Abstract
Wooded grasslands are agroforestry systems of high biological and cultural value, which are increasingly threatened by land-use abandonment in Mediterranean marginal areas. In the central-southern Apennines, little is known about their ecological dynamics under different management regimes. This study assesses how three management intensities (High: mowing plus grazing; Low: grazing only; and Abandoned: no management for ~50 years) affect the wooded grasslands in a protected area of the Central Apennines. Vascular plant composition and cover were recorded along radial transects from isolated Fagus sylvatica L. trunks to the adjacent grassland, with plots grouped in four positions (Trunk, Mid-canopy, Edge, and Grassland). The canopy cover, shrub height, species richness, and ecological roles of species were analysed. The results show that light availability, driven by canopy and shrub cover, shapes a gradient from shade-adapted species near the trunk to heliophilous grassland species in open areas. In the Abandoned site, shrub encroachment reduces light even beyond the canopy, facilitating the spread of shade-tolerant and pre-forest species, accelerating succession towards a closed-canopy forest. High-intensity management preserves floristic gradients and grassland species, while Low-intensity management shows early signs of succession at the canopy edge. These findings highlight the importance of traditional mowing and grazing in maintaining the biodiversity and ecological functions of wooded grasslands and emphasize the need for timely interventions where management declines.
1. Introduction
Wooded grasslands are agroforestry systems of high biological and cultural value, recognized at the European level, yet they are currently undergoing decline and are at risk of disappearance. These ecosystems constitute a distinctive type of grassland characterized by isolated trees or groups of trees, collectively forming a complex, multifunctional ecotonal system positioned between closed-canopy forests and open grasslands. The vertical and horizontal structural heterogeneity characterizing these ecosystems determines the presence of a wide variety of microhabitats that are favourable to biodiversity [1,2,3,4,5], which results, in terms of gamma diversity, in higher diversity than that typically observed in open grasslands [4].
Wooded grasslands are anthropogenic systems created through forest thinning and maintained over centuries by traditional agro-silvo-pastoral practices, such as grazing, mowing, and coppicing [6,7], aimed at preserving the balance between herbaceous and tree species [8]. Already widespread in association with livestock farming since the Neolithic period [8,9,10], wooded grasslands have become integral components of distinctive local socio-cultural and economic histories across different territories [7,11]. Their strong dependency on anthropogenic activities—which have shaped and maintained them over centuries—makes wooded grassland systems particularly vulnerable to variations in management intensities (e.g., overgrazing or abandonment), a vulnerability that has been further compounded by recent climate change [7,12]. Specifically, the abandonment of traditional practices observed over recent decades—particularly in the marginal and mountainous areas of the Mediterranean basin due to socio-economic transformations [13,14] leading to progressive depopulation—represents the primary threat to the existence of wooded grasslands [8,9,15]. The underutilization and abandonment of traditional management practices have triggered natural dynamic processes of grassland afforestation, consequently leading to biodiversity losses with the deterioration of related ecosystem services [16] and landscape homogenization [17]. In areas that have been abandoned for a long time, ancient trees that were once isolated within grasslands have now become incorporated into closed-canopy forests, and often only aerial photographs and historical documentation allow for the reconstruction of previous vegetation landscape structures [18,19].
It is therefore important to identify actions for their restoration and conservation as also underlined by the inclusion of wooded grasslands in the European Red List of habitats (E7.1 “Temperate wooded pastures and meadows” Vulnerable; E7.2 “Hemiboreal and boreal wooded pastures and meadows” Critically Endangered; E7.3 “Mediterranean wooded pastures and meadows” Near Threatened) [20]. Regarding European habitat protection policies, only hemiboreal and Mediterranean wooded pastures and meadows are listed as habitats of community interest: “Fennoscandian wooded meadows” and “Fennoscandian wooded pastures” (codes 6530* and 9070, respectively) for hemiboreal regions, and “Dehesas with evergreen Quercus spp.” (code 6310) for the Mediterranean area (Annex I, Directive 92/43/EEC).
Most studies concerning the appropriate management for biodiversity conservation in wooded grasslands indicate that traditional management practices adopted across various regions have historically contributed to the existence of these multifunctional systems [18,21,22]. However, this is a complex issue, as the conservation management of wooded grasslands requires a holistic approach that considers a range of ecological, institutional, and social attributes, including biodiversity, ecosystem services, the institutions associated with wood–grassland systems, and their historical interactions [23]. Specifically, as advocated by several authors (e.g., [20]), ecological research should provide essential information regarding biodiversity values, ecological processes, habitat conditions, and the primary threats to wooded grasslands. In Southern Europe, ecological studies have predominantly focused on strictly Mediterranean wooded pastures (e.g., [24,25,26,27]), while research remains extremely scarce in the central-southern Apennines (e.g., [5]), where traditional management strategies such as transhumance [28] have profoundly shaped the landscape. In this region, the term “merigge” is still in use, referring to isolated trees in grasslands that provide shade and shelter [29].
The objectives of the present study were as follows: (i) to provide a floristic and ecological characterization of wooded grasslands in relation to different management intensities; (ii) to evaluate the role of scattered trees in the dynamic processes of afforestation following the abandonment of traditional management practices; and (iii) to define suitable management strategies for the restoration and conservation of this distinctive Apennine grassland landscape.
Given that the structural and ecological features of wooded grasslands differ markedly from those of open grasslands, we have the following hypotheses:
- Different management intensities lead to changes in plant biodiversity along the transect from the tree trunk to the grassland.
- Plant bioindicators of successional dynamics are more abundant under conditions of abandonment compared to managed sites.
- The conservation of wooded grasslands is closely related to the intensity of the application of traditional management practices.
