
Long-Term Nitrogen Addition Regulates Plant-Soil 15N–13C Coupling Through Species Traits and Temporal-Spatial Dynamics in a Temperate Forest
Abstract
1. Introduction
2. Materials and Methods
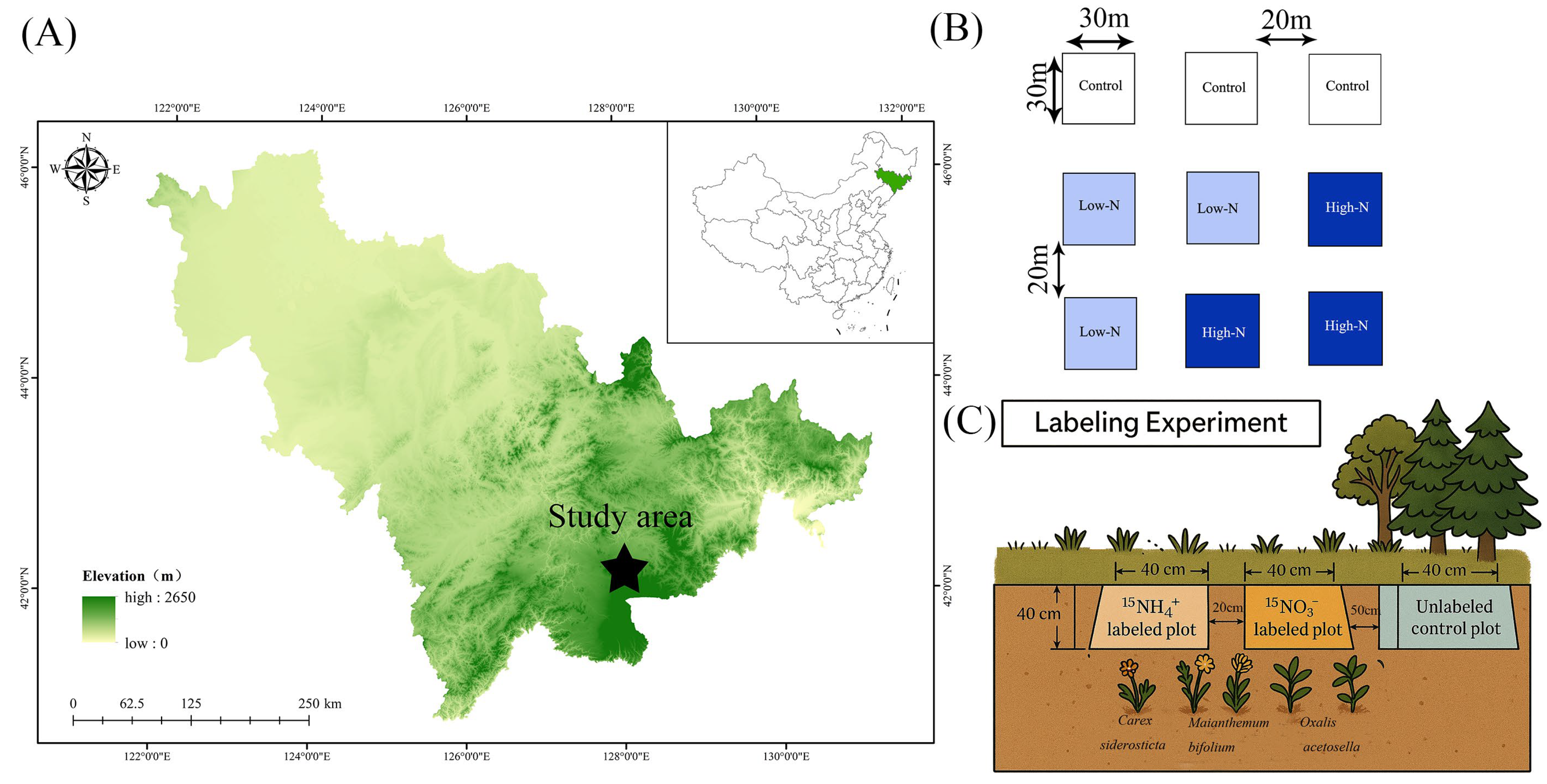
2.1. Study Sites
2.2. Experiment Design
2.3. 15N and 13C Labeling Experiment
2.3.1. 15N Labeling
2.3.2. 13C Labeling
2.4. Sampling Strategy
2.4.1. Plant Sampling
2.4.2. Soil Sampling
2.5. Measurement of Biomass, Nutrient Status, and Isotopic Abundances
2.6. Soil Physicochemical and Microbial Properties Analysis
2.7. Calculations and Statistical Analysis
3. Results
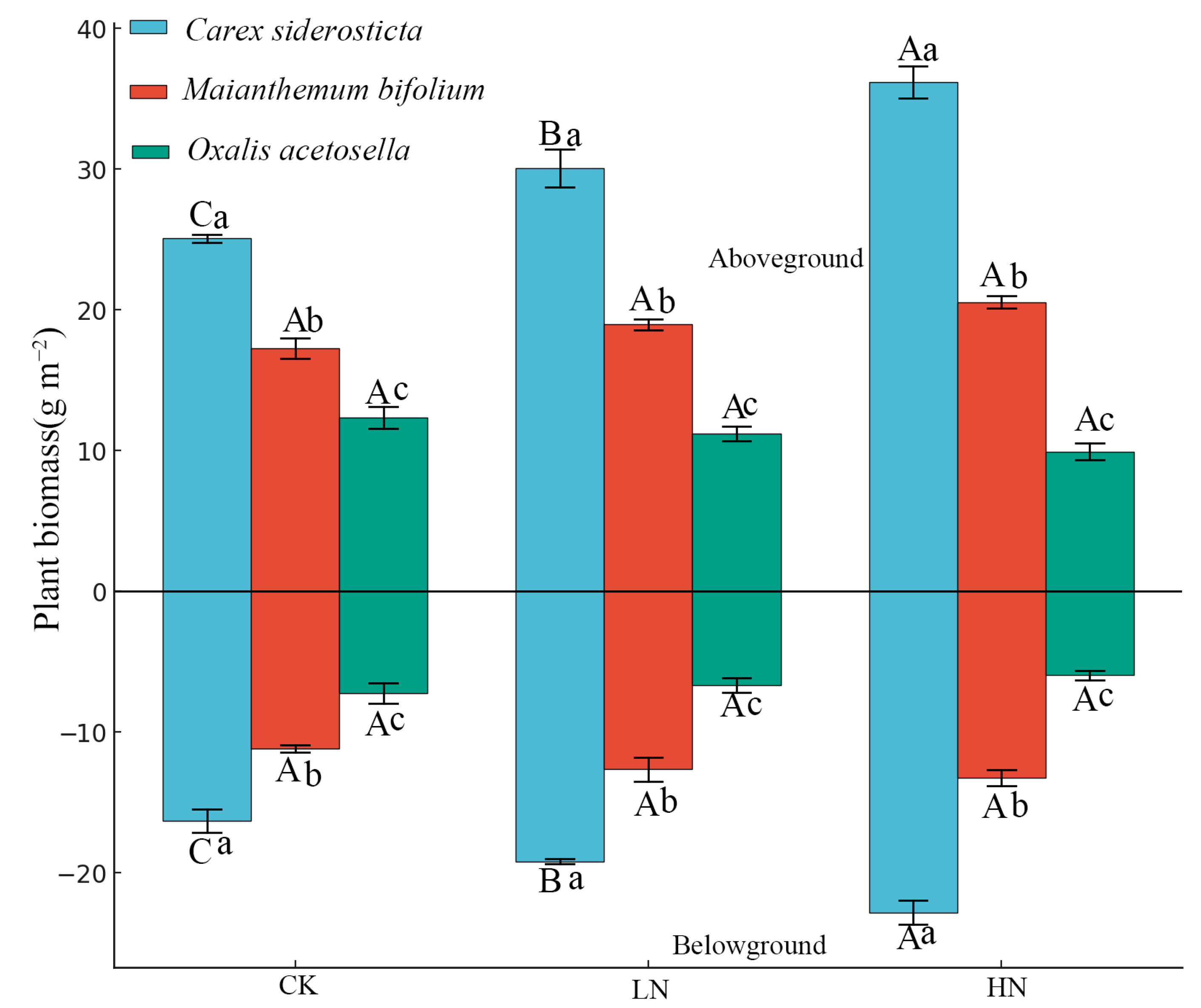
3.1. Effects of Nitrogen Addition on Plant Biomass and Soil Physicochemical Properties
3.2. Nitrogen Uptake and 13C Assimilation
3.3. Correlation Between Plant Biomass and Nitrogen or 13CO2 Assimilation
3.4. Soil 15N-TN and 13C-SOC Content
3.5. PCA and Pearson’s Correlation of Soil and Plant Variables
4. Discussion
4.1. Species-Specific Biomass Responses Mediated by Nitrogen-Induced Soil Property Shifts
4.2. Interactive Effects of Nitrogen Form, Timing, and Species Identity on Plant Nitrogen and Carbon Assimilation
4.3. Enhanced Coupling Between Biomass Accumulation and Nutrient Assimilation Under High Nitrogen Conditions
4.4. Integrated Regulation of Soil Nitrogen and Carbon Retention by Nitrogen Form, Depth, and Temporal Dynamics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilliam, F.S. Response of temperate forest ecosystems under decreased nitrogen deposition: Research challenges and opportunities. Forests 2021, 12, 509. [Google Scholar] [CrossRef]
- Lu, X.; Vitousek, P.M.; Mao, Q.; Gilliam, F.S.; Luo, Y.; Turner, B.L.; Zhou, G.; Mo, J. Nitrogen deposition accelerates soil carbon sequestration in tropical forests. Proc. Natl. Acad. Sci. USA 2021, 118, e2020790118. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Rocci, K.S.; Lehmann, A.; Rillig, M.C. Nitrogen increases soil organic carbon accrual and alters its functionality. Glob. Change Biol. 2023, 29, 1971–1983. [Google Scholar] [CrossRef]
- Franklin, J.; Serra-Diaz, J.M.; Syphard, A.D.; Regan, H.M. Global change and terrestrial plant community dynamics. Proc. Natl. Acad. Sci. USA 2016, 113, 3725–3734. [Google Scholar] [CrossRef]
- Sandhu, N.; Kumar, A. Bridging the rice yield gaps under drought: QTLs, genes, and their use in breeding programs. Agronomy 2017, 7, 27. [Google Scholar] [CrossRef]
