
Vegetation Net Primary Productivity Dynamics over the Past Three Decades and Elevation–Climate Synergistic Driving Mechanism in Southwest China’s Mountains

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
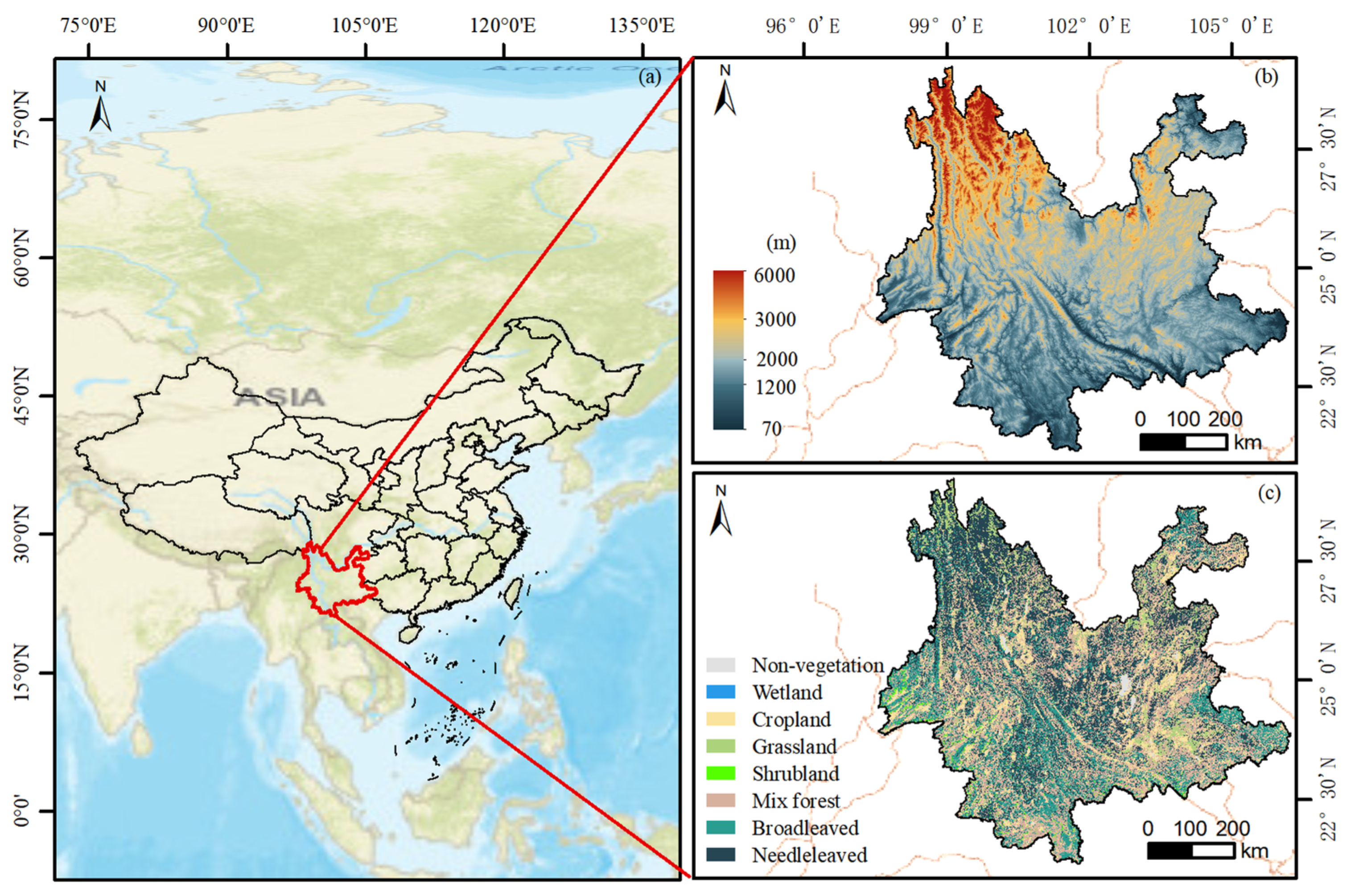
2.1. Study Area
2.2. NPP Modeling Methods
2.3. Driving Data
2.3.1. LAI Data
2.3.2. CI Data
2.3.3. LC and Meteorological Data
2.3.4. Soil Data
2.3.5. Topographic Data
2.3.6. Nighttime Light Data
2.4. Driving Factor Evaluation
2.4.1. Trend Analysis
2.4.2. Relative Importance Analysis
2.4.3. Geodetector Spatial Analysis
2.4.4. Elevation-Dependent Model
3. Results
3.1. NPP Changes in the Past Three Decades
3.2. NPP Changes in Different Vegetation Types
3.3. The Attribution of NPP Interannual Changes
3.4. The Attribution and Elevation Dependence of NPP Spatial Change
4. Discussion
4.1. Enhanced NPP Induced by CO2 Fertilization and Climate Warming
4.2. Vegetation-Specific NPP Enhancement and Biodiversity Synergy
4.3. Elevation-Dependence of NPP Spatial Patterns
4.4. Implications for Carbon-Sink Management and Climate Policy
5. Conclusions
- (1)
- NPP in the study area showed a significant increase, rising from 634 ± 325 to 748 ± 348 g C m−2 yr−1 (mean rate 4 g C m−2 yr−1), while the annual total NPP surged from 252 to 296 Tg C yr−1 (mean rate 2 Tg C yr−1). Spatially, the most rapid increases occurred in the eastern regions, contrasting with slower or negative trends in the northwestern plateau and peripheral western/southern border areas;
- (2)
- Rising CO2 (dominating 58% regions) and climate warming (dominating 17% regions) drove interannual NPP growth, with elevation thresholds dictating driver dominance. The CO2 governed low elevation, while temperature controlled higher elevation (>4800 m);
- (3)
- Our elevation-dependent model revealed a more complex, nonlinear relationship between NPP and elevation, refining traditional linear studies. Three distinct phases were identified by the nonlinear model: the saturation phase (<500 m) with stable NPP; the transition phase (500–3500 m) with linear decline (NPP loss of 29 g C m⁻2 yr⁻1 per 100 m); and the collapse phase (>3500 m) with continuously attenuated NPP losses reflecting high-elevation vegetation adaptation to extreme conditions;
- (4)