2. Materials and Methods
2.1. Study Area
The study area is located on the flattened upper sectors of the mountainous ridges of Monte San Vicino (Central Apennines) (Figure 1A), which is characterized by calcareous lithotypes. It has altitudes ranging from 1100 to 1200 m a.s.l. and falls within the lower supratemperate thermotype according to the bioclimatic classification proposed by [30,31]. The total mean annual rainfall for the period 1991–2020 is 1153 mm, exhibiting higher precipitation values during autumn and spring and minimum values during summer, while the mean annual temperature is 9.5 °C. The area falls within the Monte San Vicino and Monte Canfaito Regional Natural Reserve, which is included within two Natura 2000 sites (SCI/SAC IT5330015 Monte San Vicino; SPA IT5330025 Monte San Vicino-Monte Canfaito). The vegetation landscape is typically Apennine, characterized by submontane beech forests dominated by Fagus sylvatica (Habitat code 9210* “Apennine beech forests with Taxus and Ilex”), semi-natural grasslands dominated by Bromopsis erecta (Huds.) Fourr. (Habitat code 6210* “Semi-natural dry grasslands and scrubland facies on calcareous substrates (Festuco-Brometalia) (*important orchid sites)”) (Directive 92/42 EEC, Annex I), and wooded grasslands (E7.1 Temperate wooded pasture and meadow), mostly consisting of scattered stumps of beech (Fagus sylvatica L.) that have not been coppiced for at least fifty years.
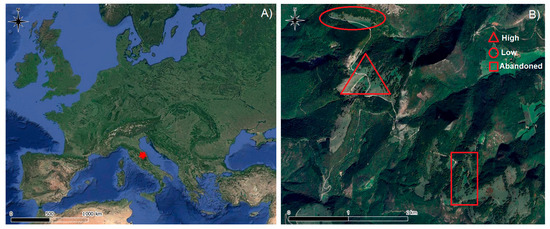
Figure 1.
Location of study area (A) and three wooded grassland sites with different management intensities (B).
2.2. Experimental Design and Surveys
Within the study area, three wooded grassland sites were selected based on varying management intensities: High (mown and grazed), Low (grazed only), and Abandoned (left unmanaged for approximately 50 years) (Figure 1B). Mowing in the High-intensity site occurs annually at the end of June, and grazing is carried out by Marchigiana cattle from mid-September to the end of October (45 days) at a stocking rate of 1.00 Livestock Units (LU) ha−1. In the Low-intensity site, grazing is carried out by Marchigiana cattle from July to the end of October (105 days) at stocking rate of 1.60 LU ha−1.
At each site, four isolated beech (Fagus sylvatica) trees within the grassland were selected, ensuring structural similarity (e.g., crown diameter) and a minimum spacing of 30 m from each other and from the forest edge. Vegetation and floristic composition were surveyed along four transects oriented towards the four cardinal points, employing contiguous 1 m2 plots beginning from the trunk and extending up to 2 m beyond the canopy projection. In each plot, vascular plant species, including seedlings, were listed along with their relative percentage cover. Vascular species identification was conducted following [32], with nomenclature based on the Checklist of the Italian Flora [33]. Each species was additionally associated with an ecological role (nemoral species, herbaceous edge species, forest and pre-forest shrubs, grassland species, and seedlings), in accordance with phytosociological literature [34]. Similarly to the approach adopted by [5], the plots were subsequently grouped into four positions along the 4 transects, and the values were averaged accordingly: “Trunk”, located near the trunk; “Mid-canopy”, positioned halfway between the trunk and the end of the canopy projection; “Edge”, positioned at the farthest part under the canopy, at the end of its projection; and “Grassland”, located in the open grassland two meters beyond the canopy projection (Figure 2). At each position, shrub height and canopy cover (percentage of the sky covered by leaves) were measured together with canopy cover assessed by the mobile application GLAMA [35] with a hemispherical lens positioned 25 cm above the ground to estimate ground-level available light.

Figure 2.
Schematic representation of one radial transect with four sampling areas following the east direction used in the experimental design.
2.3. Statistical Analysis
The Kruskal–Wallis test, designed for non-parametric analyses, was employed to assess the impact of different positions (Trunk, Mid-canopy, Edge, and Grassland) and different management intensities (High, Low, and Abandoned) on the species richness and canopy cover. The Dunn’s post hoc test was applied to compare means. Non-parametric tests were used due to the non-normal distribution of the data.
To analyse the change in species composition in the different positions and management intensities, a principal component analysis (PCA) was performed using the “vegan” package (v. 2.6-8) [36]. All analyses were performed using the software R (v. 4.2.3) [37].
3. Results
3.1. Canopy Cover and Plant Richness
Variations in canopy cover were significantly associated with positions along the transect, following a decreasing gradient from the Trunk position to the Grassland position outside the canopy; this pattern was consistent across all three management intensities (Kruskal–Wallis test, df = 3; p < 0.01 for all three management intensities).
In the Abandoned site, although the trend was similar to that observed in the two managed sites, it is noteworthy that the canopy cover from the Edge to the area outside of the canopy (the Grassland position) was significantly higher (Figure 3A and Table 1).
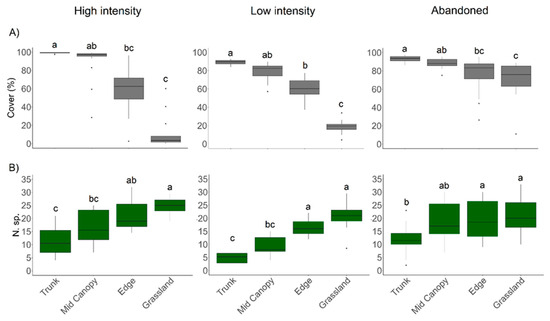
Figure 3.
