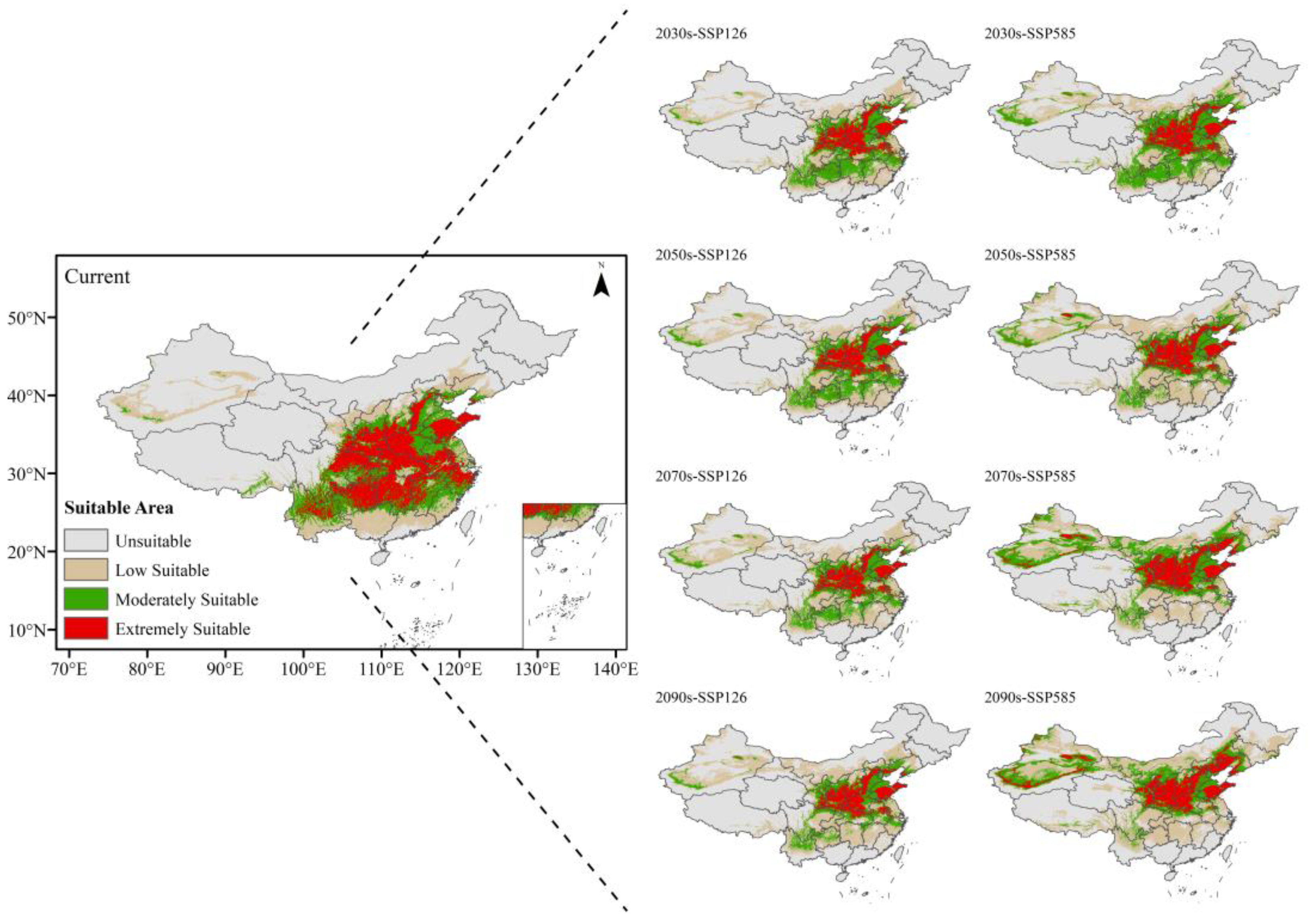
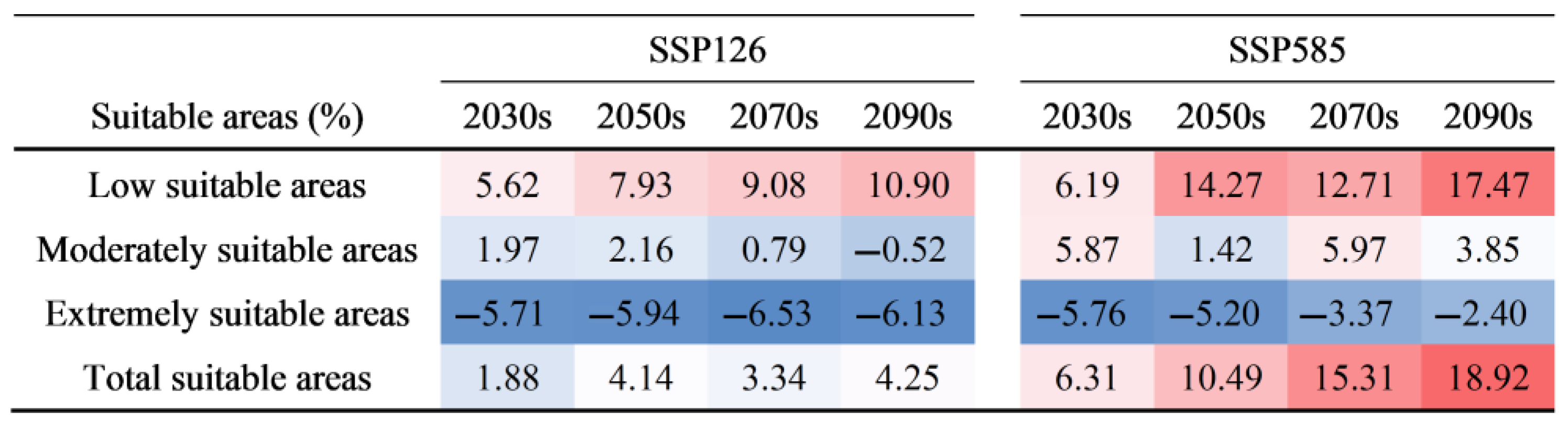
3.4. Future Potential Suitable Habitats for C. bungei
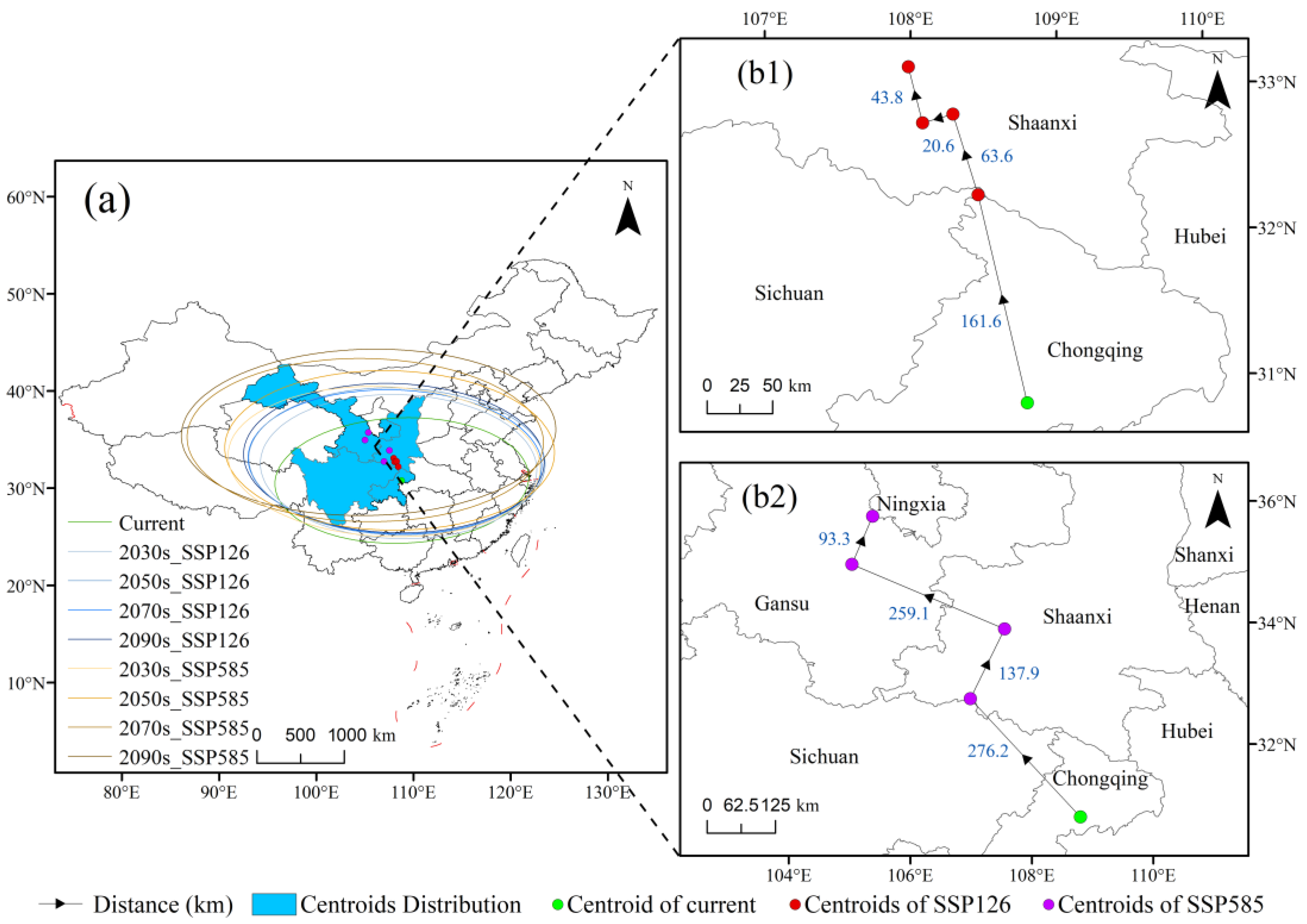
3.5. Centroid Migration of Suitable Areas Under Future Climate Scenarios
4. Discussion
4.1. Interpretation of Model Performance and Predictive Utility
4.2. Effects of Key Environmental Factors on the Distribution of C. bungei
4.3. Changes in and Management of C. bungei’s Suitable Habitats Under Future Climate Change
4.4. Limitations and Future Research Suggestions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Byers, B.A.; Cunliffe, R.N.; Hudak, A.T. Linking the conservation of culture and nature: A case study of sacred forests in Zimbabwe. Hum. Ecol. 2001, 29, 187–218. [Google Scholar] [CrossRef]
- Liu, L.; Qin, F.; Liu, Y.; Hu, Y.; Wang, W.; Duan, H.; Li, M. Forecast of potential suitable areas for forest resources in Inner Mongolia under the Shared Socioeconomic Pathway 245 scenario. Ecol. Indic. 2024, 167, 112694. [Google Scholar] [CrossRef]
- Luo, W.; Sun, C.; Yang, S.; Chen, W.; Sun, Y.; Li, Z.; Liu, J.; Tao, W.; Tao, J. Contrasting range changes and drivers of four forest foundation species under future climate change in China. Sci. Total Environ. 2024, 942, 173784. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yan, X.; Chang, Y.; Wang, Q.; Wang, Y.; Malik, A. Understanding the impact of interprovincial trade on forest resources in China. Renew. Sustain. Energy Rev. 2023, 186, 113673. [Google Scholar] [CrossRef]
- Pan, H.; Liu, G.; Muller, J.-P.; Sun, Z.; Yao, Y.; Chang, Y.; Xiong, Z.; Zhang, Y. Comprehensive Assessment of Sustainable Development of Terrestrial Ecosystem Based on SDG 15—A Case Study of Guilin City. Remote Sens. 2024, 17, 63. [Google Scholar] [CrossRef]
- Urban, M.C. Climate change extinctions. Science 2024, 386, 1123–1128. [Google Scholar] [CrossRef]
- Chen, H.; Sun, J. Anthropogenic influence has increased climate extreme occurrence over China. Sci. Bull. 2021, 66, 749–752. [Google Scholar] [CrossRef]
- Mukherji, A. Climate Change 2023 Synthesis Report; IPCC: Geneva, Switzerland, 2023. [Google Scholar]
- Liu, X.; Ma, K. Plant functional traits—Concepts, applications and future directions. Sci. Sin. Vitae 2015, 45, 325–339. [Google Scholar] [CrossRef]
- Chludil, D.; Čepl, J.; Steffenrem, A.; Stejskal, J.; Sagariya, C.; Pook, T.; Schueler, S.; Korecký, J.; Almqvist, C.; Chakraborty, D. A Pollen-Based Assisted Migration for Rapid Forest Adaptation. Glob. Change Biol. 2025, 31, e70014. [Google Scholar] [CrossRef]