- Naoura, G.; Emendack, Y.; Sawadogo, N.; Djirabaye, N.; Tabo, R.; Laza, H.; Atchozou, E.A. Assessment of photoperiod sensitivity and the effect of sowing date on dry-season sorghum cultivars in southern Chad. Agronomy 2023, 13, 932. [Google Scholar] [CrossRef]
- Mdutyana, M.; Thomalla, S.J.; Philibert, R.; Ward, B.B.; Fawcett, S.E. The seasonal cycle of nitrogen uptake and nitrification in the Atlantic sector of the Southern Ocean. Glob. Biogeochem. Cycles 2020, 34, e2019GB006363. [Google Scholar] [CrossRef]
- Zakem, E.J.; Bayer, B.; Qin, W.; Santoro, A.E.; Zhang, Y.; Levine, N.M. Controls on the relative abundances and rates of nitrifying microorganisms in the ocean. Biogeosciences 2022, 19, 5401–5418. [Google Scholar] [CrossRef]
- Lü, X.T.; Reed, S.C.; Yu, Q.; Han, X.G. Nutrient resorption helps drive intra-specific coupling of foliar nitrogen and phosphorus under nutrient-enriched conditions. Plant Soil 2016, 398, 111–120. [Google Scholar] [CrossRef]
- Zou, Y.; Tang, X.; Cui, L.; Li, H.; Lei, Y.; Zhao, X.; Zhai, X.; Li, J.; Wang, J.; Wang, R.; et al. Analysis of coupled wetland soil–plant ecological stoichiometric characteristics of carbon, nitrogen, and phosphorus under different disturbance intensities. Plant Soil 2024, in press. [Google Scholar] [CrossRef]
- Gerber, S.; Hedin, L.O.; Oppenheimer, M.; Pacala, S.W.; Shevliakova, E. Nitrogen cycling and feedbacks in a global dynamic land model. Glob. Biogeochem. Cycles 2010, 24, GB1001. [Google Scholar] [CrossRef]
- Wang, W.; Li, C.; Shu, J.; Chen, W. Changes of vegetation in southern China. Sci. China Earth Sci. 2019, 62, 1316–1328. [Google Scholar] [CrossRef]
- Schwede, D.B.; Simpson, D.; Tan, J.; Fu, J.S.; Dentener, F.; Du, E.; deVries, W. Spatial variation of modelled total, dry and wet nitrogen deposition to forests at global scale. Environ. Pollut. 2018, 243, 1287–1301. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ding, Y.; Tian, Y.; Xu, X.; Gunina, A. Exploring grazing intensity effects: Nitrogen uptake in grassland species and soil carbon allocation. Plant Soil 2024, 504, 833–846. [Google Scholar] [CrossRef]
- Xie, L.; Zhao, Y.; Wang, C.; Xu, S.; Fan, R.; Gao, H.; Xu, J.; Zhu, S.; Lei, W.; Han, S. Simultaneous determination of nitrogen content and 15N isotope abundance in plants using peak-height intensities at m/z 28 and 29. Talanta 2024, 275, 126078. [Google Scholar] [CrossRef]
