
Dynamics and Structural Changes in the Janj Mixed Old-Growth Mountain Forest: Continuing Decline of Conifers



Abstract
1. Introduction
2. Materials and Methods
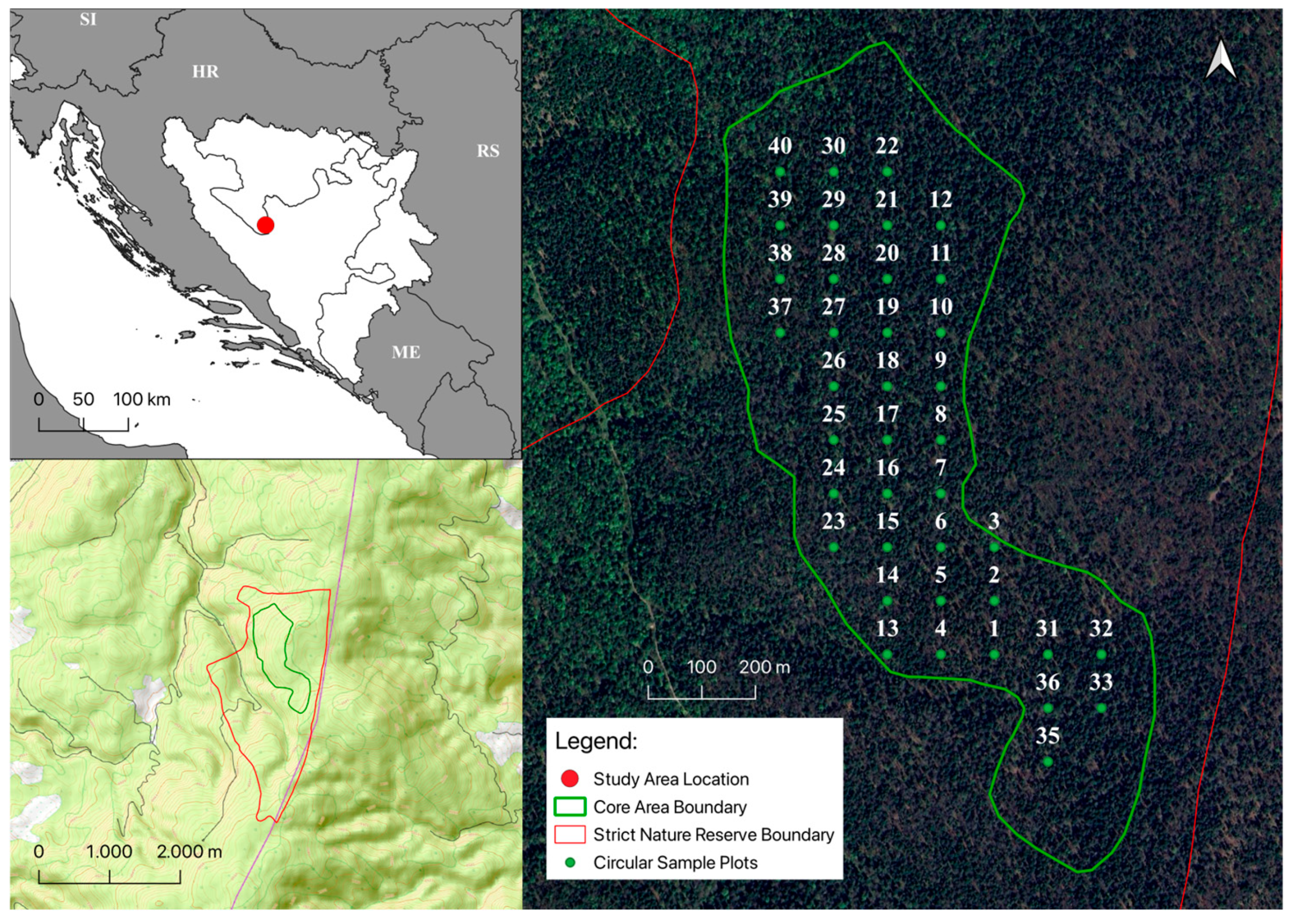
2.1. Study Site
2.2. Field Measurements
2.3. Data Analysis
3. Results
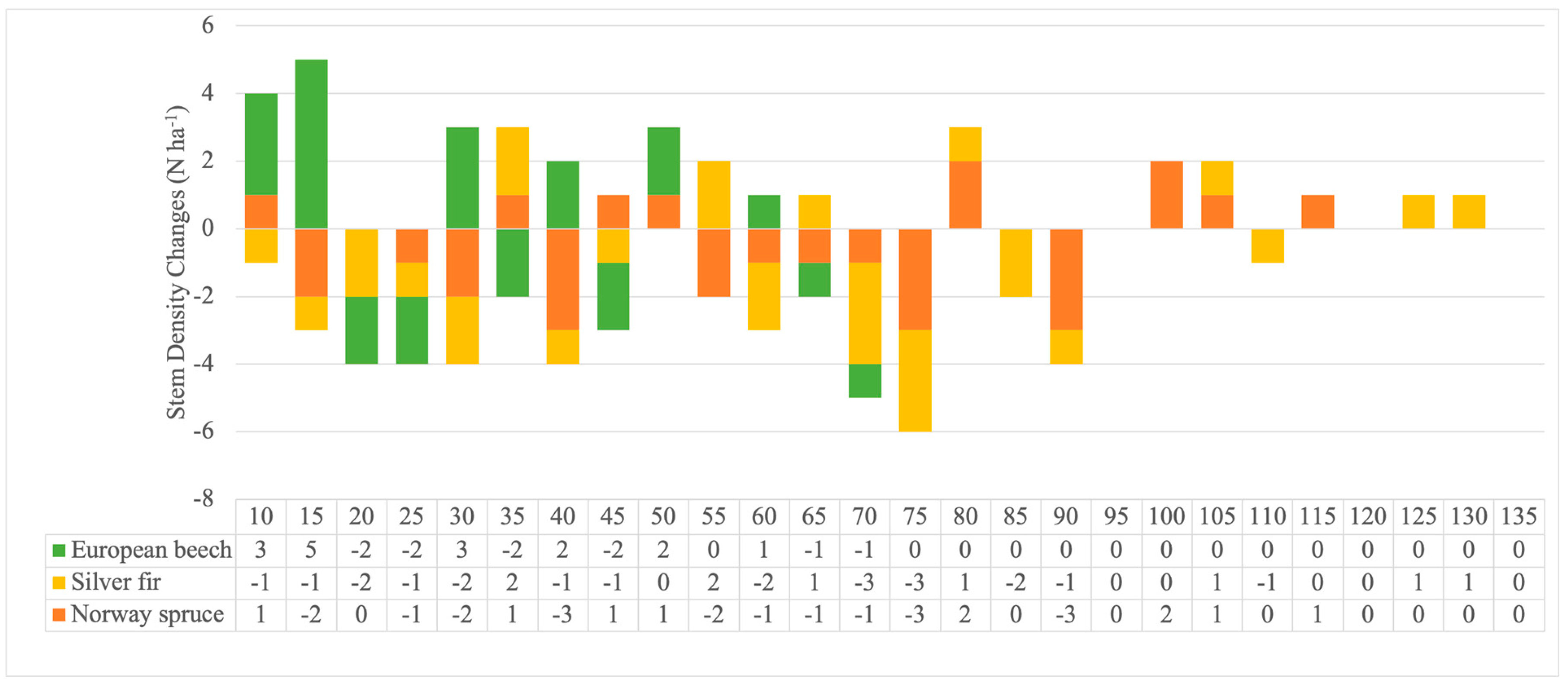
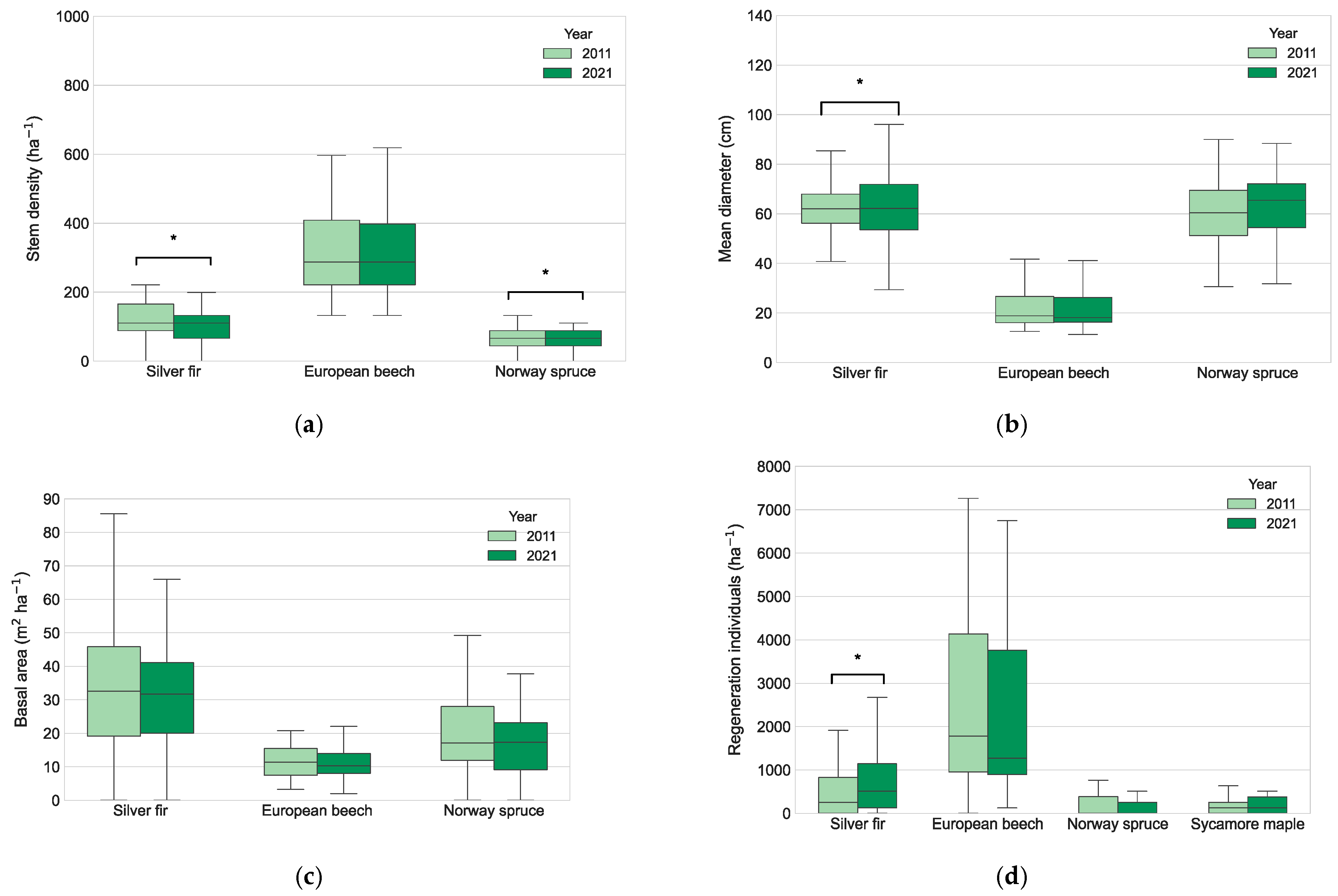
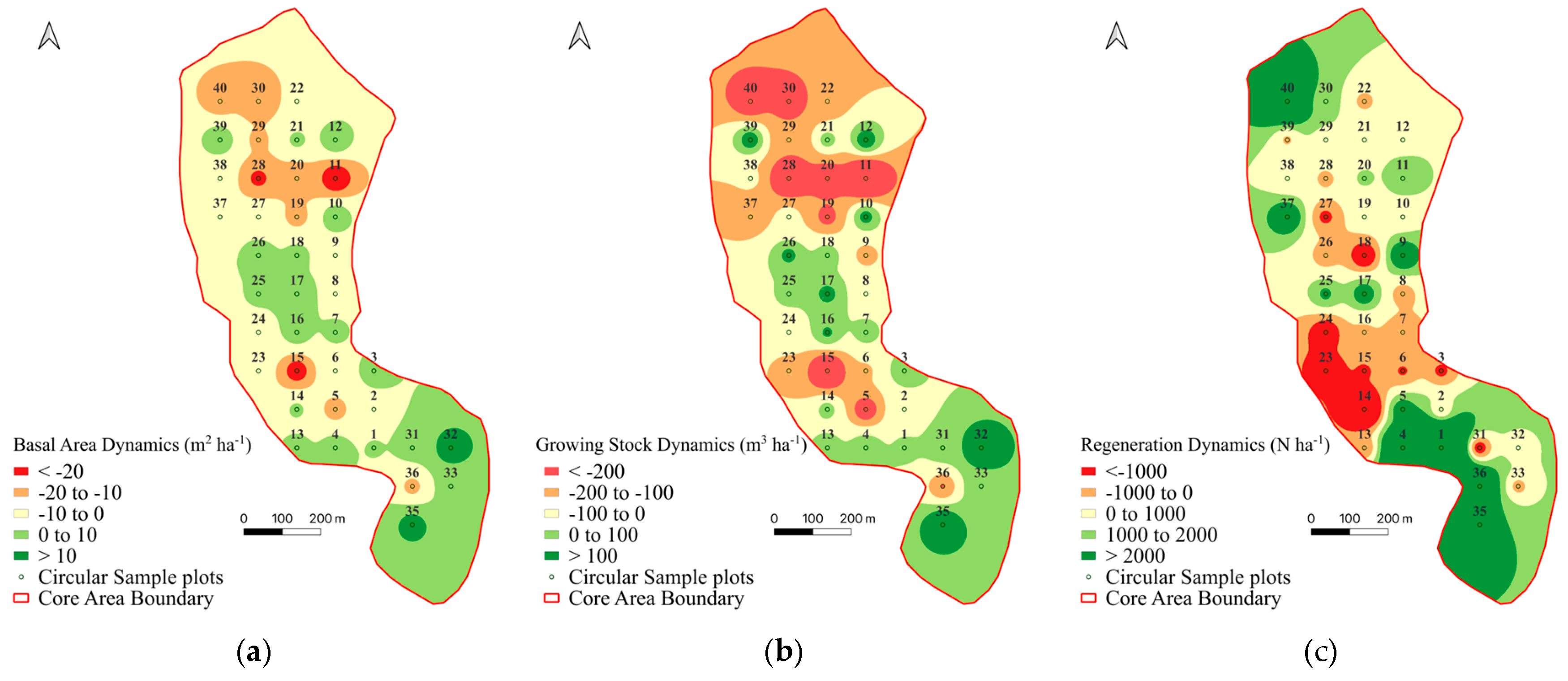
3.1. Forest Structure Changes and Regeneration
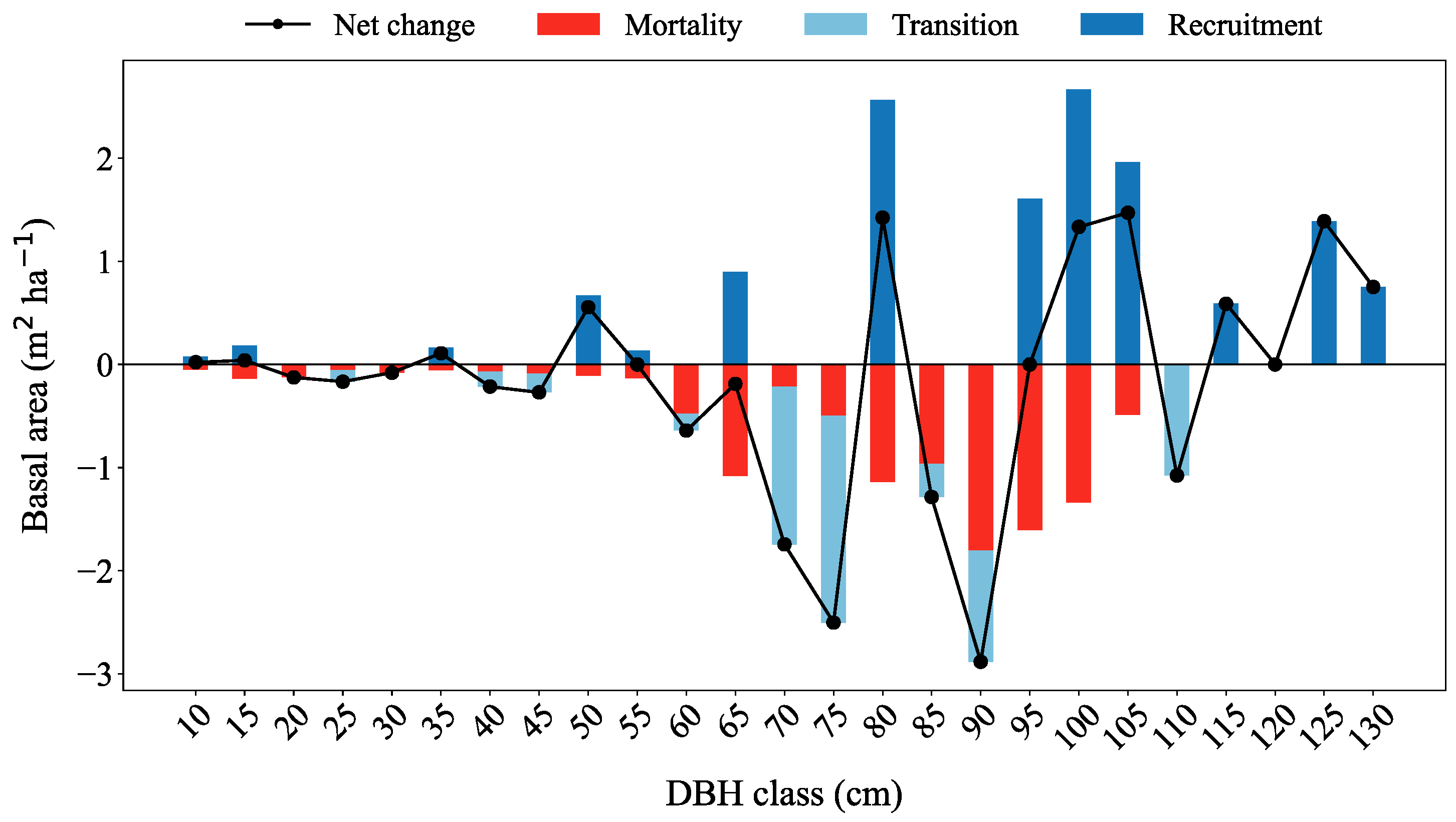
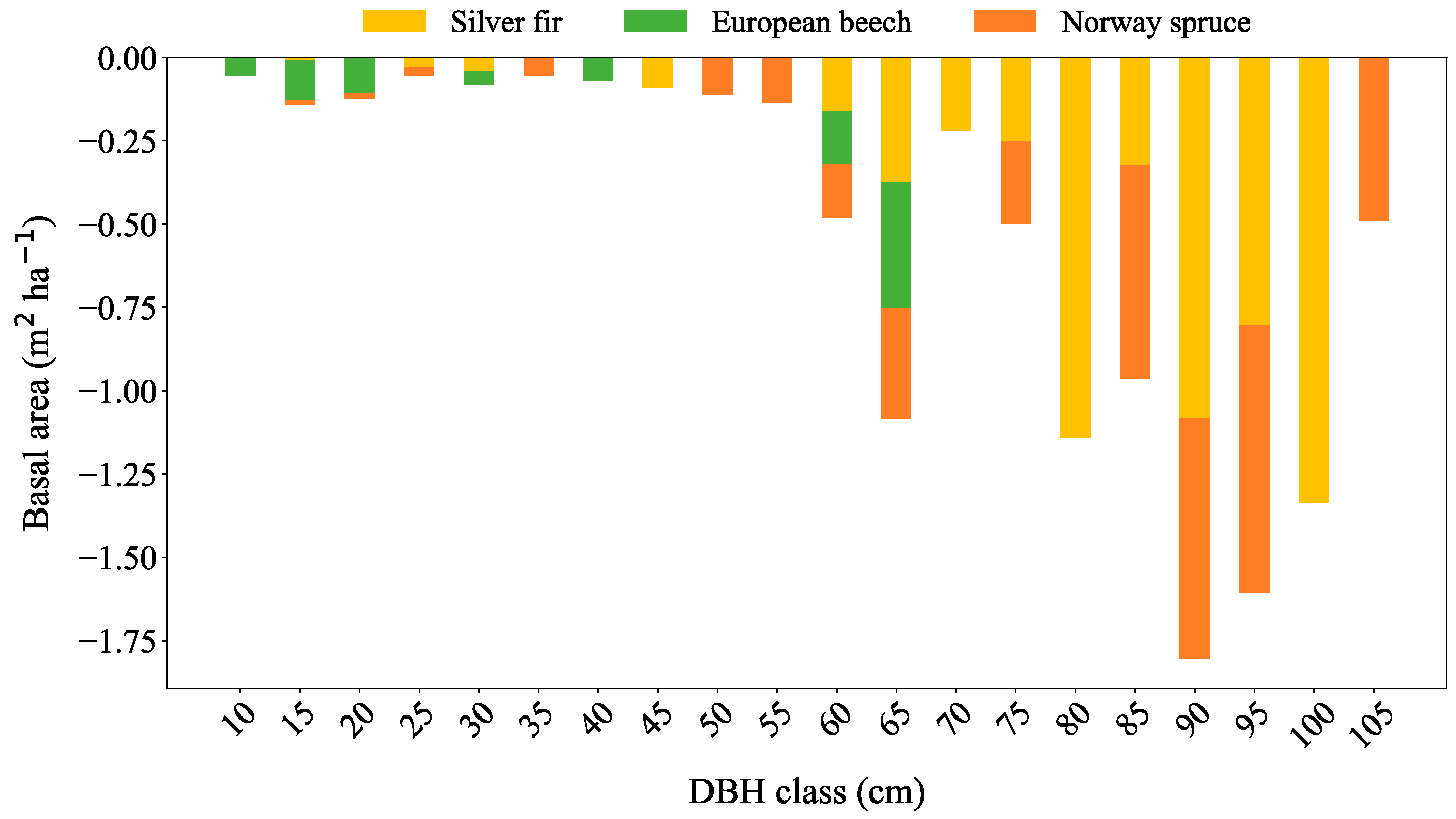
3.2. Recruitment, Transition and Mortality
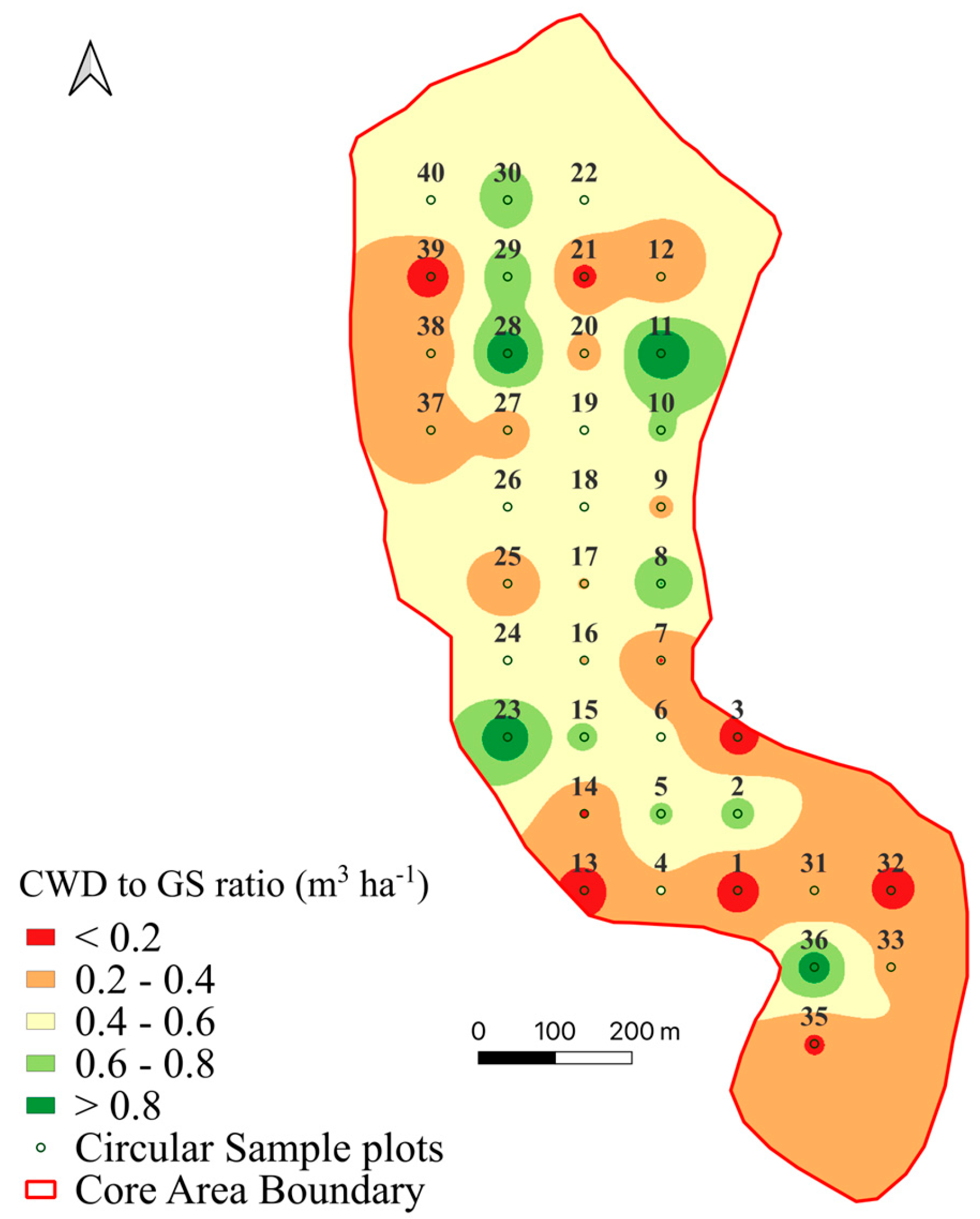
3.3. Coarse Woody Debris
4. Discussion
4.1. Structural Changes
4.2. Developmental Dynamics
4.3. Mortality Patterns
4.4. Tree Species Composition and Diameter Distribution
4.5. Implications for Regeneration Dynamics
4.6. Broader Ecological and Management Implications
4.7. Study Limitations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BA | Basal Area |
| CWD | Coarse Woody Debris |
| DBH | Diameter at breast height measured in centimeters |
| GS | Growing Stock |
| SNR | Strict Nature Reserve |
References
- Sabatini, F.M.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where Are Europe’s Last Primary Forests? Divers. Distrib. 2018, 24, 1426–1439. [Google Scholar] [CrossRef]
- Stubenrauch, J.; Garske, B. Forest Protection in the EU’s Renewable Energy Directive and Nature Conservation Legislation in Light of the Climate and Biodiversity Crisis—Identifying Legal Shortcomings and Solutions. For. Policy Econ. 2023, 153, 102996. [Google Scholar] [CrossRef]
- Barredo, J.I.; Brailescu, C.; Teller, A.; Sabatini, F.M.; Mauri, A.; Janouskova, K. Mapping and Assessment of Primary and Old-Growth Forests in Europe; Publications Office of the European Union: Luxembourg, 2021; ISBN 978-92-76-34230-4. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M.; Kobal, M. Disturbance, Life History Traits, and Dynamics in an Old-Growth Forest Landscape of Southeastern Europe. Ecol. Appl. 2014, 24, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Forest Europe. State of Europe’s Forests 2020; Köhl, M., Linser, S., Prins, K., Eds.; Forest Europe: Jönköping, Sweden, 2020. [Google Scholar]