- Wang, P.; Ma, L.; Li, Y.; Wang, S.A.; Li, L.; Yang, R.; Ma, Y.; Wang, Q. Transcriptome profiling of indole-3-butyric acid-induced adventitious root formation in softwood cuttings of the Catalpa bungei variety ‘YU-1’at different developmental stages. Genes Genom. 2016, 38, 145–162. [Google Scholar] [CrossRef]
- Wang, R.; Shi, L.; Wang, Y. Physical and mechanical properties of Catalpa bungei clones and estimation of the properties by near-infrared spectroscopy. J. Renew. Mater. 2022, 10, 3285. [Google Scholar] [CrossRef]
- Guo, P.; Zhao, X.; Yang, Z.; Wang, Y.; Li, H.; Zhang, L. Water, starch, and nuclear behavior in ray parenchyma during heartwood formation of Catalpa bungei ‘Jinsi’. Heliyon 2024, 10, e27231. [Google Scholar] [CrossRef] [PubMed]
- Mei, F.; Wang, J.-H.; Gu, J.-W.; Jiao, Y.-D.; Zhao, K. Genetic Diversity of Growth and Leaf Traits of Catalpa bungei. J. Plant Genet. Resour. 2014, 15, 207–211. [Google Scholar]
- Zheng, H.; Zhang, X.; Ma, W.; Song, J.; Rahman, S.U.; Wang, J.; Zhang, Y. Morphological and physiological responses to cyclic drought in two contrasting genotypes of Catalpa bungei. Environ. Exp. Bot. 2017, 138, 77–87. [Google Scholar] [CrossRef]
- Guan, Z.; Lin, D.; Chen, D.; Guo, Y.; Lu, Y.; Han, Q.; Li, N.; Su, Y.; Li, J.; Wang, J. Soil microbial communities response to different fertilization regimes in young Catalpa bungei plantation. Front. Microbiol. 2022, 13, 948875. [Google Scholar] [CrossRef]
- Zhou, X.; Hu, C.; Wang, Z. Distribution of biomass and carbon content in estimation of carbon density for typical forests. Glob. Ecol. Conserv. 2023, 48, e02707. [Google Scholar] [CrossRef]
- Yang, H.; Long, H.; Li, X.; Luo, X.; Liao, Y.; Wang, C.; Cai, H.; Shu, Y. Vegetation restoration improved aggregation stability and aggregated-associated carbon preservation in the karst areas of Guizhou Province, southwest China. PeerJ 2024, 12, e16699. [Google Scholar] [CrossRef]
- Xu, H.; Zhou, Z.; Dong, J.; Lei, M. Suppression of cervical cancer cell survival by ursolic acid extracted from Catalpa bungei leaves. Pharmacogn. Mag. 2018, 14, 425–431. [Google Scholar]
- Cao, S.; Zhang, J.; Chen, L.; Zhao, T. Ecosystem water imbalances created during ecological restoration by afforestation in China, and lessons for other developing countries. J. Environ. Manag. 2016, 183, 843–849. [Google Scholar] [CrossRef]
- Beniston, M. Climatic change in mountain regions: A review of possible impacts. Clim. Change 2003, 59, 5–31. [Google Scholar] [CrossRef]
- Sánchez-Benítez, A.; García-Herrera, R.; Barriopedro, D.; Sousa, P.M.; Trigo, R.M. June 2017: The earliest European summer mega-heatwave of reanalysis period. Geophys. Res. Lett. 2018, 45, 1955–1962. [Google Scholar] [CrossRef]