Mean values of canopy cover (A) and species richness (B) recorded at four positions (Trunk, Mid-canopy, Edge, and Grassland) in each of the three management intensities. Different letters in each graph indicate significant differences (Dunn’s test, p < 0.05).

Table 1.
Mean (±standard deviation) canopy cover, species richness, cover by ecological categories (nemoral, herbaceous edge, shrub forests/pre-forest, grassland, and seedling), and high shrubs in relation to different management intensities (High, Low, or Abandoned) and sampling positions (Trunk, Mid-canopy, Edge, and Grassland). Significant differences among management intensities (Kruskal–Wallis test, p < 0.05) are highlighted in bold. Different letters indicate significant differences among positions (Dunn’s test, p < 0.05).
Species richness was also significantly influenced by the position along the transect, exhibiting a progressive increase from the Trunk to Grassland positions for all the three levels of management intensity (Kruskal–Wallis test, df = 3; p < 0.01 for all three management intensities). This richness trend, clearly evident in the managed sites, remains virtually unchanged from the Mid-canopy to Grassland positions in the Abandoned site (Figure 3B and Table 1).
3.2. Ecological Role of Species
The floristic composition and species abundance, categorized by ecological roles, were significantly influenced by the transect position and management intensity (Table 1 and Figure 4). Nemoral species showed maximum coverage at the Trunk position, decreasing along the transect in all sites. In the managed sites, they were mostly confined under the canopy, with a minimal presence at the Edge, while in the Abandoned site, they remained abundant even in the Grassland. Grassland species displayed the opposite trend, peaking in the Grassland and decreasing toward the Trunk position in all sites. However, their coverage was notably higher in the managed sites, especially under High-intensity management, than in the Abandoned site.
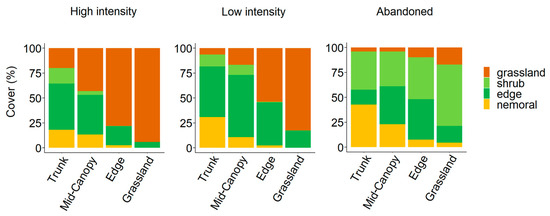
Figure 4.
Percentage cover of species, grouped by ecological categories, across four transect positions for each of the three levels of management intensity.
The herbaceous ecotonal species in the High-intensity site reached their maximum abundance at the Trunk position, progressively decreasing towards the Grassland position, where their abundance was negligible (Table 1). In both the Low-intensity and Abandoned sites, ecotonal species displayed a unimodal distribution, peaking at the Mid-canopy and Edge positions, respectively, with a substantial presence even in the Grassland.
The forest and pre-forest shrub species in the managed sites were mostly under the canopy, peaking at the Trunk position in the High-intensity sites and at the Mid-canopy position in the Low-intensity sites; in the latter, shrubs were scarcely present in the Grassland. Conversely, the shrubs in the Abandoned site were abundant along the transect, increasing from the Trunk to the Grassland position, with maximum coverage at the Grassland position.
The shrub heights (Table 1) mirrored their abundance pattern, with notably greater heights in the Abandoned site, which progressively increased from the Trunk to the Grassland position, surpassing 2 m at the latter. In the managed sites, the shrub heights remained consistently below 10 cm and showed an opposite trend to the Abandoned site, peaking at the Trunk position in the High-intensity site and at the Mid-canopy position in the Low-intensity site.
Finally, the forest and pre-forest seedlings (Table 1) exhibited consistently low and non-significant coverage values across all positions. Their coverage peaked at the Mid-canopy position in all three sites. However, while in the managed sites seedlings occurred exclusively under the canopy, in the Abandoned site, they maintained their presence beyond the canopy.
3.3. Floristic Gradient and Composition
The PCA analysis allowed the evaluation of the floristic composition variation along transect positions for each of the sites characterized by differing management intensities (High, Low, and Abandoned). Along axis 1 (PC1) of the three PCA analyses (Figure 5), from right to left, it was possible to identify a floristic gradient transitioning from the Trunk to the Grassland position, marked by a shift from typically sciaphilous forest species to heliophilous grassland species. This gradient was particularly evident in the two managed sites (Figure 5A,B).
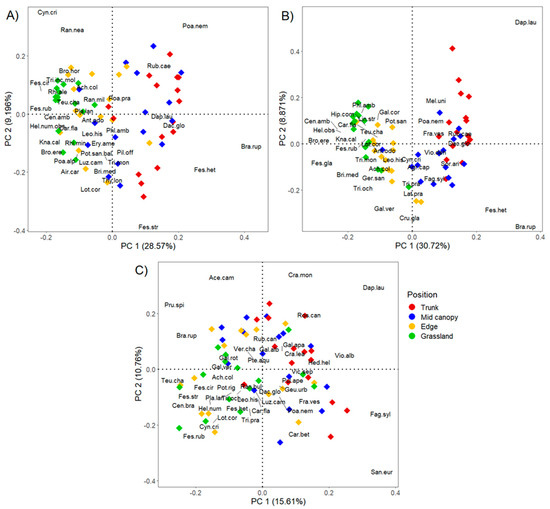
Figure 5.
The Principal Component Analysis (PCA) of the species abundance present in the four positions along the transect for the three areas characterized by different wooded grassland management intensities: High (A), Low (B), and Abandoned (C). Only the species with scores > 0.15 are shown. The abbreviation of the species is listed in the supplementary file (Table S1).