- Čilaš, M.; Leiter, M.; Višnjić, Ć.; Hasenauer, H. Adapting the Tree Growth Model MOSES to Manage Uneven-Aged Mixed Species Forests in Bosnia & Herzegovina. Trees For. People 2023, 14, 100433. [Google Scholar] [CrossRef]
- O’hara, K.L.; Hasenauer, H.; Kindermann, G. Sustainability in Multi-Aged Stands: An Analysis of Long-Term Plenter Systems. Forestry 2007, 80, 163–181. [Google Scholar] [CrossRef]
- Leiter, M.; Hasenauer, H. Continuous Cover Forestry: Which Sampling Method Should Be Used to Ensure Sustainable Management? Trees For. People 2023, 13, 100419. [Google Scholar] [CrossRef]
- Nagel, T.A.; Cerioni, M. Structure and Dynamics of Old-Growth Pinus Nigra Stands in Southeast Europe. Eur. J. For. Res. 2023, 142, 537–545. [Google Scholar] [CrossRef]
- Cagliero, E.; Morresi, D.; Paradis, L.; Čurović, M.; Spalevic, V.; Marchi, N.; Meloni, F.; Bentaleb, I.; Motta, R.; Garbarino, M.; et al. Legacies of Past Human Activities on One of the Largest Old-Growth Forests in the South-East European Mountains. Veg. Hist. Archaeobotany 2022, 31, 415–430. [Google Scholar] [CrossRef]
- Nagel, T.A.; Mikac, S.; Dolinar, M.; Klopcic, M.; Keren, S.; Svoboda, M.; Diaci, J.; Bončina, A.; Paulic, V. The Natural Disturbance Regime in Forests of the Dinaric Mountains: A Synthesis of Evidence. For. Ecol. Manag. 2017, 388, 29–42. [Google Scholar] [CrossRef]
- Paluch, J.; Keren, S.; Govedar, Z. The Dinaric Mountains versus the Western Carpathians: Is Structural Heterogeneity Similar in Close-to-Primeval Abies–Picea–Fagus Forests? Eur. J. For. Res. 2021, 140, 209–225. [Google Scholar] [CrossRef]
- Chen, B.; Wang, Y.-C. Carbon Storage in Old-Growth Homestead Windbreaks of Small Islands in Okinawa: Toward the Sustainable Management and Conservation. Forests 2020, 11, 448. [Google Scholar] [CrossRef]
- Remeš, J. Development and Present State of Close-to-Nature Silviculture. J. Landsc. Ecol. 2018, 11, 17–32. [Google Scholar] [CrossRef]
- Schütz, J.-P.; Saniga, M.; Diaci, J.; Vrška, T. Comparing Close-to-Nature Silviculture with Processes in Pristine Forests: Lessons from Central Europe. Ann. For. Sci. 2016, 73, 911–921. [Google Scholar] [CrossRef]
- Bugmann, H.; Brang, P. Ausgewählte Ergebnisse Aus Fünfzig Jahren Forschung in Schweizer Naturwaldreservaten. Forum Für Wissen WSL 2009, 2009, 93–102. [Google Scholar]