- Elisa, P.; Alessandro, P.; Andrea, A.; Silvia, B.; Mathis, P.; Dominik, P.; Manuela, R.; Francesca, T.; Voglar, G.E.; Tine, G. Environmental and climate change impacts of eighteen biomass-based plants in the alpine region: A comparative analysis. J. Clean. Prod. 2020, 242, 118449. [Google Scholar] [CrossRef]
- Rosa, R.; Simas, C.; Ataíde, R.; Soares, P.; Tomé, M. Optimal forest management under climate change variability. Ecol. Econ. 2024, 225, 108322. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, G.; Xu, Z. Driving mechanisms of climate-plant-soil patterns on the structure and function of different grasslands along environmental gradients in Tibetan and Inner Mongolian Plateaus in China. J. Clean. Prod. 2022, 339, 130696. [Google Scholar] [CrossRef]
- Quan, J.E.; Ni, R.; Wang, Y.; Sun, J.; Ma, M.; Bi, H. Effects of different growth regulators on the rooting of Catalpa bignonioides softwood cuttings. Life 2022, 12, 1231. [Google Scholar] [CrossRef]
- Manzini, J.; Hoshika, Y.; Sicard, P.; De Marco, A.; Ferrini, F.; Pallozzi, E.; Neri, L.; Baraldi, R.; Paoletti, E.; Moura, B.B. Detection of morphological and eco-physiological traits of ornamental woody species to assess their potential Net O3 uptake. Environ. Res. 2024, 252, 118844. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, B.; Fei, Y.; Yang, X.; Zhao, L.; Shi, C.; Zhang, Y.; Lu, N.; Wu, C.; Ma, W. Genetic architecture of leaf morphology revealed by integrated trait module in Catalpa bungei. Hortic. Res. 2023, 10, uhad032. [Google Scholar] [CrossRef]
- Kuniyal, C.P.; Rawat, Y.S.; Oinam, S.S.; Kuniyal, J.C.; Vishvakarma, S.C. Kuth (Saussurea lappa) cultivation in the cold desert environment of the Lahaul valley, northwestern Himalaya, India: Arising threats and need to revive socio-economic values. Biodivers. Conserv. 2005, 14, 1035–1045. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A. A standard protocol for reporting species distribution models. Ecography 2020, 43, 1261–1277. [Google Scholar] [CrossRef]
- Rivers, M.C.; Taylor, L.; Brummitt, N.A.; Meagher, T.R.; Roberts, D.L.; Nic Lughadha, E. How many herbarium specimens are needed to detect threatened species? Biol. Conserv. 2011, 144, 2541–2547. [Google Scholar] [CrossRef]
- Bakkenes, M.; Eickhout, B.; Alkemade, R. Impacts of different climate stabilisation scenarios on plant species in Europe. Glob. Environ. Change 2006, 16, 19–28. [Google Scholar] [CrossRef]
- Ma, L.; Pan, J. Spatial identification and priority conservation areas determination of wilderness in China. J. Clean. Prod. 2024, 451, 142069. [Google Scholar] [CrossRef]
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Adjemian, J.C.; Girvetz, E.H.; Beckett, L.; Foley, J.E. Analysis of Genetic Algorithm for Rule-Set Production (GARP) modeling approach for predicting distributions of fleas implicated as vectors of plague, Yersinia pestis, in California. J. Med. Entomol. 2006, 43, 93–103. [Google Scholar]