- The Geodetector analysis revealed that land cover dominated NPP spatial heterogeneity. Land cover synergistically amplified its influence through interactions with elevation and temperature, highlighting a vegetation–climate–topography coupling mechanism that critically shapes productivity patterns. Biodiversity-rich widespread mixed forests underpinned the region’s high productivity. Conservation programs should focus on protecting existing evergreen forests from logging and fragmentation, while forestation efforts should prioritize the establishment of biodiversity-rich mixed forests.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NPP | Net Primary Productivity |
| GPP | Gross Primary Productivity |
| BEPS | Boreal Ecosystem Productivity Simulator |
| LAI | Leaf area index |
| CI | Canopy clumping index |
| PAR | Photosynthetically active radiation |
| LC | Land cover |
| NTL | Nighttime light |
| DEM | Digital elevation models |
| LCCS | Land Cover Classification System |
| GRNNs | General Regression Neural Networks |
| ESA | European Space Agency |
References
- Pan, S.F.; Tian, H.Q.; Dangal, S.R.S.; Ouyang, Z.Y.; Tao, B.; Ren, W.; Lu, C.Q.; Running, S. Modeling and Monitoring Terrestrial Primary Production in a Changing Global Environment: Toward a Multiscale Synthesis of Observation and Simulation. Adv. Meteorol. 2014, 2014, 965936. [Google Scholar] [CrossRef]
- Sha, Z.Y.; Bai, Y.F.; Li, R.R.; Lan, H.; Zhang, X.L.; Li, J.; Liu, X.F.; Chang, S.J.; Xie, Y.C. The global carbon sink potential of terrestrial vegetation can be increased substantially by optimal land management. Commun. Earth Environ. 2022, 3, 8. [Google Scholar] [CrossRef]
- Huang, K.; Xia, J.Y.; Wang, Y.P.; Ahlström, A.; Chen, J.; Cook, R.B.; Cui, E.; Fang, Y.Y.; Fisher, J.B.; Huntzinger, D.N.; et al. Enhanced peak growth of global vegetation and its key mechanisms. Nat. Ecol. Evol. 2018, 2, 1897–1905. [Google Scholar] [CrossRef]
- Chen, J.M.; Ju, W.M.; Ciais, P.; Viovy, N.; Liu, R.G.; Liu, Y.; Lu, X.H. Vegetation structural change since 1981 significantly enhanced the terrestrial carbon sink. Nat. Commun. 2019, 10, 4259. [Google Scholar] [CrossRef]
- He, N.P.; Yan, P.; Liu, C.C.; Xu, L.; Li, M.X.; Van Meerbeek, K.; Zhou, G.; Zhou, G.; Liu, S.O.; Zhou, X.H.; et al. Predicting ecosystem productivity based on plant community traits. Trends Plant Sci. 2023, 28, 43–53. [Google Scholar] [CrossRef]
- Zhu, Z.C.; Piao, S.L.; Myneni, R.B.; Huang, M.T.; Zeng, Z.H.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Change 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Piao, S.L.; Liu, Q.; Chen, A.P.; Janssens, I.A.; Fu, Y.S.; Dai, J.H.; Liu, L.L.; Lian, X.; Shen, M.G.; Zhu, X.L. Plant phenology and global climate change: Current progresses and challenges. Glob. Change Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Zhu, P.F.; Liu, G.; He, J. Spatio-temporal variation and impacting factors of NPP from 2001 to 2020 in Sanjiangyuan region, China: A deep neural network-based quantitative estimation approach. Ecol. Inform. 2023, 78, 102345. [Google Scholar] [CrossRef]