- Chiti, T.; Benilli, N.; Mastrolonardo, G.; Certini, G. The Potential for an Old-Growth Forest to Store Carbon in the Topsoil: A Case Study at Sasso Fratino, Italy. J. For. Res. 2023, 35, 10. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, A.; Liu, Y.; Shen, L.; Cai, R.; Wu, J. Disturbance of Wind Damage and Insect Outbreaks in the Old-Growth Forest of Changbai Mountain, Northeast China. Forests 2023, 14, 368. [Google Scholar] [CrossRef]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for Old-Growth Attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef]
- Nagel, T.A.; Firm, D.; Rozman, A. Intermediate Disturbances Are a Key Driver of Long-Term Tree Demography Across Old-growth Temperate Forests. Ecol. Evol. 2021, 11, 16862–16873. [Google Scholar] [CrossRef]
- Boncina, A. Comparison of Structure and Biodiversity in the Rajhenav Virgin Forest Remnant and Managed Forest in the Dinaric Region of Slovenia: Structure and Biodiversity of Forests in Slovenia. Glob. Ecol. Biogeogr. 2000, 9, 201–211. [Google Scholar] [CrossRef]
- Frankovič, M.; Janda, P.; Mikoláš, M.; Čada, V.; Kozák, D.; Pettit, J.L.; Nagel, T.A.; Buechling, A.; Matula, R.; Trotsiuk, V.; et al. Natural Dynamics of Temperate Mountain Beech-Dominated Primary Forests in Central Europe. For. Ecol. Manag. 2021, 479, 118522. [Google Scholar] [CrossRef]
- Senf, C.; Sebald, J.; Seidl, R. Increasing Canopy Mortality Affects the Future Demographic Structure of Europe’s Forests. One Earth 2021, 4, 749–755. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Miller, A.D.; Mohan, J.E.; Hudiburg, T.W.; Duval, B.D.; DeLucia, E.H. Altered Dynamics of Forest Recovery under a Changing Climate. Glob. Change Biol. 2013, 19, 2001–2021. [Google Scholar] [CrossRef] [PubMed]
- Luyssaert, S.; Schulze, E.-D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-Growth Forests as Global Carbon Sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Liu, S.; Li, Z.; Zhang, D.; Tang, X.; Zhou, C.; Yan, J.; Mo, J. Old-Growth Forests Can Accumulate Carbon in Soils. Science 2006, 314, 1417. [Google Scholar] [CrossRef]