- Mori, T.; Wang, S.; Wang, C.; Mo, J.; Zhang, W. Is microbial biomass measurement by the chloroform-fumigation extraction method biased by experimental addition of N and P? iForest Biogeosciences For. 2021, 14, 408. [Google Scholar] [CrossRef]
- Shibahara, F.; Inubushi, K. Measurements of microbial biomass C and N in paddy soils by the fumigation-extraction method. Soil Sci. Plant Nutr. 1995, 41, 681–689. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Peñuelas, J.; Sardans, J.; Cheng, D.; Yu, H.; Zhong, Q. Resource-acquisitive species have greater plasticity in leaf functional traits than resource-conservative species in response to nitrogen addition in subtropical China. Sci. Total Environ. 2023, 903, 166177. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, J.; Xia, W.; Shao, Y.; Liu, Z.; Guo, J.; Qin, W.; Wan, L.; Liu, J.; Liu, Y. Influence of cover-crop root functional traits on sweet-potato yield and soil microbial communities. Microorganisms 2025, 13, 471. [Google Scholar] [CrossRef]
- Zhao, L.; Cui, J.; Fu, Y.; Liu, H. Specific rhizobacterial assembly enhances physiological adaptation of soil-cultivated wheat to simulated microgravity. Acta Astronaut. 2024, 223, 723–733. [Google Scholar] [CrossRef]
- Viancelli, A.; Michelon, W. Climate change and nitrogen dynamics: Challenges and strategies for a sustainable future. Nitrogen 2024, 5, 688–701. [Google Scholar] [CrossRef]
- Liu, Q.; Wan, W.; Chen, W.; Zhang, C.; Gao, H.; Zhang, J.; Sun, Z.; Li, H. Adaptation strategies of three legumes to soil phosphorus availability in steppes of Inner Mongolia. Plant Soil 2025, in press. [Google Scholar] [CrossRef]
- Ni, H.; Su, W.; Fan, S.; Chu, S. Intensive management practices affect rhizosphere soil properties, root growth, and nutrient uptake in Moso bamboo plantations. For. Ecol. Manag. 2021, 493, 119083. [Google Scholar] [CrossRef]
- Cao, D.; Wang, X.; Miao, Y.; Wu, C.; Zhang, H.; Wang, S.; Wang, F.; Chen, L.; Liang, C.; Kuzyakov, Y.; et al. Microbial strategies regulate organic-carbon accumulation in saline paddy soils: A millennium chronosequence. CATENA 2025, 252, 108869. [Google Scholar] [CrossRef]
- Sharma, U.C.; Datta, M.; Sharma, V. Toxicity and Tolerance. In Soil Acidity: Management Options for Higher Crop Productivity; Springer Nature: Cham, Switzerland, 2025; pp. 323–425. [Google Scholar] [CrossRef]