- Perry, D.A.; Oetter, D.R. Thirty years of forest productivity in a mountainous landscape: The Yin and Yang of topography. Ecosphere 2024, 15, e4865. [Google Scholar] [CrossRef]
- Wen, A.B.; Tang, Q.; Ouyang, C.J.; Zhu, B.; Wang, Y.K.; Li, A.N.; Li, S.; Zhu, W.Z.; Liu, L.J. Mountain Protection and Mountain Development in China: Review and Prospect. Bull. Chin. Acad. Sci. 2023, 38, 376–384. [Google Scholar]
- Wei, D.; Tao, J.; Wang, Z.; Zhao, H.; Zhao, W.; Wang, X. Elevation-dependent pattern of net CO2 uptake across China. Nat. Commun. 2024, 15, 2489. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Chen, J.; Deng, T.; Sun, H. Plant diversity in Yunnan: Current status and future directions. Plant Divers. 2020, 42, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.L.; Feng, X.F.; Ge, Y.; Li, B.L. Topographical effects of climate data and their impacts on the estimation of net primary productivity in complex terrain: A case study in Wuling mountainous area, China. Ecol. Inform. 2015, 27, 44–54. [Google Scholar]
- Sun, T.; Tong, W.J.; Chang, N.J.; Deng, A.X.; Lin, Z.L.; Feng, X.B.; Li, J.Y.; Song, Z.W. Estimation of soil organic carbon stock and its controlling factors in cropland of Yunnan Province, China. J. Integr. Agric. 2022, 21, 1475–1487. [Google Scholar] [CrossRef]
- Bi, C.Y.; Yang, K.; Zhang, S.H.; Zeng, W.X.; Liu, J.; Rao, Y.; Ma, Y.; Yang, X.F. Simulation and analysis of afforestation potential areas under different development scenarios in Yunnan Province, China. Ecol. Indic. 2024, 167, 112695. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.P. Impacts of climate, phenology, elevation and their interactions on the net primary productivity of vegetation in Yunnan, China under global warming. Ecol. Indic. 2023, 154, 110533. [Google Scholar] [CrossRef]
- Fang, Y.; Wan, L.; Tong, J.; Li, G.J.; Pang, J.; Chang, E.; Chen, L.L.; Shi, Z.X. Hydrothermal conditions dominated sensitivity and lag effect of grassland productivity in Yunnan Province, China: Implications for climate change. Agric. Water Manag. 2025, 308, 109293. [Google Scholar] [CrossRef]
- Ye, H.; Bai, D.; Wang, J.L.; Tan, S.C.; Liu, S.Y.; Wu, X.P. Landscape ecological risk assessment study of the Yunnan section of the Tropic of Cancer. Ecol. Indic. 2024, 158, 111517. [Google Scholar]