- Roigé, M.; Phillips, C.B. Validation and uncertainty analysis of the match climates regional algorithm (CLIMEX) for Pest risk analysis. Ecol. Inform. 2021, 61, 101196. [Google Scholar] [CrossRef]
- Costa, H.; Medeiros, V.; Azevedo, E.; Silva, L. Evaluating ecological-niche factor analysis as a modelling tool for environmental weed management in island systems. Weed Res. 2013, 53, 221–230. [Google Scholar] [CrossRef]
- Carpenter, G.; Gillison, A.N.; Winter, J. DOMAIN: A flexible modelling procedure for mapping potential distributions of plants and animals. Biodivers. Conserv. 1993, 2, 667–680. [Google Scholar] [CrossRef]
- Xu, T.; Hutchinson, M.F. New developments and applications in the ANUCLIM spatial climatic and bioclimatic modelling package. Environ. Model. Softw. 2013, 40, 267–279. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Wan, J.-Z.; Wang, C.-J.; Yu, F.-H. Effects of occurrence record number, environmental variable number, and spatial scales on MaxEnt distribution modelling for invasive plants. Biologia 2019, 74, 757–766. [Google Scholar] [CrossRef]
- Vignali, S.; Barras, A.G.; Arlettaz, R.; Braunisch, V. SDMtune: An R package to tune and evaluate species distribution models. Ecol. Evol. 2020, 10, 11488–11506. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Broennimann, O.; Treier, U.A.; Müller-Schärer, H.; Thuiller, W.; Peterson, A.; Guisan, A. Evidence of climatic niche shift during biological invasion. Ecol. Lett. 2007, 10, 701–709. [Google Scholar] [CrossRef]
- Fourcade, Y.; Besnard, A.G.; Secondi, J. Paintings predict the distribution of species, or the challenge of selecting environmental predictors and evaluation statistics. Glob. Ecol. Biogeogr. 2018, 27, 245–256. [Google Scholar] [CrossRef]
- Wu, K.; Wang, Y.; Liu, Z.; Huo, W.; Cao, J.; Zhao, G.; Zhang, F.-G. Prediction of potential invasion of two weeds of the genus Avena in Asia under climate change based on Maxent. Sci. Total Environ. 2024, 950, 175192. [Google Scholar] [CrossRef]
- Zhao, R.; Wang, S.; Chen, S. Predicting the potential habitat suitability of saussurea species in China under future climate scenarios using the optimized maximum entropy (maxent) model. J. Clean. Prod. 2024, 474, 143552. [Google Scholar] [CrossRef]
- Jian, S.; Zhu, T.; Wang, J.; Yan, D. The current and future potential geographical distribution and evolution process of Catalpa bungei in China. Forests 2022, 13, 96. [Google Scholar] [CrossRef]
- Liu, X.-T.; Yuan, Q.; Ni, J. Research advances in modelling plant species distribution in China. Chin. J. Plant Ecol. 2019, 43, 273. [Google Scholar] [CrossRef]