- Woods, K.D.; Nagel, T.A.; Brzeziecki, B.; Cowell, C.M.; Firm, D.; Jaloviar, P.; Kucbel, S.; Lin, Y.; Maciejewski, Z.; Szwagrzyk, J.; et al. Multi-Decade Tree Mortality in Temperate Old-Growth Forests of Europe and North America: Non-Equilibrial Dynamics and Species-Individualistic Response to Disturbance. Glob. Ecol. Biogeogr. 2021, 30, 1311–1333. [Google Scholar] [CrossRef]
- Diaci, J.; Roženbergar, D.; Anić, I.; Mikac, S.; Saniga, M.; Kucbel, S.; Višnjić, C.; Ballian, D. Structural Dynamics and Synchronous Silver Fir Decline in Mixed Old-Growth Mountain Forests in Eastern and Southeastern Europe. Forestry 2011, 84, 479–491. [Google Scholar] [CrossRef]
- Firm, D.; Nagel, T.A.; Diaci, J. Disturbance History and Dynamics of an Old-Growth Mixed Species Mountain Forest in the Slovenian Alps. For. Ecol. Manag. 2009, 257, 1893–1901. [Google Scholar] [CrossRef]
- Jaloviar, P.; Saniga, M.; Kucbel, S.; Pittner, J.; Vencurik, J.; Dovciak, M. Seven Decades of Change in a European Old-Growth Forest Following a Stand-Replacing Wind Disturbance: A Long-Term Case Study. For. Ecol. Manag. 2017, 399, 197–205. [Google Scholar] [CrossRef]
- Parobeková, Z.; Pittner, J.; Kucbel, S.; Saniga, M.; Filípek, M.; Sedmáková, D.; Vencurik, J.; Jaloviar, P. Structural Diversity in a Mixed Spruce-Fir-Beech Old-Growth Forest Remnant of the Western Carpathians. Forests 2018, 9, 379. [Google Scholar] [CrossRef]
- Šamonil, P.; Doleželová, P.; Vašíčková, I.; Adam, D.; Valtera, M.; Král, K.; Janík, D.; Šebková, B. Individual-Based Approach to the Detection of Disturbance History through Spatial Scales in a Natural Beech-Dominated Forest. J. Veg. Sci. 2013, 24, 1167–1184. [Google Scholar] [CrossRef]
- Vrška, T.; Adam, D.; Hort, L.; Kolář, T.; Janík, D. European Beech (Fagus sylvatica L.) and Silver Fir (Abies alba Mill.) Rotation in the Carpathians—A Developmental Cycle or a Linear Trend Induced by Man? For. Ecol. Manag. 2009, 258, 347–356. [Google Scholar] [CrossRef]
- Diaci, J.; Roženbergar, D.; Mikac, S.; Anić, I.; Hartman, T.; Bončina, A. Long-Term Changes in Tree Species Composition in Old-Growth Dinaric Beech-Fir Forest. Glas. Za Šumske Pokuse 2007, 42, 13–28. [Google Scholar]