- Gemada, A.R. Soil acidity challenges to crop production in Ethiopian Highlands and management strategies for mitigation. Agric. For. Fish. 2021, 10, 245–261. [Google Scholar] [CrossRef]
- Revillini, D.; Gehring, C.A.; Johnson, N.C. Locally adapted mycorrhizas and rhizobacteria in plant-soil feedback systems. Funct. Ecol. 2016, 30, 1086–1098. [Google Scholar] [CrossRef]
- Malik, A.A.; Martiny, J.B.; Brodie, E.L.; Martiny, A.C.; Treseder, K.K.; Allison, S.D. Defining trait-based microbial strategies and their consequences for soil carbon cycling under climate change. ISME J. 2020, 14, 1–9. [Google Scholar] [CrossRef]
- Qiao, J.; Zuo, X.; Yue, P.; Wang, S.; Hu, Y.; Guo, X.; Li, X.; Lv, P.; Guo, A.; Sun, S. High nitrogen addition induces functional-trait divergence of plant community in a temperate desert steppe. Plant Soil 2023, 487, 133–156. [Google Scholar] [CrossRef]
- Peng, W.; Wang, C.D.; Wang, X.L.; Wu, Y.H.; Zhang, Y.; Sun, Y.G.; Shi, Y.; Mi, G.H. Mixed NO3− and NH4+ supply promotes growth of maize seedlings via enhanced nitrogen absorption and assimilation. J. Integr. Agric. 2023, 22, 1896–1908. [Google Scholar] [CrossRef]
- Hu, J.; Zheng, Q.; Neuhäuser, B.; Dong, C.; Tian, Z.; Dai, T. Superior glucose metabolism supports NH4+ assimilation in wheat to improve ammonium tolerance. Front. Plant Sci. 2024, 15, 1339105. [Google Scholar] [CrossRef]
- Long, D.; Lin, G.; Yan, P.; Yang, H.; Deng, K.; Guo, J. Energy distribution for N, P and C sequestration by Microcystis aeruginosa under blue and orange light. J. Environ. Chem. Eng. 2024, 12, 112054. [Google Scholar] [CrossRef]
- Zhang, L.; Feng, Y.; Zhao, Z.; Baoyin, B.; Cui, Z.; Wang, H.; Li, Q.; Cui, J. Macrogenomics-based analysis of intercropped soybean photosynthetic characteristics and nitrogen-assimilating enzyme activities under different nitrogen levels. Microorganisms 2024, 12, 1220. [Google Scholar] [CrossRef]
- Tausz-Posch, S.; Tausz, M.; Bourgault, M. Elevated [CO2] effects on crops: Advances in understanding acclimation, nitrogen dynamics and interactions with drought and other organisms. Plant Biol. 2020, 22, 38–51. [Google Scholar] [CrossRef] [PubMed]
- De Frenne, P.; Brunet, J.; Cougnon, M.; Decocq, G.; Graae, B.J.; Hagenblad, J.; Hermy, M.; Kolb, A.; Lemke, I.H.; Ma, S.; et al. Biological flora of the British Isles: Milium effusum. J. Ecol. 2017, 105, 839–858. [Google Scholar] [CrossRef]
- Goudiaby, V.; Schneider, R.; Brais, S.; Raulier, F.; Berninger, F. Competition and shade tolerance effects on carbon allocation: Insights from a carbon-balance model. Forests 2022, 13, 572. [Google Scholar] [CrossRef]