- Li, D.; Li, Z.; Liu, Z.; Yang, Y.; Khoso, A.G.; Wang, L.; Liu, D. Climate change simulations revealed potentially drastic shifts in insect community structure and crop yields in China’s farmland. J. Pest Sci. 2023, 96, 55–69. [Google Scholar] [CrossRef]
- Su, B.; Huang, J.; Mondal, S.K.; Zhai, J.; Wang, Y.; Wen, S.; Gao, M.; Lv, Y.; Jiang, S.; Jiang, T. Insight from CMIP6 SSP-RCP scenarios for future drought characteristics in China. Atmos. Res. 2021, 250, 105375. [Google Scholar] [CrossRef]
- Yang, L.; Jia, H.; Hua, Q. Predicting suitable habitats of parasitic desert species based on Biomod2 ensemble model: Cynomorium songaricum rupr and its host plants as an example. BMC Plant Biol. 2025, 25, 351. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, Z.; Zhu, F.; Gao, B. The impact of global warming on the potential suitable planting area of Pistacia chinensis is limited. Sci. Total Environ. 2023, 864, 161007. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Dudik, M.; Phillips, S.J.; Schapire, R.E. Performance guarantees for regularized maximum entropy density estimation. In Proceedings of the International Conference on Computational Learning Theory, Banff, AB, Canada, 1–4 July 2004; pp. 472–486. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef]
- Alanís-Méndez, J.L.; Soto, V.; Limón-Salvador, F. Effects of Climate Change on the Distribution of Prosthechea mariae (Orchidaceae) and within Protected Areas in Mexico. Plants 2024, 13, 839. [Google Scholar] [CrossRef]
- Luo, M.; Yang, P.; Yang, L.; Zheng, Z.; Chen, Y.; Li, H.; Wu, M. Predicting potentially suitable Bletilla striata habitats in China under future climate change scenarios using the optimized MaxEnt model. Sci. Rep. 2025, 15, 21231. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Wu, H.; Li, C.; Luo, G.; Zhao, T.; Chen, C.; Liu, Y.; Duan, M.; Wang, C. A Simulation of a Suitable Habitat for Acer yangbiense and Cinnamomum chago Under Climate Change. Forests 2025, 16, 621. [Google Scholar] [CrossRef]
- Zhao, Y.; Deng, X.; Xiang, W.; Chen, L.; Ouyang, S. Predicting potential suitable habitats of Chinese fir under current and future climatic scenarios based on Maxent model. Ecol. Inform. 2021, 64, 101393. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, C.; Shi, X.; Bo, X.; Li, S.; Shang, M.; Chen, F.; Chu, Q. Modeling climatically suitable areas for soybean and their shifts across China. Agric. Syst. 2021, 192, 103205. [Google Scholar] [CrossRef]
- Yang, H.; Viña, A.; Tang, Y.; Zhang, J.; Wang, F.; Zhao, Z.; Liu, J. Range-wide evaluation of wildlife habitat change: A demonstration using Giant Pandas. Biol. Conserv. 2017, 213, 203–209. [Google Scholar] [CrossRef]