- Keren, S.; Motta, R.; Govedar, Z.; Lučić, R.; Medarević, M.; Diaci, J. Comparative Structural Dynamics of the Janj Mixed Old-Growth Mountain Forest in Bosnia and Herzegovina: Are Conifers in a Long-Term Decline? Forests 2014, 5, 1243–1266. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M.; Rugani, T.; Diaci, J. Gap Regeneration and Replacement Patterns in an Old-Growth Fagus–Abies Forest of Bosnia–Herzegovina. Plant Ecol. 2010, 208, 307–318. [Google Scholar] [CrossRef]
- Čater, M. Response and Mortality of Beech, Fir, Spruce and Sycamore to Rapid Light Exposure after Large-Scale Disturbance. For. Ecol. Manag. 2021, 498, 119554. [Google Scholar] [CrossRef]
- Keren, S.; Diaci, J.; Motta, R.; Govedar, Z. Stand Structural Complexity of Mixed Old-Growth and Adjacent Selection Forests in the Dinaric Mountains of Bosnia and Herzegovina. For. Ecol. Manag. 2017, 400, 531–541. [Google Scholar] [CrossRef]
- Ammer, C. Impact of Ungulates on Structure and Dynamics of Natural Regeneration of Mixed Mountain Forests in the Bavarian Alps. For. Ecol. Manag. 1996, 88, 43–53. [Google Scholar] [CrossRef]
- Roženbergar, D.; Mikac, S.; Anić, I.; Diaci, J. Gap Regeneration Patterns in Relationship to Light Heterogeneity in Two Old-Growth Beech–Fir Forest Reserves in South East Europe. For. Int. J. For. Res. 2007, 80, 431–443. [Google Scholar] [CrossRef]
- Ficko, A.; Roessiger, J.; Bončina, A. Can the Use of Continuous Cover Forestry Alone Maintain Silver Fir (Abies alba Mill.) in Central European Mountain Forests? For. Int. J. For. Res. 2016, 89, 412–421. [Google Scholar] [CrossRef]
- Vacek, Z. Structure and Dynamics of Spruce-Beech-Fir Forests in Nature Reserves of the Orlické Hory Mts. in Relation to Ungulate Game. Cent. Eur. For. J. 2017, 63, 23–34. [Google Scholar] [CrossRef]
- Bernard, M.; Boulanger, V.; Dupouey, J.-L.; Laurent, L.; Montpied, P.; Morin, X.; Picard, J.-F.; Saïd, S. Deer Browsing Promotes Norway Spruce at the Expense of Silver Fir in the Forest Regeneration Phase. For. Ecol. Manag. 2017, 400, 269–277. [Google Scholar] [CrossRef]
- Diaci, J.; Adamic, T.; Fidej, G.; Rozenbergar, D. Toward a Beech-Dominated Alternative Stable State in Dinaric Mixed Montane Forests: A Long-Term Study of the Pecka Old-Growth Forest. Front. For. Glob. Change 2022, 5, 937404. [Google Scholar] [CrossRef]