- Xing, X.; Wu, M.; Zhu, H.; Duan, W.; Ju, W.; Wang, X.; Ran, Y.; Zhang, Y.; Jiang, F. Optimized Gross Primary Productivity Over the Croplands Within the BEPS Particle Filtering Data Assimilation System (BEPS_PF v1.0). J. Adv. Model. Earth Syst. 2025, 17, e2024MS004412. [Google Scholar]
- Xu, M.; Chen, J.M.; Liu, Y.; Wang, R.; Shang, R.; Leng, J.; Shu, L.; Liu, J.; Liu, R.; Liu, Y.; et al. Comparative assessment of leaf photosynthetic capacity datasets for estimating terrestrial gross primary productivity. Sci. Total Environ. 2024, 926, 171400. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef]
- Lin, W.Y.; Yuan, H.; Dong, W.Z.; Zhang, S.P.; Liu, S.F.; Wei, N.; Lu, X.J.; Wei, Z.W.; Hu, Y.; Dai, Y.J. Reprocessed MODIS Version 6.1 Leaf Area Index Dataset and Its Evaluation for Land Surface and Climate Modeling. Remote Sens. 2023, 15, 2072–4292. [Google Scholar] [CrossRef]
- Liang, S.L.; Cheng, J.; Jia, K.; Jiang, B.; Liu, Q.; Xiao, Z.Q.; Yao, Y.J.; Yuan, W.P.; Zhang, X.T.; Zhao, X.; et al. The Global Land Surface Satellite (GLASS) Product Suite. Bull. Amer. Meteorol. Soc. 2021, 102, E323–E337. [Google Scholar] [CrossRef]
- Baret, F.; Hagolle, O.; Geiger, B.; Bicheron, P.; Miras, B.; Huc, M.; Berthelot, B.; Niño, F.; Weiss, M.; Samain, O.; et al. LAI, fAPAR and fCover CYCLOPES global products derived from VEGETATION: Part 1: Principles of the algorithm. Remote Sens. Environ. 2007, 110, 275–286. [Google Scholar] [CrossRef]
- Jia, K.; Yang, L.Q.; Liang, S.; Xiao, Z.Q.; Zhao, X.; Yao, Y.; Zhang, X.; Jiang, B.; Liu, D. Long-Term Global Land Surface Satellite (GLASS) Fractional Vegetation Cover Product Derived from MODIS and AVHRR Data. IEEE J. Sel. Top. Appl. Earth Observ. Remote Sens. 2019, 12, 508–518. [Google Scholar] [CrossRef]
- Wei, S.S.; Fang, H.L. Estimation of canopy clumping index from MISR and MODIS sensors using the normalized difference hotspot and darkspot (NDHD) method: The influence of BRDF models and solar zenith angle. Remote Sens. Environ. 2016, 187, 476–491. [Google Scholar] [CrossRef]
- Wei, S.S.; Fang, H.L.; Schaaf, C.B.; He, L.M.; Chen, J.M. Global 500 m clumping index product derived from MODIS BRDF data (2001–2017). Remote Sens. Environ. 2019, 232, 111296. [Google Scholar] [CrossRef]
- ESA. Land Cover CCI Product User Guide Version 2. Tech. Rep. 2017. Available online: https://maps.elie.ucl.ac.be/CCI/viewer/download/ESACCI-LC-Ph2-PUGv2_2.0.pdf (accessed on 7 May 2025).