- Yang, W.; Sun, S.; Wang, N.; Fan, P.; You, C.; Wang, R.; Zheng, P.; Wang, H. Dynamics of the distribution of invasive alien plants (Asteraceae) in China under climate change. Sci. Total Environ. 2023, 903, 166260. [Google Scholar] [CrossRef] [PubMed]
- Junliang, X.; Jiayu, H.; Tong, W.; Lexin, Z.; Pengfei, L.; Miao, W.; Yiping, Z. Intra-annual growth and its response to climatic factors in four ring-porous wood species. J. Zhejiang AF Univ. 2024, 41, 1105–1113. [Google Scholar]
- Çoban, H.O.; Örücü, Ö.K.; Arslan, E.S. MaxEnt Modeling for Predicting the Current and Future Potential Geographical Distribution of Quercus libani Olivier. Sustainability 2020, 12, 2671. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, W.; Fu, J. Vegetation response to precipitation anomalies under different climatic and biogeographical conditions in China. Sci. Rep. 2020, 10, 830. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H. Do they? How do they? WHY do they differ? On finding reasons for differing performances of species distribution models. Ecography 2009, 32, 66–77. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Peterson, A.T.; Soberón, J.; Overton, J.; Aragón, P.; Lobo, J.M. Use of niche models in invasive species risk assessments. Biol. Invasions 2011, 13, 2785–2797. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Li, C.; Liu, Z. Optimized maxent model predictions of climate change impacts on the suitable distribution of Cunninghamia lanceolata in China. Forests 2020, 11, 302. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J.; Hijmans, M.R.J. Package ‘dismo’. Circles 2017, 9, 1–68. [Google Scholar]
- Aidoo, O.F.; Souza, P.G.C.; da Silva, R.S.; Santana, P.A., Jr.; Picanço, M.C.; Kyerematen, R.; Sètamou, M.; Ekesi, S.; Borgemeister, C. Climate-induced range shifts of invasive species (Diaphorina citri Kuwayama). Pest Manag. Sci. 2022, 78, 2534–2549. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Powers, J.; Cochard, H.; Choat, B. Hanging by a thread? Forests and drought. Science 2020, 368, 261–266. [Google Scholar] [CrossRef]
- Zhou, W.; Chen, F.; Meng, Y.; Chandrasekaran, U.; Luo, X.; Yang, W.; Shu, K. Plant waterlogging/flooding stress responses: From seed germination to maturation. Plant Physiol. Biochem. 2020, 148, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jing, Z.; Li, Z.; Liu, Y.; Fang, S. Predictive modeling of suitable habitats for Cinnamomum Camphora (L.) presl using maxent model under climate change in China. Int. J. Environ. Res. Public Health 2019, 16, 3185. [Google Scholar] [CrossRef]
- Li, T.; Zhang, J.; Zhang, H.; Niu, S.; Qian, J.; Chen, Z.; Ma, T.; Meng, Y.; Di, B. Identification of Catalpa bungei Aquaporin Gene Family Related to Low Temperature Stress. Forests 2024, 15, 1063. [Google Scholar] [CrossRef]