- Unkule, M.; Piedallu, C.; Balandier, P.; Courbaud, B. Climate and Ungulate Browsing Impair Regeneration Dynamics in Spruce-Fir-Beech Forests in the French Alps. Ann. For. Sci. 2022, 79, 11. [Google Scholar] [CrossRef]
- Govedar, Z.; Krstić, M.; Keren, S.; Babić, V.; Zlokapa, B.; Kanjevac, B. Actual and Balanced Stand Structure: Examples from Beech-Fir-Spruce Old-Growth Forests in the Area of the Dinarides in Bosnia and Herzegovina. Sustainability 2018, 10, 540. [Google Scholar] [CrossRef]
- Petrovska, R.; Bugmann, H.; Hobi, M.L.; Brang, P. Replace Me If You Can: Abundance of Advance Regeneration under Canopy Trees in a Primeval Beech Forest. For. Ecol. Manag. 2023, 537, 120939. [Google Scholar] [CrossRef]
- Kanjevac, B.; Babić, V.; Stajić, S.; Martać, N.; Pavlović, B.; Furtula, D.; Čokeša, V. Key Drivers Affecting the Spatial Heterogeneity of the Regeneration Process in Old-Growth Beech Forests in Southeastern Europe. Front. For. Glob. Change 2023, 6, 1304037. [Google Scholar] [CrossRef]
- Braga, C.I.; Crisan, V.E.; Petritan, I.C.; Scarlatescu, V.; Vasile, D.; Lazar, G.; Petritan, A.M. Short-Term Effects of Anthropogenic Disturbances on Stand Structure, Soil Properties, and Vegetation Diversity in a Former Virgin Mixed Forest. Forests 2023, 14, 742. [Google Scholar] [CrossRef]
- Govedar, Z.; Prokhorova, N.; Babić, V.; Dukić, V.; Kanjevac, B.; Bilić, S. Natural Regeneration on Deadwood in the Primeval Forest Janj. Lesn. Zhurnal For. J. 2023, 5, 90–102. [Google Scholar] [CrossRef]
- Govedar, Z. Elementi Strukture i Prirodno Podmlađivanje Mješovite Sastojine Bukve, Jele i Smrče u Prašumi Janj. Šumarstvo 2002, 4, 9–24. [Google Scholar]
- Keren, S.; Diaci, J. Comparing the Quantity and Structure of Deadwood in Selection Managed and Old-Growth Forests in South-East Europe. Forests 2018, 9, 76. [Google Scholar] [CrossRef]
- UNESCO Ancient and Primeval Beech Forests of the Carpathians and Other Regions of Europe—UNESCO World Heritage Centre. Available online: https://whc.unesco.org/en/list/1133 (accessed on 21 March 2023).
- Maunaga, Z.; Burlica, Č.; Pavlović, B.; Bozalo, G.; Koprivica, M.; Lazarev, V.; Mataruga, M.; Repajić, Ž.; Govedar, Z.; Stanivuković, Z.; et al. Plan Gazdovanja Za Šume Sa Posebnom Namjenom u Strogim Rezervatima Prirode “Janj” i “Lom; Šumarski Fakultet Univerziteta u Banjoj Luci: Banja Luka, Bosnia and Herzegovina, 2005. [Google Scholar]