- He, J.; Yang, K.; Tang, W.J.; Lu, H.; Qin, J.; Chen, Y.Y.; Li, X. The first high-resolution meteorological forcing dataset for land process studies over China. Sci. Data 2020, 7, 25. [Google Scholar] [CrossRef]
- Tang, W.; Qin, J.; Yang, K.; Jiang, Y.; Pan, W. Mapping long-term and high-resolution global gridded photosynthetically active radiation using the ISCCP H-series cloud product and reanalysis data. Earth Syst. Sci. Data 2022, 14, 2007–2019. [Google Scholar] [CrossRef]
- Shangguan, W.; Dai, Y.J.; Liu, B.Y.; Ye, A.Z.; Yuan, H. A soil particle-size distribution dataset for regional land and climate modelling in China. Geoderma 2012, 171–172, 85–91. [Google Scholar] [CrossRef]
- Li, Q.; Shi, G.; Shangguan, W.; Nourani, V.; Li, J.; Li, L.; Huang, F.; Zhang, Y.; Wang, C.; Wang, D.; et al. A 1 km daily soil moisture dataset over China using in situ measurement and machine learning. Earth Syst. Sci. Data 2022, 14, 5267–5286. [Google Scholar] [CrossRef]
- Hu, Y.F.; Wang, G.J.; Wei, X.K.; Zhou, F.H.; Kattel, G.; Amankwah, S.O.Y.; Hagan, D.F.T.; Duan, Z. Reconstructing long-term global satellite-based soil moisture data using deep learning method. Front. Earth Sci. 2023, 11, 1130853. [Google Scholar] [CrossRef]
- ESA. Copernicus DEM-Global and European Digital Elevation Model. Eur. Space Agency 2019. [CrossRef]
- Chen, X.; Wang, Z.; Zhang, F.; Shen, G.; Chen, Q. A global annual simulated VIIRS nighttime light dataset from 1992 to 2023. Sci. Data 2024, 11, 1380. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Dong, Q.; Pan, Y.; Yang, F.; He, M.; Huang, X.; Xu, J. Spatiotemporal variations and driving forces of regional-scale NPP based on a multi-method integration: A case study in the Beibu Gulf Economic Zone. Ecol. Indic. 2025, 174, 113453. [Google Scholar] [CrossRef]
- Yang, S.; Yang, X.; Gao, X.; Zhang, J. Spatial and temporal distribution characteristics of carbon emissions and their drivers in shrinking cities in China: Empirical evidence based on the NPP/VIIRS nighttime lighting index. J. Environ. Manag. 2022, 322, 116082. [Google Scholar] [CrossRef]
- Groemping, U. Relative Importance for Linear Regression in R: The Package relaimpo. J. Stat. Softw. 2006, 17, 1–27. [Google Scholar]
- Ren, D.F.; Cao, A.H. Analysis of the heterogeneity of landscape risk evolution and driving factors based on a combined GeoDa and Geodetector model. Ecol. Indic. 2022, 144, 109568. [Google Scholar] [CrossRef]
- Forkel, M.; Carvalhais, N.; Rödenbeck, C.; Keeling, R.; Heimann, M.; Thonicke, K.; Zaehle, S.; Reichstein, M. Enhanced seasonal CO2 exchange caused by amplified plant productivity in northern ecosystems. Science 2016, 351, 696–699. [Google Scholar] [CrossRef]
- Wang, S.H.; Zhang, Y.G.; Ju, W.M.; Chen, J.M.; Ciais, P.; Cescatti, A.; Sardans, J.; Janssens, I.A.; Wu, M.S.; Berry, J.A.; et al. Recent global decline of CO2 fertilization effects on vegetation photosynthesis. Science 2020, 370, 1295–1300. [Google Scholar] [CrossRef]
- Piao, S.L.; Yin, G.D.; Tan, J.G.; Cheng, L.; Huang, M.T.; Li, Y.; Liu, R.G.; Mao, J.F.; Myneni, R.B.; Peng, S.S.; et al. Detection and attribution of vegetation greening trend in China over the last 30 years. Glob. Change Biol. 2015, 21, 1601–1609. [Google Scholar] [CrossRef]