Specifically, in wooded grasslands under High-intensity management (Figure 5A), there was a clear transition from sciaphilous forest species (Daphne laureola L. and Poa nemoralis L.), pre-forest shrub species (Rubus caesius L.), and heliophilous ecotonal edge species (Brachypodium rupestre (Host) Roem. & Schult.) at the Trunk and Mid-canopy positions, to heliophilous grassland species (Centaurea ambigua Guss., Festuca rubra L., B. erecta, Cynosurus cristatus L., and Ranunculus neapolitanus Ten.) at the Edge and Grassland positions.
In wooded grasslands subjected to Low-intensity management (Figure 5B), the transition proceeded from typical sciaphilous forest species (D. laureola, Melica uniflora Retz., and Festuca heterophylla Lam.), herbaceous ecotonal species (B. rupestre), and shrub forest species (R. caesius, F. sylvatica, and Sorbus aria (L.) Crantz) at the Trunk and Mid-canopy positions, to herbaceous ecotonal species (Cruciata glabra (L.) C.Bauhin ex Opiz and Geranium sanguineum L.) at the Edge position, marginally extending into the Grassland position (Trifolium ochroleucon Huds.), and finally to distinctly heliophilous grassland species (C. ambigua, F. rubra, B. erecta, and Knautia calycina (C.Presl) Guss.) at the Grassland position.
In the Abandoned site (Figure 5C), the floristic composition appeared relatively homogeneous across the different positions along the transect. Nevertheless, along the PC1 axis, a subtle floristic gradient could be discerned from left to right, ranging from under-canopy positions (Trunk, Mid-canopy, and Edge), which are characterized by typical sciaphilous forest species (Viola alba subsp. dehnardtii (Ten.) W.Becker, Sanicula europaea L., Pulmonaria vallarsae subsp. apennina (Cristof. & Puppi) L.Cecchi & Selvi, and D. laureola), shrub forest species (F. sylvatica and Carpinus betulus L.), and pre-forest species (Rosa canina L.), to the Grassland position, which hosted herbaceous ecotonal species (B. rupestre), pre-forest shrub species (Prunus spinosa L.), and heliophilous grassland species (C. cristatus, F. rubra, and F. stricta).
4. Discussion
The results highlighted that the amount of filtered light reaching the ground is one of the principal drivers of the floristic gradient observed from the Trunk to Grassland position, influencing the species richness, species ecological category coverage, and overall floristic composition across the three levels of management intensity. Numerous studies have further demonstrated that light availability is among the most critical environmental factors shaping both diversity and abundance patterns within plant communities (e.g., [5,38]).
4.1. Management Intensity and Its Effects on Canopy Cover and Species Richness
In all three sites, the hypothesis of the progressive increase in species richness along the transect from the Trunk to the Grassland position was confirmed. This was associated with an increase in the amount of filtered light reaching the ground and is particularly evident in sites characterized by High- and Low-intensity management. Conversely, in the Abandoned site, this increase is less pronounced, remaining nearly constant from the Mid-canopy to the Grassland position. Unlike in the managed sites, in the Abandoned site, the filtered light reaching the ground at the Edge position is influenced not only by the canopy cover of the isolated tree but also by the shrub cover, while in the Grassland position it is predominantly affected by the shrub cover alone. Consequently, at the Grassland and Edge positions, in addition to a higher canopy cover, the Abandoned site also displays a smaller variation in its canopy cover compared to the managed sites, leading to similar plant richness values across these positions. This increase in the species richness of the herbaceous layer with greater light availability at the ground level has been extensively documented in the literature for temperate forests (e.g., [38,39]). Similar results have also been reported in a recent study on Apennine wooded grasslands [5].
4.2. Management Intensity and Its Effects on Plant Ecological Categories and Floristic Composition
The results of the study support the hypothesis concerning the presence of plant bioindicators of successional dynamics, which are more abundant under abandonment conditions, as evidenced by the occurrence of shrub and ecotonal species outside the canopy area—species that are absent in managed sites. However, while the herbaceous ecotonal species typical of the early stages of vegetation succession are present in the Low-intensity management site, they are lacking in the site subjected to High-intensity management.
The amount of filtered light reaching the ground drives the floristic variation gradient (from sciaphilous to heliophilous species) and determines the presence of four distinct vegetation belts, which are spatially and dynamically interconnected. Each of these belts is characterized by specific ranges of floristic bioindicators. This pattern is particularly evident in the two managed sites, where the light intensity at the ground level is determined exclusively by the canopy of the isolated tree.
Under the canopy of scattered trees, the limited light penetration can significantly suppress the growth and diversity of the herbaceous understory. This competition for light creates a zone beneath the canopy where only a few shade-tolerant species [40] or those adapted to low-light forest conditions (e.g., D. laureola, M. uniflora, and S. europaea) can thrive. As the distance from the tree Trunk increases, the amount of solar radiation reaching the ground also increases, favouring the establishment of more heliophilous edge species (e.g., B. rupestre and C. glabra), similar to the dynamics observed at forest edges that are in contact with managed grasslands [41].
In the Abandoned site, the floristic variation from beneath the canopy to the open grassland is less pronounced, resulting in a more homogeneous floristic composition across positions. Here, shrub cover in the Grassland position outside the canopy generates light conditions comparable to those found beneath the canopy, thus promoting the persistence of shade-tolerant species. This leads to a spatial expansion of the ecotone (the herbaceous edge and pre-forest shrubs) from the Edge into the Grassland position.
As highlighted in various studies, the initial stages of vegetation succession are typically dominated by a few heliophilous edge species, such as B. rupestre, which are particularly effective in dynamically colonizing underused or abandoned grasslands. The expansion of these species causes a significant decline in grassland biodiversity and pastoral value, while simultaneously facilitating the establishment of pre-forest and forest shrub species [42,43,44]. Therefore, the presence and cover of heliophilous edge herbaceous species can serve as reliable indicators not only of the conservation status of secondary grasslands [45] but also of successional processes [46,47], thereby supporting timely restoration and biodiversity conservation efforts.