- Manuševa, L.; Fukarek, P.; Panov, A. Proučavanje Sastojina Prašumskog Tipa; Šumarski fakultet u Sarajevu: Sarajevo, Bosnia and Herzegovina, 1967. [Google Scholar]
- Govedar, Z. Načini Prirodnog Obnavljanja Mešovitih Šuma jele i smrče (Abieti—Piceetum Illyricum) na Području Zapadnog Dijela Republike Srpske. Ph.D. Thesis, University of Beograd, Beograd, Serbia, 2005. [Google Scholar]
- Keren, S. Complexity of Stand Structures in Two Mixed Mountain Old-Growth Forests and Adjacent Managed Forests in Bosnia and Herzegovina. Ph.D. Thesis, Biotechnical Faculty, University of Ljubljana, Ljubljana, Slovenia, 2015. [Google Scholar]
- Motta, R.; Berretti, R.; Castagneri, D.; Dukić, V.; Garbarino, M.; Govedar, Z.; Lingua, E.; Maunaga, Z.; Meloni, F. Toward a Definition of the Range of Variability of Central European Mixed Fagus–Abies–Picea Forests: The Nearly Steady-State Forest of Lom (Bosnia and Herzegovina). Can. J. For. Res. 2011, 41, 1871–1884. [Google Scholar] [CrossRef]
- Lojo, A.; Balić, B.; Mekić, F.; Beus, V.; Koprivica, M.; Treštić, T.; Musić, J.; Čabaravdić, A.; Hočevar, M. Državna Inventura Šuma—Bosna i Hercegovina—Faza 2: Manual—Uputstvo Za Snimanja Na Terenu. Rad. Šumar. Fak. Univ. U Sarajevu 2008, 20, 1–82. [Google Scholar] [CrossRef]
- Motta, R.; Berretti, R.; Lingua, E.; Piussi, P. Coarse Woody Debris, Forest Structure and Regeneration in the Valbona Forest Reserve, Paneveggio, Italian Alps. For. Ecol. Manag. 2006, 235, 155–163. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M. Gap Disturbance Regime in an Old-Growth Fagus–Abies Forest in the Dinaric Mountains, Bosnia-Herzegovina. Can. J. For. Res. 2008, 38, 2728–2737. [Google Scholar] [CrossRef]
- Le Roux, R.; Henrico, S.; Bezuidenhout, J.; Henrico, I. Inverse Distance Weighting as an Alternative Interpolation Method to Create Radiometric Maps of Natural Radionuclide Concentrations Using QGIS. Proc. Int. Cartogr. Assoc. 2023, 5, 1–7. [Google Scholar] [CrossRef]
- QGIS.org. QGIS 3.28. Geographic Information System. 2023. Available online: https://qgis.org/en/site/ (accessed on 31 March 2023).