- Wenzel, S.; Cox, P.M.; Eyring, V.; Friedlingstein, P. Projected land photosynthesis constrained by changes in the seasonal cycle of atmospheric CO2. Nature 2016, 538, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Haverd, V.; Smith, B.; Canadell, J.G.; Cuntz, M.; Mikaloff-Fletcher, S.; Farquhar, G.; Woodgate, W.; Briggs, P.R.; Trudinger, C.M. Higher than expected CO2 fertilization inferred from leaf to global observations. Glob. Change Biol. 2020, 26, 2390–2402. [Google Scholar] [CrossRef] [PubMed]
- He, Y.L.; Yan, W.B.; Cai, Y.; Deng, F.Y.; Qu, X.X.; Cui, X.L. How does the Net primary productivity respond to the extreme climate under elevation constraints in mountainous areas of Yunnan, China? Ecol. Indic. 2022, 138, 108817. [Google Scholar] [CrossRef]
- Xu, Y.; Lu, Y.G.; Zou, B.; Xu, M.; Feng, Y. Unraveling the enigma of NPP variation in Chinese vegetation ecosystems: The interplay of climate change and land use change. Sci. Total Environ. 2024, 912, 169023. [Google Scholar] [CrossRef]
- Reader, M.O.; Eppinga, M.B.; de Boer, H.J.; Damm, A.; Petchey, O.L.; Santos, M.J. Biodiversity mediates relationships between anthropogenic drivers and ecosystem services across global mountain, island and delta systems. Glob. Environ. Change-Hum. Policy Dimens. 2023, 78, 102612. [Google Scholar] [CrossRef]
- Xiao, D.O.; Deng, L.; Kim, D.; Huang, C.B.; Tian, K. Carbon budgets of wetland ecosystems in China. Glob. Change Biol. 2019, 25, 2061–2076. [Google Scholar] [CrossRef]
- Xu, H.Y.; Guli, J.; Yu, T.; Li, X.; Chen, B.J.; Abdalla, S. Analysis of spatio-temporal variation characteristics and influencing factors of net primary productivity in terrestrial ecosystems of China. Acta Ecol. Sin. 2023, 43, 1219–1233. [Google Scholar]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and Ecosystem Functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Feng, Y.H.; Schmid, B.; Loreau, M.; Forrester, D.I.; Fei, S.L.; Zhu, J.X.; Tang, Z.Y.; Zhu, J.L.; Hong, P.B.; Ji, C.J.; et al. Multispecies forest plantations outyield monocultures across a broad range of conditions. Science 2022, 376, 865–868. [Google Scholar] [CrossRef]
- Fei, S.L.; Jo, I.; Guo, Q.F.; Wardle, D.A.; Fang, J.Y.; Chen, A.P.; Oswalt, C.M.; Brockerhoff, E.G. Impacts of climate on the biodiversity-productivity relationship in natural forests. Nat. Commun. 2018, 9, 5436. [Google Scholar] [CrossRef]
- Shao, J.; Diao, Y.; Zhang, Q.; Wang, J. Optimizing conservation planning: An integrated approach to cost-effective biodiversity and carbon sequestration management in Yunnan Province, China. Glob. Ecol. Conserv. 2024, 56, e03336. [Google Scholar] [CrossRef]
- Li, W.; Wang, W.; Chen, J.; Zhang, Z. Assessing effects of the Returning Farmland to Forest Program on vegetation cover changes at multiple spatial scales: The case of northwest Yunnan, China. J. Environ. Manag. 2022, 304, 114303. [Google Scholar] [CrossRef]