- Vermaat, J.E. Linking clonal growth patterns and ecophysiology to predict meadow-scale dynamics of seagrass beds. Perspect. Plant Ecol. Evol. Syst. 2009, 11, 137–155. [Google Scholar] [CrossRef]
- Klimešová, J.; Martínková, J. Clonal Growth, Resprouting, and Vegetative Propagation of Weeds. In Persistence Strategies of Weeds; Wiley-Blackwell: Hoboken, NJ, USA, 2022; pp. 200–218. [Google Scholar] [CrossRef]
- Duan, S.; Feng, G.; Limpens, E.; Bonfante, P.; Xie, X.; Zhang, L. Cross-kingdom nutrient exchange in the plant–arbuscular mycorrhizal fungus–bacterium continuum. Nat. Rev. Microbiol. 2024, 22, 773–790. [Google Scholar] [CrossRef]
- Moreau, D.; Bardgett, R.D.; Finlay, R.D.; Jones, D.L.; Philippot, L. A plant perspective on nitrogen cycling in the rhizosphere. Funct. Ecol. 2019, 33, 540–552. [Google Scholar] [CrossRef]
- González de Andrés, E. Climate–nutrient cycle interactions governing forest responses to global change: The role of mixed forests. Forests 2019, 10, 609. [Google Scholar] [CrossRef]
- Chandrasekaran, M. Arbuscular mycorrhizal fungi enhance biomass, root morphological traits and nutrient uptake under drought stress: A meta-analysis. J. Fungi 2022, 8, 660. [Google Scholar] [CrossRef]
- Paré, D.; Thiffault, E. Nutrient budgets in forests under increased biomass-harvesting scenarios. Curr. For. Rep. 2016, 2, 81–91. [Google Scholar] [CrossRef]
- Iqbal, A.; Dong, Q.; Wang, X.; Gui, H.; Zhang, H.; Zhang, X.; Song, M. High nitrogen enhances drought tolerance in cotton via antioxidant enzymes, nitrogen metabolism and osmotic adjustment. Plants 2020, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G.; Hodges, M. Respiration and nitrogen assimilation: Targeting mitochondria-associated metabolism to enhance nitrogen-use efficiency. J. Exp. Bot. 2011, 62, 1467–1482. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, X.; Li, G.; Liu, F. Elevated CO2 modulates carbon assimilation and leaf water-use efficiency of Nicotiana tabacum under patchy soil-nutrient deficiency. Ind. Crops Prod. 2021, 166, 113500. [Google Scholar] [CrossRef]
- Dalling, J.W.; Flores, M.R.; Heineman, K.D. Wood nutrients: Under-explored traits with functional and biogeochemical consequences. New Phytol. 2024, 244, 1694–1708. [Google Scholar] [CrossRef]