In the study area, the potential spatio-temporal successional dynamics can be traced from the sites subjected to a high management intensity to the abandoned one by analysing the position and advancement of ecotonal herbaceous species along the transect. In the High-intensity managed sites, these species exhibit peak cover in the Trunk and Mid-canopy positions, are reduced at the Edge, and are nearly absent in the Grassland position, which is dominated by typical heliophilous grassland species (e.g., C. cristatus and B. erecta). Forest shrubs, represented only by R. caesius and not exceeding 10 cm in height, are also restricted to the Trunk and Mid-canopy positions, with maximum cover near the Trunk position.
In the site under Low-intensity management, the early stages of succession are evident. Ecotonal herbaceous species (B. rupestre, G. sanguineum, C. glabra) are more abundant compared to the High-intensity management site and occupy more advanced positions along the transect: B. rupestre dominates the Mid-canopy position, while C. glabra and G. sanguineum are prevalent at the Edge. These species are also well represented in the Grassland position (e.g., T. ochroleucon), which, however, still hosts heliophilous grassland species (B. erecta, B. media and C. ambigua). Forest and pre-forest shrubs (R. caesius, F. sylvatica, and Sorbus aria) are mostly confined to the Trunk and Mid-canopy positions but are also present, albeit with negligible cover and height (<10 cm), at the Edge (0.6%) and in the Grassland (0.1%).
In the Abandoned site, the most advanced successional stage is recorded. Here, both ecotonal herbaceous species (B. rupestre) and forest/pre-forest shrubs (P. spinosa and Acer campestre) occur in the Grassland position, where they reach their highest cover and greatest height (up to 2 m). The presence of shrubs outside the canopy fosters the establishment of shade-tolerant nemoral species (F. heterophylla and Galium rotundifolium L.), which expand from beneath the canopy (Trunk and Mid-canopy position) toward the open grassland, simultaneously inhibiting the occurrence and cover of heliophilous grassland species. Moreover, shrub cover outside the canopy creates microclimatic conditions that are favourable to the establishment and advancement of forest and pre-forest seedlings [17,48]. In the long term, this process may have conflicting implications for the conservation and sustainability of wooded grasslands: on one hand, the regeneration of forest species may contribute to their rejuvenation and resilience in the presence of senescent isolated trees or under overgrazing pressure [2]; on the other hand, as observed in the Abandoned site, the uncontrolled regrowth of trees may accelerate afforestation and canopy closure, ultimately enhancing the senescence and death of the ancient solitary trees [3].
Spatio-temporal vegetation dynamics are initiated in the sites with a low management intensity, beginning from the canopy edge and extending toward the open grassland, driven by the significant presence of heliophilous edge species. If this process is not counteracted, it will inevitably lead to shrub encroachment and grassland colonization, as already observed in the Abandoned site. This suggests that, should grazing cease in the Low-intensity management site, conditions would rapidly favour the afforestation of grasslands starting from the ecotonal edge, mirroring the dynamics observed in the Abandoned site.
4.3. Management Strategies for Wooded Grasslands
As hypothesized, the persistence of wooded grasslands is dependent on the intensity of traditional management practices. The results indicate that High-intensity management, through mowing and subsequent grazing, effectively supports the maintenance of wooded grasslands. In contrast, Low-intensity management of wooded grasslands initiates the natural reforestation process (i.e., expansion of edge species outside the canopy), which proves insufficient for habitat persistence. Approximately 50 years after abandonment, natural succession has led to the dominance of tall shrub vegetation on the open grasslands between isolated trees, resulting in the loss of the habitat (E 7.1 “Temperate wooded pasture and meadow”).
For the conservation of the biodiversity of wooded grasslands and their associated ecosystem services, management should follow traditional practices, as widely recommended in the literature [18,21,22]. In the study area, grazing and mowing (High-intensity management) represent the most appropriate management regime to safeguard biodiversity and related ecosystem services. Under Low-intensity management, occasional but targeted management intensification could be implemented (e.g., increasing instantaneous livestock density, or adopting mowing or chopping). In sites that have been abandoned, the restoration of wooded grasslands may be achieved by removing shrubs outside the canopy, although this approach can entail considerable costs.
Recent efforts to restore scattered trees in grasslands, including interventions within protected areas, represent a promising new approach to conserving the wooded grassland systems of the Apennines.
5. Conclusions
The present study highlights that canopy cover variation, by modulating light availability, is a key driver of the floristic gradient in wooded grasslands. Understanding the vegetation dynamics, particularly the identification of floristic bioindicators during the early stages of succession (e.g., heliophilous herbaceous species) and their distribution along the transect (from the Trunk to the Grassland position), is of paramount importance for planning timely interventions aimed at preserving wooded grasslands.
Plant bioindicators of successional dynamics are more evident under conditions of management abandonment but are also detectable under Low-intensity management, particularly at the canopy edge, where early signs of succession appear. In this regard, High-intensity management proved to be the most effective strategy for maintaining biodiversity and ensuring the persistence of wooded grasslands.
Isolated trees act as centroids of a potential radial succession process that proceeds from the perimeter of the canopy projection toward the grassland (from the Edge to Grassland positions). Initially driven by ecotonal herbaceous vegetation and subsequently by shrubs, this process, if left unchallenged, ultimately leads to the closure of the grassland. Within this framework, the ecotonal zone at the edge of the canopy projection is crucial for the natural reforestation dynamics of grasslands, underscoring the necessity of focusing monitoring activities at this position to ensure the recovery and long-term conservation of wooded grasslands.