- Pederson, G.T.; Stahle, D.; McWethy, D.B.; Toohey, M.; Jungclaus, J.; Lee, C.; Martin, J.; Alt, M.; Kichas, N.; Chellman, N. Dynamic treeline and cryosphere response to pronounced mid-Holocene climatic variability in the US Rocky Mountains. Proc. Natl. Acad. Sci. USA 2025, 122, e2412162121. [Google Scholar] [CrossRef]
- Wu, J.; Su, Y.; Wang, J.; He, Q.; Qiu, Q.; Ma, J.; Li, J. Morphological and physiological acclimation of Catalpa bungei plantlets to different light conditions. Photosynthetica 2018, 56, 537–548. [Google Scholar] [CrossRef]
- Omann, I.; Stocker, A.; Jäger, J. Climate change as a threat to biodiversity: An application of the DPSIR approach. Ecol. Econ. 2009, 69, 24–31. [Google Scholar] [CrossRef]
- Yang, X.; Zhou, B.; Xu, Y.; Han, Z. CMIP6 evaluation and projection of temperature and precipitation over China. Adv. Atmos. Sci. 2021, 38, 817–830. [Google Scholar] [CrossRef]
- Zhang, C.; Xiao, C.; Li, S.; Ren, Y.; Zhang, S.; Cai, X.; Sangbu, Z. Analysis of the composite risk grade for multi extreme climate events in China in recent 60 years. Climate 2023, 11, 191. [Google Scholar] [CrossRef]
- Wang, Y.; Bai, X.; Lei, Y.; Ding, W.; Wu, S. The Projection of Extreme Heat and Precipitation Events in China Response to Global Warming Under the SSP1-2.6 and SSP5-8.5 Scenarios. Int. J. Climatol. 2025, 45, e8807. [Google Scholar] [CrossRef]
- Tang, C.Q.; Dong, Y.-F.; Herrando-Moraira, S.; Matsui, T.; Ohashi, H.; He, L.-Y.; Nakao, K.; Tanaka, N.; Tomita, M.; Li, X.-S. Potential effects of climate change on geographic distribution of the Tertiary relict tree species Davidia involucrata in China. Sci. Rep. 2017, 7, 43822. [Google Scholar] [CrossRef]
- Xiao, F.; She, Y.; She, J.; Zhang, J.; Zhang, X.; Luo, C. Assessing habitat suitability and selecting optimal habitats for relict tree Cathaya argyrophylla in Hunan, China: Integrating pollen size, environmental factors, and niche modeling for conservation. Ecol. Indic. 2022, 145, 109669. [Google Scholar] [CrossRef]
- Wang, L.; Liu, J.; Liu, J.; Wei, H.; Fang, Y.; Wang, D.; Chen, R.; Gu, W. Revealing the long-term trend of the global-scale Ginkgo biloba distribution and the impact of future climate change based on the ensemble modeling. Biodivers. Conserv. 2023, 32, 2077–2100. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Variable Description | Unit |
|---|---|---|
| Bio1 | Annual Mean Temperature | °C |
| Bio2 | Mean Diurnal Range | °C |
| Bio3 | Isothermality | – |
| Bio4 | Temperature Seasonality | – |
| Bio5 | Max Temperature of Warmest Month | °C |
| Bio6 | Min Temperature of Coldest Month | °C |
| Bio7 | Annual Temperature Range | °C |