- Ding, D.; Liu, M.; Arif, M.; Yuan, Z.; Li, J.; Hu, X.; Zheng, J.; Li, C. Ecological stoichiometry (C:N:P) responses to periodic submergence in mega-reservoirs: Growth of Taxodium distichum and Taxodium ascendens. Plants 2021, 10, 2040. [Google Scholar] [CrossRef]
- Wipf, H.M.L.; Xu, L.; Gao, C.; Spinner, H.B.; Taylor, J.; Lemaux, P.; Mitchell, J.; Coleman-Derr, D. Agricultural soil management practices differentially shape bacterial and fungal microbiomes of Sorghum bicolor. Appl. Environ. Microbiol. 2021, 87, e02345-20. [Google Scholar] [CrossRef]
- Broadbent, E.N.; Almeyda Zambrano, A.M.; Asner, G.P.; Soriano, M.; Field, C.B.; de Souza, H.R.; Peña-Claros, M.; Adams, R.I.; Dirzo, R.; Giles, L. Foliar nutrient dynamics during forest succession following slash-and-burn agriculture in the Bolivian Amazon. PLoS ONE 2014, 9, e86042. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, W.; Hou, X.; Li, Y.; Tong, J. Nutrient loads influence microbial-derived carbon accumulation in wetlands: Insights from microbial metabolic-investment strategies. Environ. Res. 2023, 217, 114981. [Google Scholar] [CrossRef]
- Schulte-Uebbing, L.; de Vries, W. Global-scale impacts of nitrogen deposition on tree carbon sequestration in tropical, temperate and boreal forests: A meta-analysis. Glob. Change Biol. 2018, 24, e416–e431. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, Z.; Deng, X.; Herbert, S.; Xing, B. Biochar addition enhances nitrogen retention and bioavailability in agricultural soil. Geoderma 2013, 206, 32–39. [Google Scholar] [CrossRef]
- Yadav, M.R.; Kumar, R.; Parihar, C.M.; Yadav, R.K.; Jat, S.L.; Ram, H.; Meena, R.K.; Singh, M.; Verma, A.P.; Kumar, U.J.A.R.; et al. Strategies for improving nitrogen-use efficiency: A review. Agric. Rev. 2017, 38, 29–40. [Google Scholar] [CrossRef]
- Li, Z.; Zeng, Z.; Song, Z.; Wang, F.; Tian, D.; Mi, W.; Huang, X.; Wang, J.; Song, L.; Yang, Z.; et al. Vital roles of soil microbes in driving terrestrial nitrogen immobilization. Glob. Change Biol. 2021, 27, 1848–1858. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Graham, N.J.D.; Su, Z.; Xu, L.; Yu, W. Decomposition of natural organic matter by biofiltration: Capabilities of microbial consortia from disparate matrices. Water Res. 2024, 262, 122047. [Google Scholar] [CrossRef]
- Meng, D.; Cheng, H.; Shao, Y.; Luo, M.; Xu, D.; Liu, Z.; Ma, L. Effects of nitrogen on transformation of soil organic carbon: Progress and perspectives. Processes 2022, 10, 2425. [Google Scholar] [CrossRef]