Furthermore, within the context of current climate change scenarios, future research could investigate how isolated trees can contribute to mitigating the effects of rising temperatures. They can therefore be interpreted as biodiversity islands, potentially serving as refugia for species that are particularly threatened by climate change. Under extreme conditions, these trees may also function as islands of biomass production, providing forage resources for livestock, as observed in Mediterranean dehesas.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/f16071034/s1, Table S1: Table of floristic surveys.
Author Contributions
Conceptualization, M.A. and G.T.; methodology, M.A. and G.T.; software, G.T.; formal analysis, G.T. and M.F.; investigation, M.A., G.T., M.F., D.G., M.B. and P.D.; data curation, G.T.; writing—original draft preparation, M.A.; writing—review and editing, M.A., G.T., M.F., D.G., M.B. and P.D.; supervision, M.A. and P.D.; funding acquisition, M.A. and P.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research is funded by the PASTINNOVA project “Innovative models for sustainable future of Mediterranean pastoral systems”, which is part of the Partnership for Research and Innovation in the Mediterranean Area (PRIMA) Programme, under grant agreement No. 2113, supported by the European Union’s HORIZON 2020 Research and Innovation Programme and by the MERIDIES project “Innovazioni e strategie condivise per il recupero e la conservazione delle praterie arborate a supporto dei sistemi di allevamento della montagna”, which is part of the Rural Development Programme of Marche Region (Programma di Sviluppo Rurale della Regione Marche 2014/2020—M16.1.A.2—No. 59732).
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Materials. Further inquiries can be directed to the corresponding author.
Acknowledgments
We are grateful to Mattia Di Silvestri, Diego Giulietti, Debora Solforati, Filippo Ronconi, Laura Trozzo, and Silvia Montecchiari for their assistance with the fieldwork. We also thank the Regional Natural Reserve of Monte San Vicino and Monte Canfaito, as well as the Unione Montana Potenza Esino Musone—the managing authority of the protected area—for granting authorization and for providing logistical support and access to the study sites. In particular, we acknowledge Stefania Ramazzotti, Iole Folchi Vici, and Michele Caporaletti of the Unione Montana Potenza Esino Musone for their valuable collaboration. We also express our appreciation to the local farms involved in the MERIDIES project, including Filippo Zenobi and Loris Pierangeli, Francesco and Aldo Lorenzotti, Renzo and Omar Turchi, Maurizio Lanari, and Giovanni Pieraligi. We further thank Marco Giustozzi, who was responsible for drafting the feasibility plan and for technical coordination, and Dimitri Giardini (Marca di Ancona CIA s.r.l.), who coordinated outreach and dissemination activities within the MERIDIES project.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Manning, A.D.; Fischer, J.; Lindenmayer, D.B. Scattered trees are keystone structures—Implications for conservation. Biol. Conserv. 2006, 132, 311–321. [Google Scholar] [CrossRef]
- Bergmeier, E.; Roellig, M. Diversity, threats and conservation of European wood pastures. In European Wood Pastures in Transition: A Social-Ecological Approach, 1st ed.; Hartel, T., Plieninger, T., Eds.; Routledge: Abingdon, UK, 2014; Chapter 2. [Google Scholar]
- Garbarino, M.; Bergmeier, E. Plant and vegetation diversity in European wood-pastures. In European Wood Pastures in Transition: A Social-Ecological Approach, 1st ed.; Hartel, T., Plieninger, T., Eds.; Routledge: Abingdon, UK, 2014; Chapter 7. [Google Scholar]
- Seddaiu, G.; Bagella, S.; Pulina, A.; Cappai, C.; Salis, L.; Rossetti, I.; Lai, R.; Roggero, P.P. Mediterranean cork oak wood grasslands: Synergies and trade-offs between plant diversity, pasture production and soil carbon. Agrofor. Syst. 2018, 92, 893–908. [Google Scholar] [CrossRef]
- Tesei, G.; Bonanomi, G.; D’Ottavio, P.; Francioni, M.; Giovagnoli, D.; Landi, L.; Murolo, S.; Trozzo, L.; Allegrezza, M. Under the Crown of Scattered Beech (Fagus sylvatica): Impact on Plant Community, Soil Resources, and Fungal Diversity. J. Veg. Sci. 2025, 36, e70021. [Google Scholar] [CrossRef]
- Hartel, T.; Dorresteijn, I.; Klein, C.; Máthé, O.; Moga, C.I.; Öllerer, K.; Roellig, M.; von Wehrden, H.; Fischer, J. Wood-pastures in a traditional rural region of Eastern Europe: Characteristics, management and status. Biol. Conserv. 2013, 166, 267–275. [Google Scholar] [CrossRef]
- Plieninger, T.; Hartel, T.; Martín-López, B.; Beaufoy, G.; Bergmeier, E.; Kirby, K.; Montero, M.J.; Moreno, G.; Oteros-Rozas, E.; Van Uytvanck, J. Wood-pastures of Europe: Geographic coverage, social–ecological values, conservation management, and policy implications. Biol. Conserv. 2015, 190, 70–79. [Google Scholar] [CrossRef]