- Liu, F.; Feng, C.; Zhou, Y.; Zhang, L.; Du, J.; Huang, W.; Luo, J.; Wang, W. Effectiveness of functional zones in National Nature Reserves for the protection of forest ecosystems in China. J. Environ. Manag. 2022, 308, 114593. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhou, Z.; Wu, X.; Wan, J.; Wang, J.; Zheng, J.; Liu, R.; Li, F. Prediction and spillover effects of forest expansion and management to increase carbon sinks in karst mountainous areas: A case study in Guizhou, China. Land Use Policy 2025, 151, 107491. [Google Scholar] [CrossRef]
- Adams, E.A.; Kuusaana, E.D.; Ahmed, A.; Campion, B.B. Land dispossessions and water appropriations: Political ecology of land and water grabs in Ghana. Land Use Policy 2019, 87, 104068. [Google Scholar] [CrossRef]
- Zhao, J.; Fu, X.; Sa, N.; Kou, X.; He, X.; Zheng, S.; Lu, Z.; Wu, G.; Sang, W. Forest eco-function restoration and its positive effects on biodiversity improvement in China’s ecological conservation programs. Ecol. Eng. 2025, 212, 107530. [Google Scholar] [CrossRef]
- Chen, C.; Xiao, W.Y.; Chen, H.Y.H. Meta-analysis reveals global variations in plant diversity effects on productivity. Nature 2025, 638, 435–440. [Google Scholar] [CrossRef]
- Xu, H.; Yue, C.; Zhang, Y.; Liu, D.; Piao, S.L. Forestation at the right time with the right species can generate persistent carbon benefits in China. Proc. Natl. Acad. Sci. USA 2023, 120, e2304988120. [Google Scholar] [CrossRef]
- Rodrigo, M.A. Wetland Restoration with Hydrophytes: A Review. Plants 2021, 10, 1035. [Google Scholar] [CrossRef]
- Tammeorg, O.; Chorus, I.; Spears, B.; Nõges, P.; Nürnberg, G.K.; Tammeorg, P.; Søndergaard, M.; Jeppesen, E.; Paerl, H.; Huser, B.; et al. Sustainable lake restoration: From challenges to solutions. WIREs Water 2024, 11, e1689. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhou, S.; Hou, Y.; Hu, Y.; Chen, C.; Liu, Y.; Yuan, L.; Cao, H.; Qian, B.; Liu, Y.; et al. Vegetation Net Primary Productivity Dynamics over the Past Three Decades and Elevation–Climate Synergistic Driving Mechanism in Southwest China’s Mountains. Forests 2025, 16, 919. https://doi.org/10.3390/f16060919
Li Y, Zhou S, Hou Y, Hu Y, Chen C, Liu Y, Yuan L, Cao H, Qian B, Liu Y, et al. Vegetation Net Primary Productivity Dynamics over the Past Three Decades and Elevation–Climate Synergistic Driving Mechanism in Southwest China’s Mountains. Forests. 2025; 16(6):919. https://doi.org/10.3390/f16060919
Chicago/Turabian StyleLi, Yang, Shaokun Zhou, Yongping Hou, Yuekai Hu, Chunpeng Chen, Yuanyuan Liu, Lin Yuan, Haobing Cao, Bintian Qian, Ying Liu, and et al. 2025. "Vegetation Net Primary Productivity Dynamics over the Past Three Decades and Elevation–Climate Synergistic Driving Mechanism in Southwest China’s Mountains" Forests 16, no. 6: 919. https://doi.org/10.3390/f16060919
APA StyleLi, Y., Zhou, S., Hou, Y., Hu, Y., Chen, C., Liu, Y., Yuan, L., Cao, H., Qian, B., Liu, Y., Yang, C., Wu, C., & Song, Y. (2025). Vegetation Net Primary Productivity Dynamics over the Past Three Decades and Elevation–Climate Synergistic Driving Mechanism in Southwest China’s Mountains. Forests, 16(6), 919. https://doi.org/10.3390/f16060919