- Ye, R.; Parajuli, B.; Sigua, G. Subsurface clay-soil application improves aggregate stability, nitrogen availability and organic-carbon preservation in degraded ultisols with cover-crop mixtures. Soil Sci. Soc. Am. J. 2019, 83, 597–604. [Google Scholar] [CrossRef]
- Long, C.; Liu, Z.; Liu, R.; Yin, L.; Tan, F.; Wang, Y.; He, G. Disparities in soil microbial CO2-fixation rates under different vegetation at an acidic red-soil experimental station in China. Front. Microbiol. 2024, 15, 1480484. [Google Scholar] [CrossRef]
- Xing, Y.; Zhang, X.; Wang, X. Water-fertilizer coupling technology enhances soil health and crop yields. Front. Sustain. Food Syst. 2024, 8, 1494819. [Google Scholar] [CrossRef]
- Tipping, E.; Davies, J.A.C.; Henrys, P.A.; Kirk, G.J.; Lilly, A.; Dragosits, U.; Carnell, E.J.; Dore, A.J.; Sutton, M.A.; Tomlinson, S.J. Long-term increases in soil carbon driven by ecosystem fertilization from atmospheric nitrogen deposition: Evidence from modelling and observations. Sci. Rep. 2017, 7, 1890. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | CK | LN | HN | |||
|---|---|---|---|---|---|---|
| 0–5 cm | 5–15 cm | 0–5 cm | 5–15 cm | 0–5 cm | 5–15 cm | |
| Soil Moisture (%) | 31.46 ± 0.62 Ab | 35.27 ± 0.67 Aa | 29.68 ± 0.59 Bb | 33.59 ± 0.65 Ba | 27.92 ± 0.56 Cb | 31.72 ± 0.62 Ca |
| Soil pH | 5.81 ± 0.04 Ab | 6.01 ± 0.05 Aa | 5.51 ± 0.04 Bb | 5.71 ± 0.04 Ba | 5.38 ± 0.04 Ba | 5.51 ± 0.04 Ca |
| NH4+-N (µg Ng−1 Soil) | 10.23 ± 0.38 Ca | 8.69 ± 0.33 Cb | 14.76 ± 0.55 Ba | 12.48 ± 0.47 Bb | 18.99 ± 0.71 Aa | 16.11 ± 0.61 Ab |
| NO3−-N (µg Ng−1 Soil) | 4.12 ± 0.15 Ca | 3.29 ± 0.12 Cb | 6.53 ± 0.25 Ba | 5.18 ± 0.19 Bb | 9.44 ± 0.36 Aa | 7.54 ± 0.29 Ab |
| Total N (µmol N g−1) | 477.75 ± 18.11 Ba | 401.51 ± 15.24 Bb | 603.01 ± 22.82 Aa | 506.11 ± 19.18 Ab | 630.08 ± 23.89 Aa | 531.00 ± 20.01 Ab |
| SOC (µmol C g−1) | 9303.01 ± 279.11 Ba | 8373.03 ± 251.35 Bb | 10103.10 ± 302.99 Aa | 9067.05 ± 272.24 Ab | 10496.23 ± 314.86 Aa | 9446.12 ± 283.63 Ab |
| MBN (µg N g−1 Soil) | 216.11 ± 8.43 Ca | 176.01 ± 6.84 Cb | 258.21 ± 10.09 Ba | 205.06 ± 8.02 Bb | 288.06 ± 11.23 Aa | 228.07 ± 8.87 Ab |
| MBC (µg C g−1 Soil) | 1315.21 ± 52.61 Ba | 1189.02 ± 47.56 Cb | 1512.01 ± 60.48 Aa | 1369.11 ± 54.68 Bb | 1673.22 ± 66.99 Aa | 1534.01 ± 61.55 Ab |
| Source of Variation | Df | Aboveground Plant N Uptake | Belowground Plant N Uptake | Aboveground Plant 13C Assimilation | Belowground Plant 13C Assimilation |