- Vojta, J.; Drhovska, L. Are abandoned wooded pastures suitable refugia for forest species? J. Veg. Sci. 2012, 23, 880–891. [Google Scholar] [CrossRef]
- Bergmeier, E.; Petermann, J.; Schröder, E. Geobotanical survey of wood-pasture habitats in Europe: Diversity, threats and conservation. Biodivers. Conserv. 2010, 19, 2995–3014. [Google Scholar] [CrossRef]
- Centeri, C.; Renes, H.; Roth, M.; Kruse, A.; Eiter, S.; Kapfer, J.; Santoro, A.; Agnoletti, M.; Emanueli, F.; Sigura, M.; et al. Wooded Grasslands as Part of the European Agricultural Heritage. In Biocultural Diversity in Europe Environmental History; Agnoletti, M., Emanueli, F., Eds.; Springer: Cham, Switzerland, 2016; Volume 5, pp. 75–103. [Google Scholar] [CrossRef]
- Mosquera-Losada, M.R.; McAdam, J.H.; Romero-Franco, R. Definitions and components of agroforestry practices in Europe. In Agroforestry in Europe. Current Status and Future Prospects; Rigueiro-Rodríguez, A., McAdam, J., Mosquera-Losada, M.R., Eds.; Springer: Berlin, Germany, 2009; pp. 3–19. [Google Scholar]
- Bianchini, M.; Tarhouni, M.; Francioni, M.; Fiorentini, M.; Rivosecchi, C.; Msadek, J.; Tlili, A.; Chouikhi, F.; Allegrezza, M.; Tesei, G.; et al. Modeling Climate-Driven Vegetation Changes Under Contrasting Temperate and Arid Conditions in the Mediterranean Basin. Ecol. Evol. 2025, 15, e70753. [Google Scholar] [CrossRef]
- Pardini, A.; Nori, M. Agro-silvo-pastoral systems in Italy: Integration and diversification. Pastor. Res. Policy Pract. 2011, 1, 26. [Google Scholar] [CrossRef]
- Costa, A.; Madeira, M.; Santos, J.L.; Plieninger, T. Recent dynamics of evergreen oak wood-pastures in south-western Iberia. In European Wood Pastures in Transition: A Social-Ecological Approach, 1st ed.; Hartel, T., Plieninger, T., Eds.; Routledge: Abingdon, UK, 2014; Chapter 5. [Google Scholar]
- Garrido, P.; Elbakidze, M.; Angelstam, P. Stakeholders’ perceptions on ecosystem services in Östergötland’s (Sweden) threatened oak wood-pasture landscapes. Landsc. Urban Plan. 2017, 158, 96–104. [Google Scholar] [CrossRef]
- Dean, G.; Francioni, M.; Toderi, M.; Lopez-i-Gelats, F.; Trozzo, L.; Rivera-Ferre, M.G.; Franca, A.; Altana, G.M.; Karatassiou, M.; Parissi, Z.M.; et al. Nature’s contribution to people provided by pastoral systems across European, African, and Middle East Mediterranean countries: Trends, approaches and gaps. Reg. Environ. Change 2024, 24, 77. [Google Scholar] [CrossRef]
- Garbarino, M.; Morresi, D.; Urbinati, C.; Malandra, F.; Motta, R.; Sibona, E.M.; Vitali, A.; Weisberg, P.J. Contrasting land use legacy effects on forest landscape dynamics in the Italian Alps and the Apennines. Landsc. Ecol. 2020, 35, 2679–2694. [Google Scholar] [CrossRef]
- Garbarino, M.; Lingua, E.; Subira, M.M.; Motta, R. The larch wood pasture: Structure and dynamics of a cultural landscape. Eur. J. For. Res. 2011, 130, 491–502. [Google Scholar] [CrossRef]
- Jørgensen, D.; Quelch, P. The origins and history of medieval wood-pastures. In European Wood Pastures in Transition: A Social-Ecological Approach, 1st ed.; Hartel, T., Plieninger, T., Eds.; Routledge: Abingdon, UK, 2014; Chapter 4. [Google Scholar]
- Janssen, J.A.M.; Rodwell, J.S.; García Criado, M.; Gubbay, S.; Haynes, T.; Nieto, A.; Sanders, N.; Landucci, F.; Loidi, J.; Symank, A.; et al. (Eds.) European Red List of Habitats. Part 2. Terrestrial and Freshwater Habitats; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar] [CrossRef]
- Plieninger, T.; Dijks, S.; Oteros-Rozas, E.; Bieling, C. Assessing, mapping, and quantifying cultural ecosystem services at community level. Land Use Policy 2013, 33, 118–129. [Google Scholar] [CrossRef]
- Varga, A.; Zsolt, M. The role of traditional ecological knowledge in managing wood-pastures. In European Wood Pastures in Transition: A Social-Ecological Approach, 1st ed.; Hartel, T., Plieninger, T., Eds.; Routledge: Abingdon, UK, 2014; Chapter 11. [Google Scholar]
- Plieninger, T.; Hartel, T. European wood-pastures in transition: Lessons for science, conservation and policy development in high nature value landscapes. In European Wood Pastures in Transition: A Social-Ecological Approach, 1st ed.; Hartel, T., Plieninger, T., Eds.; Routledge: Abingdon, UK, 2014; Chapter 17. [Google Scholar]
- Alias, S.; Bianchi, L.; Calamini, G.; Gregori, E.; Sioni, S. Shrub facilitation of Quercus ilex and Quercus pubescens regeneration in a wooded pasture in central Sardinia (Italy). iForest Biogeosci. For. 2010, 3, 16–22. [Google Scholar] [CrossRef]
- Rossetti, I.; Bagella, S.; Cappai, C.; Caria, M.; Lai, R.; Roggero, P.; da Silva, P.M.; Sousa, J.; Querner, P.; Seddaiu, G. Isolated cork oak trees affect soil properties and biodiversity in Mediterranean wooded grassland. Agric. Ecosyst. Environ. 2015, 202, 203–216. [Google Scholar] [CrossRef]