| Bio8 | Mean Temperature of Wettest Quarter | °C |
| Bio9 | Mean Temperature of Driest Quarter | °C |
| Bio10 | Mean Temperature of Warmest Quarter | °C |
| Bio11 | Mean Temperature of Coldest Quarter | °C |
| Bio12 | Annual Precipitation | mm |
| Bio13 | Precipitation of Wettest Month | mm |
| Bio14 | Precipitation of Driest Month | mm |
| Bio15 | Precipitation Seasonality | – |
| Bio16 | Precipitation of Wettest Quarter | mm |
| Bio17 | Precipitation of Driest Quarter | mm |
| Bio18 | Precipitation of Warmest Quarter | mm |
| Bio19 | Precipitation of Coldest Quarter | mm |
| Ele | Elevation | m |
| Slp | Slope | ° |
| Asp | Aspect | – |
| Factor | PC (%) | PI (%) | RTGw | RTGO | TGW | TGO | AUCw | AUCO |
|---|---|---|---|---|---|---|---|---|
| Bio3 | 1.88 | 0.69 | 1.21 | 0.14 | 1.31 | 0.26 | 0.90 | 0.72 |
| Bio5 | 1.62 | 1.23 | 1.21 | 0.33 | 1.31 | 0.33 | 0.90 | 0.71 |
| Bio6 | 79.87 | 76.17 | 0.86 | 1.00 | 1.01 | 1.05 | 0.87 | 0.86 |
| Bio12 | 6.28 | 7.12 | 1.17 | 0.48 | 1.26 | 0.57 | 0.89 | 0.77 |
| Bio15 | 0.86 | 1.55 | 1.20 | 0.18 | 1.30 | 0.24 | 0.90 | 0.69 |
| Slp | 4.29 | 5.17 | 1.15 | 0.01 | 1.21 | 0.03 | 0.89 | 0.53 |
| Asp | 1.01 | 0.39 | 1.20 | 0.01 | 1.31 | 0.00 | 0.90 | 0.51 |
| Ele | 4.18 | 7.68 | 1.20 | 0.32 | 1.28 | 0.31 | 0.90 | 0.70 |
| Suitability Level | Bio5 (°C) | Bio6 (°C) | Bio12 (mm) | Elevation (m) | ||||
|---|---|---|---|---|---|---|---|---|
| Range | Mean | Range | Mean | Range | Mean | Range | Mean | |
| Low | 16.2–23.6 | 20.8 | 16.1–9.9 | −3.0 | 242–458 | 380 | 2040–3628 | 2081 |
| Moderate | 23.6–39.9 | 32.1 | 12.1–4.8 | −3.5 | 458–542 | 3005 | 889–2040 | 1427 |
| Extreme | 28.1–33.6 or 33.9–47.0 | 37.6 | −9.8–3.1 | −3.5 | 1630–4334 | 1086 | −154–830 | 339 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.; Zhao, J.; Wang, Y.; Wu, G.; Hou, Y.; Yu, C. Optimized MaxEnt Modeling of Catalpa bungei Habitat for Sustainable Management Under Climate Change in China. Forests 2025, 16, 1150. https://doi.org/10.3390/f16071150
Shi X, Zhao J, Wang Y, Wu G, Hou Y, Yu C. Optimized MaxEnt Modeling of Catalpa bungei Habitat for Sustainable Management Under Climate Change in China. Forests. 2025; 16(7):1150. https://doi.org/10.3390/f16071150
Chicago/Turabian StyleShi, Xiaomeng, Jingshuo Zhao, Yanlin Wang, Guichun Wu, Yingjie Hou, and Chunyan Yu. 2025. "Optimized MaxEnt Modeling of Catalpa bungei Habitat for Sustainable Management Under Climate Change in China" Forests 16, no. 7: 1150. https://doi.org/10.3390/f16071150
APA StyleShi, X., Zhao, J., Wang, Y., Wu, G., Hou, Y., & Yu, C. (2025). Optimized MaxEnt Modeling of Catalpa bungei Habitat for Sustainable Management Under Climate Change in China. Forests, 16(7), 1150. https://doi.org/10.3390/f16071150