|---|---|---|---|---|---|
| S | 2 | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| T | 2 | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| N | 1 | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| TP | 1 | <0.001 *** | <0.01 ** | <0.001 *** | <0.001 *** |
| S × T | 4 | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| S × N | 2 | <0.001 *** | <0.001 *** | <0.001 *** | <0.05 * |
| S × TP | 2 | <0.05 * | NS | <0.001 *** | <0.001 *** |
| T × N | 2 | <0.001 *** | <0.001 *** | <0.01 ** | NS |
| T × TP | 2 | NS | NS | <0.001 *** | <0.001 *** |
| N × TP | 1 | NS | NS | <0.001 *** | <0.001 *** |
| S × T × N | 4 | <0.001 *** | NS | <0.05 * | <0.05 * |
| S × T × TP | 4 | NS | <0.001 *** | <0.001 *** | <0.001 *** |
| S × N × TP | 2 | NS | NS | NS | NS |
| T × N × TP | 2 | NS | NS | NS | NS |
| S × T × N × TP | 4 | NS | NS | NS | NS |
| Source of Variation | Df | Soil 15N Retention | Soil 13C Assimilation |
|---|---|---|---|
| T | 2 | <0.001 *** | <0.05 * |
| N | 1 | <0.05 * | NS |
| TP | 1 | <0.001 *** | NS |
| D | 1 | NS | <0.001 *** |
| T × N | 2 | <0.001 *** | NS |
| T × TP | 2 | <0.001 *** | NS |
| T × D | 2 | <0.001 *** | <0.001 *** |
| N × TP | 1 | <0.001 *** | NS |
| N × D | 1 | NS | NS |
| TP × D | 1 | <0.05 * | <0.001 *** |
| T × N × TP | 2 | NS | NS |
| T × N × D | 2 | NS | NS |
| T × TP × D | 2 | NS | NS |
| N × TP × D | 1 | NS | NS |
| T × N × TP × D | 2 | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Li, Y. Long-Term Nitrogen Addition Regulates Plant-Soil 15N–13C Coupling Through Species Traits and Temporal-Spatial Dynamics in a Temperate Forest. Forests 2025, 16, 1046. https://doi.org/10.3390/f16071046
Zhou M, Li Y. Long-Term Nitrogen Addition Regulates Plant-Soil 15N–13C Coupling Through Species Traits and Temporal-Spatial Dynamics in a Temperate Forest. Forests. 2025; 16(7):1046. https://doi.org/10.3390/f16071046
Chicago/Turabian StyleZhou, Mingxin, and Yibo Li. 2025. "Long-Term Nitrogen Addition Regulates Plant-Soil 15N–13C Coupling Through Species Traits and Temporal-Spatial Dynamics in a Temperate Forest" Forests 16, no. 7: 1046. https://doi.org/10.3390/f16071046
APA StyleZhou, M., & Li, Y. (2025). Long-Term Nitrogen Addition Regulates Plant-Soil 15N–13C Coupling Through Species Traits and Temporal-Spatial Dynamics in a Temperate Forest. Forests, 16(7), 1046. https://doi.org/10.3390/f16071046