- Moreno, G.; Gonzalez-Bornay, G.; Pulido, F.; Lopez-Diaz, M.L.; Bertomeu, M.; Juárez, E.; Diaz, M. Exploring the causes of high biodiversity of Iberian dehesas: The importance of wood pastures and marginal habitats. Agrofor. Syst. 2016, 90, 87–105. [Google Scholar] [CrossRef]
- Bagella, S.; Caria, M.C.; Seddaiu, G.; Leites, L.; Roggero, P.P. Patchy landscapes support more plant diversity and ecosystem services than wood grasslands in Mediterranean silvopastoral agroforestry systems. Agric. Syst. 2020, 185, 102945. [Google Scholar] [CrossRef]
- Susmel, P.; Fabro, C.; Filacorda, S. Transhumance in the Italian Alps and Apennines. In Transhumance and Biodiversity in European Mountains, 1st ed.; Bunce, G.H., Pérez-Soba, M., Jongman, R.H.G., Sal, A.G., Herzog, F., Eds.; IALE: Wageningen, The Netherlands, 2004; pp. 231–232. [Google Scholar]
- Alinei, M. (Ed.) From Lat. meridies ‘Sheep’s Meriggio’, to Lat. mora and Lat. umbra: Italic Origins and Linguistic Development of a Term from Transhumant Pastoralism; Quaderni di Semantica, a. XXX, 1; Cooperativa Libraria Universitaria CLUEB: Bologna, Italy, 2009. (In Italian) [Google Scholar]
- Rivas-Martínez, S.; Sáenz, S.R.; Penas, À. Worldwide bioclimatic classification system. Glob. Geobot. 2011, 1, 1–634. [Google Scholar] [CrossRef]
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Pignatti, S.; Guarino, R.; La Rosa, M. (Eds.) Flora of Italy, 2nd ed.; Edagricole New Business Media: Milano, Italy, 2017. (In Italian) [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. 2014, 148, 728–814. [Google Scholar] [CrossRef]
- Tichý, L. GLAMA-Gap Light Analysis Mobile Application; Department of Botany and Zoology: Brno, Czech Republic, 2014. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 January 2025).
- R Core Team. R: A Language and Environment for Statistical Computing. Version 4.2.2. 2021. Available online: https://www.R-project.org/ (accessed on 10 November 2024).
- Dormann, C.F.; Bagnara, M.; Boch, S.; Hinderling, J.; Janeiro-Otero, A.; Schäfer, D.; Schall, P.; Hartig, F. Plant species richness increases with light availability, but not variability, in temperate forests understorey. BMC Ecol. 2020, 20, 43. [Google Scholar] [CrossRef] [PubMed]
- Márialigeti, S.; Tinya, F.; Bidló, A.; Ódor, P. Environmental drivers of the composition and diversity of the herb layer in mixed temperate forests in Hungary. Plant Ecol. 2016, 217, 549–563. [Google Scholar] [CrossRef]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Biondicazione attraverso le piante vascolari. Valori di indicazione secondo Ellenberg (Zeigerwerte) per le specie della Flora d’Italia. Braun-Blanquetia 2005, 39, 1–97. [Google Scholar]
- De Pauw, K.; Meeussen, C.; Govaert, S.; Sanczuk, P.; Vanneste, T.; Bernhardt-Römermann, M.; Bollmann, K.; Brunet, J.; Calders, K.; Cousins, S.A.O.; et al. Taxonomic, phylogenetic and functional diversity of understorey plants respond differently to environmental conditions in European forest edges. J. Ecol. 2021, 109, 2629–2648. [Google Scholar] [CrossRef]
- Allegrezza, M.; Biondi, E.; Ballelli, S.; Tesei, G.; Ottaviani, C. The edge communities of Asphodelus macrocarpus subsp. macrocarpus: The different ecological aspects and a new case study in the central Apennines. Plant Sociol. 2015, 52, 19–40. [Google Scholar] [CrossRef]
- Allegrezza, M.; Biondi, E.; Ballelli, S.; Tesei, G.; Ottaviani, C.; Zitti, S. Brachypodium rupestre (Host) Roem. & Schult. Herbaceous communities of heliophilous edge in the Trifolio medii-Geranietea sanguinei Müller 1962 class. Plant Sociol. 2016, 53, 59–76. [Google Scholar] [CrossRef]
- Tesei, G.; D’oTtavio, P.; Toderi, M.; Ottaviani, C.; Pesaresi, S.; Francioni, M.; Trozzo, L.; Allegrezza, M. Restoration strategies for grasslands colonized by asphodel-dominant communities. Grassl. Sci. 2020, 66, 54–63. [Google Scholar] [CrossRef]
- Angelini, P.; Casella, L.; Grignetti, A.; Genovesi, P. (Eds.) Manuali per Il Monitoraggio di Specie e Habitat di Interesse Comunitario (Direttiva 92/43/CEE) in Italia: Habitat; Serie Manuali e Linee Guida, 142/2016; ISPRA: Roma, Italy, 2016. [Google Scholar]
- Biondi, E.; Allegrezza, M.; Casavecchia, S.; Galdenzi, D.; Gasparri, R.; Pesaresi, S.; Soriano, P.; Tesei, G.; Blasi, C. New insight on Mediterranean and sub-Mediterranean syntaxa included in the Vegetation Prodrome of Italy. Flora Mediterr. 2015, 25, 77–102. [Google Scholar] [CrossRef]
- De Simone, W.; Allegrezza, M.; Frattaroli, A.R.; Montecchiari, S.; Tesei, G.; Zuccarello, V.; Di Musciano, M. From Remote Sensing to Species Distribution Modelling: An Integrated Workflow to Monitor Spreading Species in Key Grassland Habitats. Remote Sens. 2021, 13, 1904. [Google Scholar] [CrossRef]
- Holl, K.D. Effect of shrubs on tree seedling establishment in an abandoned tropical pasture. J. Ecol. 2002, 90, 179–187. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).