- Drinić, P.; Matić, V.; Pavlič, J.; Prolić, N.; Stojanović, O.; Vukmirović, V.; Koprivica, M. Tablice Taksacionih Elemenata Visokih i Izdanačkih šuma u SR Bosni i Hercegovini; Šumarski Fakultet Univerziteta u Sarajevu: Sarajevo, Bosnia and Herzegovina, 1990; Volume Posebno Izdanje. [Google Scholar]
- Avery, T.E.; Burkhart, H.E. Forest Measurements; McGraw-Hill: New York, NY, USA, 2002; ISBN 978-0-07-113005-9. [Google Scholar]
- Motta, R.; Alberti, G.; Ascoli, D.; Berretti, R.; Bilić, S.; Bono, A.; Milić, Ć.; Vojislav, D.; Finsinger, W.; Garbarino, M.; et al. Old-Growth Forests in the Dinaric Alps of Bosnia-Herzegovina and Montenegro: A Continental Hot-Spot for Research and Biodiversity. Front. For. Glob. Change 2024, 7, 1371144. [Google Scholar] [CrossRef]
- Krause, G.H.M.; Arndt, U.; Brandt, C.J.; Bucher, J.; Kenk, G.; Matzner, E. Forest Decline in Europe; Development and Possible Causes. Water. Air. Soil Pollut. 1986, 31, 647–668. [Google Scholar] [CrossRef]
- Mina, M.; Bugmann, H.; Klopcic, M.; Cailleret, M. Accurate Modeling of Harvesting Is Key for Projecting Future Forest Dynamics: A Case Study in the Slovenian Mountains. Reg. Environ. Change 2017, 17, 49–64. [Google Scholar] [CrossRef]
- Piedallu, C.; Dallery, D.; Bresson, C.; Legay, M.; Gégout, J.-C.; Pierrat, R. Spatial Vulnerability Assessment of Silver Fir and Norway Spruce Dieback Driven by Climate Warming. Landsc. Ecol. 2023, 38, 341–361. [Google Scholar] [CrossRef]
- Drinić, P. Taksacioni Elementi Sastojina Jele, Smrče i Bukve Prašumskog Tipa u Bosni. Rad. Poljopr. Šumar. Fak. Univ. U Sarajev. 1956, 4, 107–160. [Google Scholar] [CrossRef]
- Anić, I.; Mikac, S. Struktura, tekstura i pomlađivanje dinarske bukovo-jelove prašume Čorkova uvala. Šumar. List 2008, 132, 505–515. [Google Scholar]
- Korpel, Š. Razvoj i Struktura Bukovo-Jelovih Prašuma i Njihova Primjena Kod Gospodarenja Prebornom Šumom. Šumar. List 1996, 120, 203–208. [Google Scholar]
- Pantić, D.; Medarević, M.; Banković, S.; Obradović, S.; Šjukić, B.; Pešić, B. Structural, Production and Dynamic Characteristics of the Strict Forest Reserve “Račanska Šljivovica” on Mt. Tara. Glas. Šumar. Fak. 2011, 103, 93–114. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Lexer, M.J. Unraveling the Drivers of Intensifying Forest Disturbance Regimes in Europe: Drivers of Forest Disturbance Intensification. Glob. Change Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Paillet, Y.; Parvainen, J.; Gosselin, M.; Gosselin, F.; Lier, M. Monitoring Forest Biodiversity in Europe: State of the Art, Challenges, and Opportunities. In Integrative Approaches as an Opportunity for the Conservation of Forest Biodiversity; European Forest Institute: Joensuu, Finland, 2013; p. 242. Available online: https://hal.inrae.fr/hal-02599083 (accessed on 9 June 2025).
- Solomon, L.W.; Arunrat, N.; Phutthai, T.; Wisawapipat, W.; Sereenonchai, S.; Hatano, R. Carbon Stock Estimation and Human Disturbance in Selected Urban Un-Conserved Forests in Entoto Mountain Forest, Addis Ababa, Ethiopia. Diversity 2025, 17, 225. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Year | Stem Density (ha−1 ± σ) | Mean Diameter (cm ± σ) | Basal Area (m2 ha−1 ± σ) | Growing Stock (m3 ha−1 ± σ) |
|---|---|---|---|---|---|
| Silver fir | 2011 | 119 ± 53.6 * | 60.1 ± 14.5 * | 34.6 ± 3.4 | 623.4 ± 389.6 |
| 2021 | 107 ± 48.5 | 62.2 ± 16.0 | 32.3 ± 3.1 | 579.1 ± 346.7 | |
| European beech | 2011 | 327 ± 163.4 | 58.6 ± 18.7 | 12.1 ± 1.0 | 191.5 ± 141.5 |
| 2021 | 333 ± 157.1 | 59.9 ± 20.1 | 12.1 ± 1.1 | 191.1 ± 150.1 | |
| Norway spruce | 2011 | 73 ± 44.3 * | 22.8 ± 9.0 | 21.1 ± 2.7 | 353.7 ± 282.9 |
| 2021 | 65 ± 38.4 | 22.2 ± 8.7 | 19.9 ± 2.9 | 331.2 ± 296.2 | |
| Total | 2011 | 520 ± 155.5 | 41.0 ± 7.1 | 67.8 ± 2.7 | 1168.7 ± 303.1 |
| 2021 | 504 ± 147.0 | 41.6 ± 6.9 | 64.3 ± 2.7 | 1101.4 ± 286.4 |
| Species | BA 2011 (m2 ha−1) | Mortality (m2 ha−1) | % | Recruitment | % | Transition | % | BA 2021 (m2 ha−1) | Difference (m2 ha−1) |
|---|---|---|---|---|---|---|---|---|---|
| Silver fir | 34.6 | −5.9 | −16.9 | 8.5 | 24.5 | −4.9 | −14.1 | 32.3 | −2.3 |
| European beech | 12.1 | −0.9 | −7.7 | 1.5 | 14.9 | −0.8 | −6.9 | 12.1 | 0.0 |
| Norway spruce | 21.1 | −3.8 | −17.8 | 5.9 | 28.1 | −3.4 | −16.2 | 19.9 | −1.3 |
| Total | 67.8 | −10.6 | −15.6 | 13.7 | 20.1 | −6.6 | −9.7 | 64.3 | −3.5 |
| Decay Class | % | Snags | Logs | Stumps | Total CWD |
|---|---|---|---|---|---|
| 1 | 7.9 | 29.2 | 7.4 | 0.0 | 36.7 |
| 2 | 25.1 | 71.4 | 44.2 | 0.2 | 115.9 |
| 3 | 17.9 | 36.4 | 45.5 | 1.0 | 82.8 |
| 4 | 22.7 | 17.3 | 82.7 | 5.1 | 105.1 |
| 5 | 26.4 | 1.5 | 117.4 | 3.3 | 122.2 |
| Total | 100.00 | 155.8 | 297.2 | 9.6 | 462.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilić, S.; Dukić, V.; Keren, S.; Ochał, W. Dynamics and Structural Changes in the Janj Mixed Old-Growth Mountain Forest: Continuing Decline of Conifers. Forests 2025, 16, 988. https://doi.org/10.3390/f16060988
Bilić S, Dukić V, Keren S, Ochał W. Dynamics and Structural Changes in the Janj Mixed Old-Growth Mountain Forest: Continuing Decline of Conifers. Forests. 2025; 16(6):988. https://doi.org/10.3390/f16060988
Chicago/Turabian StyleBilić, Srdjan, Vojislav Dukić, Srdjan Keren, and Wojciech Ochał. 2025. "Dynamics and Structural Changes in the Janj Mixed Old-Growth Mountain Forest: Continuing Decline of Conifers" Forests 16, no. 6: 988. https://doi.org/10.3390/f16060988
APA StyleBilić, S., Dukić, V., Keren, S., & Ochał, W. (2025). Dynamics and Structural Changes in the Janj Mixed Old-Growth Mountain Forest: Continuing Decline of Conifers. Forests, 16(6), 988. https://doi.org/10.3390/f16060988